
Abstract
Many of the Adélie penguin colonies used for long-term demographic studies are located near research stations, and there is a need to disentangle the effects of human activities and environmental variability on Adélie penguin population trends. This study used Geographic Information Systems and decision trees to examine whether potential changes in snow cover and/or proximity to human activities were able to explain the varying population trends of colonies at two breeding localities near Casey, East Antarctica. At the less visited site, Whitney Pt, wind exposure and snow accumulation correctly predicted the broad trends of colonies in most (84%) instances, whereas at Shirley I their predictive accuracy fell to 58%. At Shirley I, proximity to human activity correctly predicted the broad trends for 84% of colonies. While snow accumulation patterns are a primary driver of variation in population trends among colonies, the effect of snow accumulation is outweighed by proximity to human activities near Casey.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Adélie penguins (Pygoscelis adeliae) have been widely used as indicators of changes in Antarctic Ocean ecosystems (Micol and Jouventin 2001; Croxall et al. 2002; Kato et al. 2002, 2004). There are a number of reasons for this, including their perceived primary role in these ecosystems and the relative ease with which their populations can be monitored (Micol and Jouventin 2001; Kato et al. 2002). However, it has also been acknowledged that the use of birds as bioindicators of climate change may be problematic due to the complex and numerous interactions in Antarctic ecosystems (Croxall et al. 2002), and the potential confounding effects of human impacts at local scales. Adélie penguin colonies are known to be abandoned and recolonised as the climate changes (Ainley 2002; Emslie and Woehler 2005).
Many recent investigations of Adélie penguin population trends have focused on the effects of climate variability, mostly at meso- and macro-scales (Trivelpiece and Fraser 1996; Croxall et al. 2002; Kato et al. 2002, 2004; Forcada et al. 2006). Others have attempted to separate the effects of climate change and human activities (Fraser and Patterson 1997; Micol and Jouventin 2001, Patterson et al. 2003). Long-term studies of Adélie penguin populations have generally been conducted at sites that are easily accessed from research stations, where human activities are focused (e.g. Woehler et al. 1994; Fraser and Patterson 1997; Micol and Jouventin 2001; Woehler et al. 2001). Variability in the sizes of stations, the activities associated with them historically and currently, and their distances from penguin colonies makes it difficult to establish a broad understanding of the effects of human activities on Adélie penguins (Thomson 1977; Ainley et al. 1983; Woehler et al. 1994; Fraser and Patterson 1997; Micol and Jouventin 2001; Patterson et al. 2003). This makes it difficult to disentangle the relative effects of climatic variability and human activities on trends in breeding Adélie penguins (Fraser and Patterson 1997; Micol and Jouventin 2001; Patterson et al. 2003). As the number of people visiting Antarctica increases, so does concern about the potential impacts of human disturbance on Antarctic wildlife (Pfeiffer and Peter 2004). In 2005/06, more than 26,000 people visited Antarctica on tourism vessels (IAATO 2006). In addition, almost 4,000 people live in research stations located throughout Antarctica during the summer (COMNAP 2006).
Very few studies have attempted to investigate the spatial variability within and among Adélie penguin colonies within a breeding locality. Only Fraser and Patterson (1997) and Patterson et al. (2003) simulated snow accumulation patterns based on a digital elevation model (DEM) and hillshade analysis. They used human visitation records to investigate the relative effects of snow accumulation patterns and human impacts on breeding success of resident penguins.
This study aimed to determine the contribution of snow accumulation patterns, wind exposure and proximity to human activities to the observed long-term population trends of Adélie penguin colonies at two breeding localities in the Windmill Is, Antarctica. In addition, this study attempted to differentiate between environmental and human-induced effects on penguin population trends at a local scale. It improved on previous investigations of the effects of human activities and snow accumulation patterns on population trends in Adélie penguin colonies, by using a more accurate physically-based snow accumulation model and a high-resolution DEM to model finer scale patterns.
Methods
Study areas
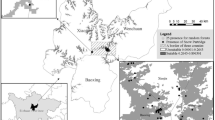
The Windmill Is are the islands and coastline covering an area of about 80 km2 around Casey (66° 17′ S, 110° 32′ E) in Wilkes Land, East Antarctica (Fig. 1). They comprise four large peninsulas and more than 30 islands (Murray and Luders 1990; Kirkup et al. 2002). During summer, the Windmill Is contain the only extensive areas of snow-free land in approximately 800 km of coast (Murray and Luders 1990; Kent et al. 1998). The Windmill Is contain extant Adélie penguin colonies on 14 islands and peninsulas, with a total population estimated at 93,000 ± 9,300 pairs in 1990 (Woehler et al. 1991). The Windmill Is weather is frigid-Antarctic (Melick et al. 1994). Weather observations between 1989 and 2004 showed that in the warmest month, January, mean daily temperatures ranged from −2.6° to 2.1°C. October was the coldest month of the breeding season, with a mean daily temperature range of −15.3° to −8.3°C.

Map of the Windmill Is, East Antarctica. Localities mentioned in the text are shown
Census data
A breeding colony is here defined as an area of contiguous nest territories sensu Woehler et al. (1991, 1994). A breeding locality is a geographical feature, either an island or a discrete area of mainland, on which breeding colonies are found. This contrasts with the use of the term “colony” to describe a breeding locality sensu Ainley (2002).
Whitney Pt (66° 15′ S, 110° 32′ E) is one of two mainland Adélie penguin breeding localities in the Windmill Is and is located on the Clark Peninsula, which was designated a site of special scientific interest (SSSI) in 1985. In 1996, the Clark Peninsula SSSI was redesignated Antarctic Specially Protected Area 136 (AAD 2006). Shirley I (66° 17′ S, 110° 29′ E) lies approximately 750 m west of Casey, across a 100 m-wide channel that is blocked with sea-ice for part of the year. Adélie penguin census data were available for the colonies at Whitney Pt for 22 breeding seasons between 1959/60 and 2005/06, and for the Shirley I colonies for 18 breeding seasons between 1968/69 and 2005/06 (Woehler et al. 1994; EJ Woehler unpub. data).
In February 2006, the Adélie penguin colonies at Shirley I and Whitney Pt were mapped with a differential GPS. The boundaries had a positional accuracy of approximately ±1 m (Bricher 2006). DEMs with a cell size of 2 m were derived from stereophotographs of the study sites (AADC 1994, 2001). These DEMs had a vertical accuracy of ±2 m (90% certainty) for snow-free areas. The extents of permanent snow and ice cover were digitised from an orthophoto of Shirley I derived from photographs taken in 2001, and from geo-referenced photographs of Whitney Pt taken in 1990. The boundary cells (2 m × 2 m) of current and relic Adélie penguin colonies were removed from the analyses because of the positional uncertainty of the colony boundaries. This increased the certainty that all cells described as having Adélie penguins present represented actual penguin colony habitat.
NCEP/NCAR weather reanalysis data
The NCEP/NCAR weather reanalysis data (Kistler et al. 2001) for the Casey region were extracted for three key periods in 1959/60, 1968/69 and 2005/06. The first two periods correspond to the summers in which the first Adélie penguin counts were conducted at Whitney Pt and Shirley I, respectively, and the 2005/06 data correspond with the most recent counts used in analyses for this study. The data were calculated for a point 63 km NNE of Casey (65.7125° S, 110.625° E). Australian Bureau of Meteorology data were available for the Windmill Is region from 1960 onwards, but it was considered that any changes in weather conditions were likely to have been obscured by two changes in the locations of weather observing equipment.
GIS processing methods
Wind exposure
The NCEP/NCAR data showed that almost all daily mean wind speeds greater than 10 ms−1 came from an ESE direction (100°–135°) (Fig. 2). Using ArcGIS 9.0 (ESRI 2005), we approximated the spatial distribution of wind exposure with a hillshade model, following the approach of Patterson et al. (2003). A hillshade model is a derivative of a DEM that simulates relative solar insolation for each grid cell based on its slope, aspect, and the position of the sun (as defined by an elevation and azimuth angle). Here, we used the hillshade model to simulate wind exposure by replacing the sun azimuth with the prevailing wind direction (122°) and by selecting a low elevation angle (5°). This approach generalized the physical characteristics of wind flow, but we hypothesize that the hillshade model provided a sufficiently accurate surrogate for wind exposure in the context of this study. For Shirley I, this layer was also used as a surrogate for exposure to potential airborne pollutants from Casey (Fig. 2).

Wind rose showing the direction of prevailing winds during the 2005/2006 breeding season, as modelled by the NCEP/NCAR weather reanalysis project. Data for the period 15 October 2005–31 January 2006 (3-h means) are shown. The dark bars indicate winds ≤10 m s−1, pale bars indicate winds 11–20 m s−1
Snow accumulation model
No long-term observations of snow cover were available for the Windmill Is. As snow cover is thought to be a major driving force for penguin colony locations, we chose to model the spatial patterns of snow accumulation, using a new GIS model to simulate the drifting of snow across Whitney Pt and Shirley I and the resulting short-term distribution of snow (Wallace 2005). Due to the physical complexity of the forces that drive snow transport, models of snow accumulation have generally involved a balance between accuracy, and the available input data and computational power (Liston and Sturm 1998). Therefore, factors which were considered less important for a particular application have typically been excluded in order to minimise computational expense (see Walter et al. 2004; Tappeiner et al. 2001).
The model used here required a DEM, an initial snow layer and daily mean temperature, air pressure, wind speed and direction data. The model accounted for wind deflection, wind speed, wind shear stresses, saltation, suspension and snow density (Wallace 2005). Saltation and suspension were treated as a single variable, after the approach taken by Kind (1981). The algorithms underlying the model are detailed in Bricher (2006). Commonly incorporated factors that have been excluded from this model comprise precipitation, sublimation, vegetation and snow melt, due to a lack of available data (Wallace 2005). Without direct measures of precipitation or initial snow depth, it was not possible to derive absolute snow-depths. Instead, the model produced a map of relative snow accumulation, following the approach of Purves et al. (1998), Ishikawa and Sawagaki (2001) and Orndorff and van Hoesen (2001).
The model was applied to a 5 m resolution DEM that covered the northern Windmill Is and the area immediately inland. This provided sufficiently large areas from which snow could blow into the study sites. Mid-November marks the peak laying period for Adélie penguins and is hence the time when snow cover was considered to be most crucial. We initialised the snow model with a uniform snow cover layer of 1 m thickness and used daily average temperature, wind speed, wind direction and air pressure data from the NCEP/NCAR weather reanalysis project (Kistler et al. 2001). To eliminate artefacts caused by the initial uniform snow cover, we ran the model for the 3-month period before November 15. This procedure was performed for the years of the first and most recent censuses at each site, i.e. 1959, 1968 and 2005. Once the snow accumulation data layers had been calculated, the layers for 1959 and 1968 (first counts for Whitney Pt and Shirley I, respectively) were subtracted from the 2005 layer. This produced layers showing the changes in short-term snow accumulation.
Human influences
Euclidean distance from Casey was calculated for both breeding localities. Shirley I is regularly visited by station personnel, so distance from the main island access point was also calculated. No equivalent variable could be applied to Whitney Pt, which is rarely visited because of its protected status as an ASPA. Likewise, at Shirley I, wind exposure was used as a surrogate for potential exposure to any airborne pollutants from Casey, as the island is located immediately downwind of the station. This is not the case for Whitney Pt, which is located 3 km upwind from Casey.
Population trends
Each colony was assigned to a population trend class. The census data for each colony was converted into percentages, with the initial census year assigned the value of 100%. The baseline was typically 1959/60 for Whitney Pt and 1968/69 for Shirley I. For colonies that were established after those years, the baseline year was the first year in which penguins were counted at that colony. The trend was based on the percentage difference between the year of first count and 2005. Population trends were then grouped into five classes: strongly decreasing (<50% of baseline population); moderately decreasing (50–80%); stable (80–120%); moderately increasing (120–150%) or strongly increasing (>150%) (Fig. 3).

Observed population trends of Adélie penguin colonies at (a) Whitney Pt and (b) Shirley I
Statistical analyses
To determine which combination of variables had the greatest impact on the population trends of Adélie penguins, the variables were examined individually and in combination to determine the relative effects of weather variables and proximity to human activities (Table 1). All analyses were conducted on the values of all cells within colonies and on the mean values for each colony. Analysing the individual cell values allowed for analysis of intra-colony variability, but left the analyses vulnerable to the effects of spatial autocorrelation. The results for the individual cell values are presented here, and for the colony means in Bricher (2006). However, both sets of analyses produce broadly similar results. None of the variables was normally distributed, so non-parametric tests were used. Wilcoxon two-group tests for difference were used to compare the distributions of values for each variable among population trend classes (Zar 1999).
A decision tree is a multivariate non-parametric classification technique capable of handling different data types (such as categorical and nonlinear data) and non-normal data (Witten and Frank 2005). Decision trees use Boolean logic to classify data into predetermined groups. This study used the J48 decision tree in Weka 3.4 (Witten and Frank 2005), which was derived from the C4.5 model developed by Quinlan (1993). Each of the study sites was examined individually. When classifying data into five groups, a purely random classifier could be expected to correctly predict the class with approximately 20% accuracy. Therefore, models were considered to have some explanatory power if they correctly classified significantly more than 20% of the test data points. The cells were randomly divided and 80% of grid cells were used to train the models and 20% used for validation. Decision trees were used to classify the population trends of grid cells using modelled snow cover for 15 November 2005, changes in the modelled snow cover between the first year of Adélie penguin count data (1959 for Whitney Pt and 1968 for Shirley I) and 2005, and exposure to prevailing winds as input variables. For Shirley I, the decision tree used distance from Casey, distance from the main island access point and wind exposure as input variables. The results of the analyses of wind exposure needed to be interpreted with caution, due to the potential for wind exposure to have dual impacts, such as potential air-borne pollution from Casey, in addition to its role in mediating snow cover. For Whitney Pt, Euclidean distance from Casey was the sole measure of proximity to human activity.
Results
Weather
The area had a modelled mean annual snowfall of 224.6 mm snow water equivalent (Bureau of Meteorology 2004). Between 1996 and 2006, monthly mean wind directions ranged between 92° and 186°, with the prevailing winds coming from ESE (100°–135º). During the Adélie penguin breeding seasons, the mean wind speed was 12.6 knots, with a mean monthly maximum wind gust of 65.1 knots. On average winds exceeded gale force (37 kts) on 32.5 days a year (AADC 2006).
Wind exposure and snow accumulation model
There were very weak negative correlations between snow accumulation and wind exposure (Whitney Pt R 2 = 0.006; Shirley I R 2 = 0.043). Areas of snow ablation and accumulation generally agreed with snow accumulation patterns observed in aerial photographs (AADC 1990, 2001). The model did not predict the areas of permanent snow and ice that occur along the eastern and southern coasts of the study sites, but it did predict the snow that accumulates in the valley to the south of Whitney Pt (Fig. 4). This suggests that the model could not account for the local effects of sea-ice that builds up around the Antarctic coast in the study area. No area of either study site showed broad patterns of increases or decreases in snow accumulation between the two years for which snow cover was modelled (1959–2005 for Whitney Pt, 1968–2005 for Shirley I).

a Modelled snow accumulation at Whitney Pt, November 2005; b modelled difference in snow accumulation Whitney Pt, November 1959–November 2005; c modelled snow accumulation at Shirley I, November 2005; d Modelled difference in snow accumulation Shirley I, November 1968–November 2005
Adélie penguin colony population trends
Snow accumulation
At Whitney Pt, the colonies with strong population increases (>150% of the baseline population) were associated with the thinnest snow cover, while stable colonies were found in areas with thicker snow cover. Wilcoxon tests found significant differences among trend classes for each of the three snow accumulation variables (Table 2). Colonies in all population trend classes were associated with areas where there were minimal changes in the modelled snow cover between 1959 and 2005.
The difference in snow cover between 1959 and 2005 did not improve the predictive power of the decision tree. Validation showed that the tree based on the 2005 snow accumulation and wind exposure had 83.7% accuracy. The model was most accurate in predicting strongly increasing colonies (class 5: 98.8%, Table 3). In contrast, the model failed to predict accurately any of the decreasing colonies (classes 1 and 2) at Whitney Pt. However, these are small groups. The predictive map showed that the model predicted strong population increases for almost all cells in all colonies (Fig. 5a). The major exception was a small group of colonies at the western end of Whitney Pt, which is located at the base of a short snow cliff, where the model predicted a mixture of stable and strongly increasing trends. Overall, the map showed strong predictions except where the DEM failed to show a small topographic feature.

Predicted population trends based on snow variables at (a) Whitney Pt and (b) Shirley I
At Shirley I, the Wilcoxon test showed significant differences among the five population trend classes for all the variables associated with snow accumulation patterns (Table 4). A decision tree was constructed using the November 2005 snow cover, wind exposure and the change in snow cover between 1959 and 2005 as inputs. Validation showed that it had an overall accuracy of 57.8%. The model most accurately predicted strongly increasing colonies (class 5: 73%: Table 5). The resulting map (Fig. 5b) predicted that the colonies at the western end would have stable or increasing populations, while those at the eastern were generally predicted to be decreasing. The model correctly predicted the trend class for some cells in seven out of 21 strongly decreasing colonies, and entirely failed to predict accurately a cluster of strongly decreasing colonies in the central north of the island. For most of the island, errors were within one class of the observed trend, but in the central north, the strongly decreasing colonies were predicted as either stable or strongly increasing. At Shirley Island, a decision tree based on snow cover variables had limited power to predict Adélie penguin colony population trends.
Proximity to human activities
At Whitney Pt, no clear relationship could be found between the colonies’ population trend and their distance from Casey, despite the Wilcoxon test showing a significant difference among the classes for the distance from Casey (Table 6). Strongly increasing colonies occurred at all distances from Casey, while stable (class 3) and moderately decreasing (class 2) colonies were clustered closest to Casey. Strongly decreasing (class 1) and moderately increasing (class 4) colonies were found near the middle of the study site (Fig. 3a).
A decision tree was constructed based on distance from Casey. Validation showed that it correctly predicted penguin colony population trends for 90.9% of test cells at Whitney Pt. The predictive map (Fig. 6a) shows three distinct bands of cells in class 5 (strong increase), interspersed by a band of cells with stable penguin populations and a band of cells with moderately increasing penguin colony populations (class 4). Both the colonies closest to Casey and farthest away were classified as class 5. The model most accurately predicted increasing colonies (class 4: 100% and class 5: 94.1%). It failed to predict accurately the trends of any decreasing colonies (Table 7) although these classes contained low numbers of colonies. This model more accurately predicted the performance of the stable colonies at the western end of the point than the model based on snow variables. It also correctly predicted the trends of two moderately increasing colonies at the western end of Whitney Pt, which the other model failed to.

Predicted population trends based on human proximity variables at (a) Whitney Pt and (b) Shirley I
At Shirley I, there were significant differences among the five population trend classes for all the parameters associated with proximity to human activity (Table 8). All population trend classes were clustered around the median values for wind exposure. Almost all the strongly increasing colonies were found farthest from Casey and the island access point. Most of the stable colonies occurred at medium to long distances from Casey and the island access point. Moderately decreasing colonies were clustered very close to Casey, and close to the access point. Strongly decreasing colonies were bimodal, occurring close to Casey and the access point, and also at moderately long distances from both (Fig. 3b).
A decision tree based on the variables associated with human proximity predicted Shirley I population trends with 83.8% accuracy. The distance from the sea-ice crossing point and from Casey had the most predictive power. The model most accurately predicted trends for increasing colonies (class 4: 87.7% and class 5: 86.5%; Table 9). The model predicted (Fig. 6b) that all increasing colonies occur at the greatest distance from Casey, with stable and decreasing colonies occurring closer to the station. The predictive map closely matched the map of observed population trends. The biggest exception was five very small colonies near the middle of the island, which were predicted to be decreasing strongly, but were observed to be decreasing moderately.
Discussion
The DEMs that were created for this study are the most detailed elevation data produced to date for Shirley I and Whitney Pt. However, it is likely that improved DEMs, as a result of better aerial photography and more accurate ground control points, would increase the reliability of the results reported here as wind exposure and snow accumulation models rely heavily on fine scale terrain information. Improved positional accuracy would also reduce the need to remove cells at the edges of colonies from the analysis. This would in turn mean that very small colonies would not be excluded from the analyses. This is potentially significant because smaller colonies have higher perimeter to area ratios, and it is possible that the landscape properties of these small colonies differ from those of larger colonies. It is also possible that the effects of environmental or human-related stressors are more severe for these small colonies, as has been shown in other parts of Antarctica (e.g. Giese 1996; Patterson et al. 2003). Since the smallest colonies on Shirley I are concentrated at the eastern end of the island, where the majority of strongly decreasing colonies are located, this is a subject that needs further investigation.
The models presented here are potentially affected by spatial autocorrelation. However, the analyses were repeated on mean values for each colony, hence removing intra-colony spatial autocorrelation, and produced similar results (see Bricher 2006). In future work, we are planning to include spatial autocorrelation in our models.
The NCEP/NCAR weather reanalysis data were produced for a grid point located 63 km to sea NNE of Casey. Thus, the effects of local topographic features in mediating the weather conditions at the study sites could not be examined in this study. There is also a need for further validation of the snow accumulation model, though it showed strong visual agreement with the available aerial photography.
The effects of snow accumulation patterns, wind exposure and proximity to human activities on Adélie penguin colony population trends.
On the Antarctic Peninsula, snow accumulation patterns have been found to be important predictors of Adélie penguin colony population trends, and of colony distributions (Fraser and Patterson 1997; Patterson et al. 2003). This study’s results supported those findings. At Whitney Pt, the modelled snow accumulation and wind exposure layers explained much of the variability in colony population trends. These layers had less predictive power on Shirley I, where proximity to human activities explained a significant proportion of the variability on colony population trends.
Patterson et al. (2003) conducted their study in an area where increasing mean temperatures led to increased snowfall. In the Windmill Is, little attention has been paid to local effects of climate change, and the modelled change in snow cover in this study suggested that there was little change in snow accumulation patterns between the beginning and end of the study period (1959–2005). The results suggest that while a broader-scale process is driving the overall increase in Adélie penguin numbers for the Whitney Pt colonies, differences in snow accumulation patterns mediate that increase in individual colonies.
Distance from Casey had a significant effect on the population trends of colonies at Whitney Pt. The model predicted trends with 91% accuracy. The fact that the colonies both closest to and farthest from Casey had strong population increases suggests that some other environmental factor is driving the variability in population trends. At Shirley I, proximity to human activities had greater predictive power for colony population trends than at Whitney Pt. Significant differences were present among population trend classes for distance from Casey and from the sea-ice crossing point used by visitors to access the island. While these results suggest that proximity to human activities was a significant driver of Adélie penguin colony population trends, further investigations are required to separate the potential effects of station-related activities, such as noise and particulate emission, and the effects of visits to the colonies by station personnel, given that Shirley I is downwind of Casey and the two distance measures covaried.
Previous studies of the effects of human activities on Adélie penguin population trends have produced site-specific results. The results of the present study at least partially support the proposition of Woehler et al. (1994) that visits by station personnel appeared to be causing decreases in populations among some Adélie penguin colonies on Shirley I. At Cape Bird on Ross I, Adélie penguin colonies close to a small research station underwent significant population decreases at a time when the overall penguin population was increasing (Young 1990). Human activities there included helicopter operations, whereas helicopter flights are routed away from penguin colonies around Casey. Similarly, Giese (1996) reported significantly lower breeding success in Adélie penguin colonies that had been subjected to daily recreational visitors or to regular scientific nest-checks. She concluded that the frequency of disturbance influenced the magnitude of the decrease in breeding success. Giese’s study was conducted in a breeding locality that had been little disturbed by previous human activities. In contrast, Patterson et al. (2003) investigated colonies on Torgersen I (64° 46′ S, 64° 5′ W) near Palmer on the Antarctic Peninsula that had been regularly visited by tourists and researchers for many years. They found that tourism had no detectable effect on Adélie penguin colony population size or breeding success, but noted that landscape-scale processes were operating. Similarly, Fraser and Patterson (1997) found no correlation between Adélie penguin population trends and human-use histories of breeding localities in the vicinity of Palmer.
Thomson (1977) found that after the Adélie penguin population at Cape Royds had decreased markedly, the colony began to recover after the introduction of regulations preventing helicopter over-flights and people from walking through the colonies. These restrictions were similar to those in operation at Shirley I. Ainley et al. (1983) found that human activities were likely to be responsible for reductions in the number of chicks in colonies near a research hut at Cape Crozier.
A few studies of the effects of proximity and/or exposure to human activities on the breeding success and population trends of Adélie penguins have also examined the effects of habitat modification arising from infrastructure construction (Wilson et al. 1990; Micol and Jouventin 2001). From these studies, it appears that the effects of proximity and/or exposure to human activities are determined by a combination of the types of activities involved and the history of interactions between humans and Adélie penguins at a given site.
The results of this study suggest that proximity to human activities plays a highly significant role in mediating the observed long-term increases in Adélie penguin numbers in the Windmill Is. Of the colonies for which long-term census data are available, the majority of colonies with decreasing population trends are located at the eastern end of Shirley I, which is the closest breeding locality to Casey (Woehler et al. 1991).
References
Ainley DG (2002) The Adélie penguin: bellwether of climate change. Columbia University Press, New York
Ainley DG, LeResche RE, Sladen WJL (1983) Breeding Biology of the Adélie Penguin. University of California Press, Berkeley
Australian Antarctic Data Centre (1994) Aerial photographs of Whitney Point, Film ANT 1007, Run 19, Australian Antarctic Division (Commonwealth of Australia), http://www.aad.gov.au
Australian Antarctic Data Centre (2001) Aerial photographs of Shirley Island, Film ANTC 1099, Run, 1–3, Australian Antarctic Division (Commonwealth of Australia), http://www.aad.gov.au
Australian Antarctic Data Centre (2006) Automatic weather station data from Casey, http://aadc-db.aad.gov.au, last updated 25 May 2006. Accessed 28 August 2006
Australian Antarctic Division (2006) Clark Peninsula ASPA No 136, http://www.aad.gov.au, last updated 1 September 2004. Accessed 7 September 2006
Bricher PK (2006) Spatial ecology of Adélie penguin breeding colonies: The effects of landscape, environmental variability and human activities. M Env Mgt thesis, University of Tasmania, Hobart
Bureau of Meteorology (2004) Climate averages for Australian sites, http://www.bom.gov.au, last updated 16 August 2004. Accessed 5 September 2006
COMNAP (2006) Antarctic facilities in operation. http://www.comnap.aq/operations/facilities/, Council of Managers of National Antarctic Programs. Accessed 28 October 2006
Croxall JP, Trathan PN, Murphy EJ (2002) Environmental change and Antarctic Seabird Populations. Science 297:1510–1514
Emslie SD, Woehler EJ (2005) A 9000-year record of Adélie penguin occupation and diet in the Windmill Islands, East Antarctica. Antarct Sci 17(1):57–66
Forcada J, Trathan PN, Reid K, Murphy EJ, Croxall JP (2006) Contrasting population changes in sympatric penguin species in association with climate warming. Glob Change Biol 12:411–423
Fraser WR, Patterson DL (1997) Human disturbance and long-term changes in Adélie penguin populations: a natural experiment at Palmer Station, Antarctic Peninsula. In: Battaglia B, Valencia J, Walton D (eds) Antarctic communities: species, structure and survival. Cambridge University Press, Cambridge, pp 445–452
Giese M (1996) Effects of human activity on Adélie penguin Pygoscelis adeliae breeding success. Biol Conserv 75:157–164
International Association of Antarctic Tourism Operators (IAATO) (2006) http://www.iaato.org. Accessed 28 August 2006
Ishikawa M, Sawagaki T (2001) GIS-simulation of the spatial distribution of snow cover and observed ground temperatures in the Daisetsu Mountains, Japan. Nor Geogr Tidsskr 55:212–218
Kato A, Ropert-Coudert Y, Naito Y (2002) Changes in Adélie penguin breeding populations in Lützow-Holm Bay, Antarctica, in relation to sea-ice conditions. Polar Biol 25:934–938
Kato A, Watanabe K, Naito Y (2004) Population changes of Adélie and Emperor penguins along the Prince Olav Coast and on the Riiser-Larsen Peninsula. Polar Biosci 17:117–122
Kent S, Seddon J, Robertson G, Wienecke B (1998) Diet of Adélie Penguins Pygoscelis adeliae at Shirley Island, East Antarctica, January 1992. Mar Ornithol 26:7–10
Kind RJ (1981) Snow drifting. In: Gray DM, Male DH (eds) Handbook of snow. Pergamon, Tarrytown, pp 338–359
Kirkup H, Melles M, Gore D (2002) Late Quaternary environment of southern Windmill Islands, East Antarctica. Antarct Sci 14:385–394
Kistler R, Kalnay E, Collins W, Saha S, White G, Woollen J, Chelliah M, Ebisuzaki W, Kanamitsu M, Kousky V, van den Dool H, Jenne R, Fiorino M (2001) The NCEP/NCAR 50-year reanalysis. Bull Am Meteorol Soc 82(2):247–267
Liston GE, Sturm M (1998) A snow-transport model for complex terrain. J Glaciol 44(148):498–516
Melick DR, Hovenden MJ, Seppelt RD (1994) Phytogeography of bryophyte and lichen vegetation in the Windmill Islands, Wilkes Land, Continental Antarctica. Plant Ecol 111(1):71–87
Micol T, Jouventin P (2001) Long-term population trends in seven Antarctic seabirds at Pointe Géologie (Terre Adélie): human impact compared with environmental change. Polar Biol 24:175–185
Murray MD, Luders DJ (1990) Faunistic studies at the Windmill Islands, Wilkes Land, East Antarctica, 1959–80. ANARE Res Notes 73
Orndorff RL, Van Hoesen JG (2001) Using GIS to predict the spatial distribution of perennial snow under modern and late Pleistocene conditions in the Snake Range, Nevada. 69th Annual Meeting of the Western Snow Conference, Las Vegas, Nevada, 2001, pp 119–122
Patterson DL, Easter-Pilcher AL, Fraser WR (2003) The effects of human activity and environmental variability on long-term changes in Adélie penguin populations at Palmer Station, Antarctica. In: Huiskes AHL, Gieskes WWC, Rozema J, Schorno RML, van der Vies SM, Wolf WJ (eds) Antarctic Biology in a Global Context. Backhuys Publishers, Leiden, pp 301–307
Pfeiffer S, Peter H-U (2004) Ecological studies toward the management of an Antarctic tourist landing site (Penguin Island, South Shetland Islands). Polar Rec 40(215):345–353
Purves RS, Barton JS, Mackaness WA, Sugden DE (1998) The development of a rule-based spatial model of wind transport and deposition of snow. Ann Glaciol 26:197–202
Quinlan JR (1993) C4.5: programs for machine learning. Morgan Kaufmann Publishers Inc, San Francisco, CA
Tappeiner U, Tappeiner G, Aschenwald J, Tasser E, Ostendorf B (2001) GIS-based modelling of spatial pattern of snow cover duration in an alpine area. Ecol Model 138:265–275
Thomson RB (1977) Effects of human disturbance on an Adélie penguin rookery and measures of control. In: Llano GA (ed) Adaptations Within Antarctic Ecosystems: Proceedings of the Third SCAR Symposium on Antarctic Biology. Gulf Publishing Company, Houston, Texas
Trivelpiece, WZ, Fraser, WR (1996) The breeding biology and distribution of Adélie penguins: adaptations to environmental variability. Foundations for Ecological Research West of the Antarctic Peninsula, Antarct Res Ser 70. Am Geophys Union, Washington DC, pp 273–285
Wallace L (2005) GIS based snow accumulation model for the Windmill Islands. Geomatics Studio Project, University of Tasmania, Hobart
Walter MT, McCool DK, King LG, Molnau M, Campbell GS (2004) Simple snowdrift model for distributed hydrological modeling. J Hydrol Eng July/August 2004:280–287
Wilson K-J, Taylor RH, Barton KJ (1990) The impact of man on Adélie penguins at Cape Hallett, Antarctica. In: Kerry KR, Hempel G (eds) Antarctic ecosystems. Ecological change and conservation. Springer, Berlin, pp 183–190
Witten IH, Frank E (2005) Data mining: practical machine learning tools and techniques, 2nd edn. Morgan Kaufmann, San Francisco
Woehler EJ, Slip DJ, Robertson LM, Fullagar PJ, Burton HR (1991) The distribution, abundance and status of Adélie penguins Pygoscelis adeliae at the Windmill Islands, Wilkes Land, Antarctica. Mar Ornithol 19(1):1–18
Woehler EJ, Penney RL, Creet SM, Burton HR (1994) Impacts of human visitors on breeding success and long-term population trends in Adélie penguins at Casey, Antarctica. Polar Biol 14:269–274
Woehler EJ, Cooper J, Croxall JP, Fraser WR, Kooyman GL, Miller GD, Nel DC, Patterson DL, Peter H-U, Ribic CA, Salwicka K, Trivelpiece WZ, Weimerskirch, H (2001) A statistical assessment of the status and trends of Antarctic and Subantarctic seabirds. SCAR Cambridge
Young EC (1990) Long-term stability and human impact in Antarctic skuas and Adélie penguins. In: Kerry KR, Hempel G (eds) Antarctic ecosystems. ecological change and conservation. Springer, Berlin, pp 231–236
Zar JH (1999) Biostatistical analysis, 4th edn. Prentice-Hall, Englewood Cliffs
Acknowledgments
The DEM was created by Robert Anders and the snow accumulation model by Luke Wallace at the University of Tasmania. We thank Ben Raymond and Neil Adams for their advice, and Casey personnel for their assistance in counting penguins. We also thank Rob Crawford and David Ainley for their kind comments and helpful suggestions on an earlier draft.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Bricher, P.K., Lucieer, A. & Woehler, E.J. Population trends of Adélie penguin (Pygoscelis adeliae) breeding colonies: a spatial analysis of the effects of snow accumulation and human activities. Polar Biol 31, 1397–1407 (2008). https://doi.org/10.1007/s00300-008-0479-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00300-008-0479-z