
Abstract
The Ross Sea, a large, high-latitude (72–78°S) embayment of the Antarctic continental shelf, averages 500 m deep, with troughs to 1,200 m and the shelf break at 700 m. It is covered by pack ice for 9 months of the year. The fish fauna of about 80 species includes primarily 4 families and 53 species of the endemic perciform suborder Notothenioidei. This review focuses on the diet and role in the food web of notothenioids and top-level bird and mammal predators, and also includes new information on the diets of artedidraconids and bathydraconids. Although principally a benthic group, notothenioids have diversified to form an adaptive radiation that includes pelagic and semipelagic species. In the southern Ross Sea, notothenioids dominate the fish fauna at levels of abundance and biomass >90% and are, therefore, inordinately important in the food web. Antarctic krill (Euphausia superba) and mesopelagic fishes are virtually absent from the shelf waters of the Ross Sea. Of the four notothenioid families, nototheniids show the most ecological and dietary diversification, with pelagic, cryopelagic, epibenthic and benthic species. Neutrally buoyant Pleuragramma antarcticum constitutes >90% of both the abundance and biomass of the midwater fish fauna. Most benthic nototheniids are opportunistic and feed on seasonally or locally abundant zooplanktonic prey. Artedidraconids are benthic sit-and-wait predators. Larger bathydraconids are benthic predators on fish while smaller species feed mainly on benthic crustaceans. Channichthyids are less dependent on the bottom for food than other notothenioids. Some species combine benthic and pelagic life styles; others are predominantly pelagic and all consume euphausiids and/or fish. South polar skuas, Antarctic petrels, Adélie and emperor penguins, Weddell seals and minke and killer whales are the higher vertebrate components of the food web, and all prey on notothenioids to some extent. Based on the frequency of occurrence of prey items in the stomachs of fish, bird and mammal predators, P. antarcticum and ice krill E. crystallorophias are the key species in the food web of the Ross Sea. P. antarcticum is a component of the diet of at least 11 species of nototheniid, bathydraconid and channichthyid fish and, at frequencies of occurrence from 71 to 100%, is especially important for Dissostichus mawsoni, Gvozdarus svetovidovi and some channichthyids. At least 16 species of notothenioids serve as prey for bird and mammal predators, but P. antarcticum is the most important and is a major component of the diet of south polar skua, Adélie and emperor penguins and Weddell seals, at frequencies of occurrence from 26 to 100%. E. crystallorophias is consumed by some nototheniid and channichthyid fish and can be of importance in the diet of emperor and Adélie penguins, although in the latter case, this is dependent on location and time of year.
Unlike the linear phytoplankton→E. superba→consumers of the E. superba food chain hypothesized for much of the Southern Ocean, the food web of the Ross Sea shelf is non-linear, with complex prey-predator interactions. Notothenioid fish play a key role: as predators, they occupy most of the trophic niches available in the ecosystem, relying on benthic, zooplanktonic and nektonic organisms; as prey, they are important food resources for each other and for most top predators living and foraging on the shelf. They also constitute the major link between lower (invertebrates) and higher (birds and mammals) levels of the food web. This is especially true for P. antarcticum. Along with E. crystallorophias, its ecological role in the Ross Sea is equivalent to that of myctophids and E. superba elsewhere in the Southern Ocean.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The Southern Ocean consists of the southern parts of the Atlantic, Indian and Pacific oceans (Lutjeharms 1990). The northern boundary of the Southern Ocean is the Antarctic Polar Front, located at about 50°S in the Atlantic and Indian sectors and 60°S in the Pacific sector (Hureau 1994; Lutjeharms 1990). The Antarctic Polar Front is characterized by a sudden drop of surface temperature of 3–4°C, as well as by changes in other oceanographic parameters like salinity. At the front, northward-flowing Antarctic Surface Water sinks beneath warmer Subantarctic Surface Water, generating a transition zone between surface water masses of different temperatures (Eastman 1993). From an oceanographic perspective, the surface circulation around Antarctica consists of two main currents. Close to the Antarctic Continent, easterly winds generate a counterclockwise water circulation, i.e. the Antarctic Coastal Current or East Wind Drift. North of 60°S, westerly winds produce a clockwise and northward water flow, known as Antarctic Circumpolar Current or West Wind Drift, representing the major current in the Southern Ocean. In large embayments of the Antarctic Continent, particularly the Weddell and Ross Seas, the Antarctic Coastal Current forms clockwise gyres, which probably concentrate nutrients like silicate, phosphate and nitrate (Lutjeharms 1990). Unlike other areas of Antarctica, the continental shelves of the Weddell and Ross Seas are wide and 500–600 m deep, with inner shelf depressions reaching depths of over 1,200 m (Anderson 1999). Furthermore, much of the Antarctic coastline is covered by ice shelves, which are exceptionally large in the Weddell and Ross Seas. During much of the year, the adjacent shelf waters are covered by pack ice.
Based on the distribution of the pelagic and demersal fishes, three main ichthyofaunal subregions are recognized in the Southern Ocean: from north to south, the Ice-free Zone, the Seasonal Pack-ice Zone and the High-Antarctic Zone (Kock 1992).
The Ice-free Zone, where ice cover is absent all year, lies between the Antarctic Polar Front and the northern limit of the pack ice in winter/spring. It is deep sea, almost exclusively inhabited by mesopelagic and bathypelagic fish, and most of these are cosmopolitan rather than members of the indigenous Antarctic perciform suborder Notothenioidei.
The Seasonal Pack-ice Zone extends between the limit of pack ice in winter/spring and in autumn/summer, including the islands north of the maximum limit of the pack ice. The pelagic fauna is represented by mesopelagic species and early life-stages of notothenioids. The benthic fish fauna is dominated by nototheniids of the genera Notothenia and Lepidonotothen, as well as by harpagiferids, bathydraconids and channichthyids.
Finally, the High-Antarctic Zone is that region immediately adjacent to the continent which is covered by ice for most of the year. The fish fauna is similar to that of the Seasonal Pack-ice Zone, at least for the pelagic fauna. However, the benthic fauna is greatly restricted in diversity of higher taxa, and characterized by nototheniids of the genus Trematomus and a high proportion of other notothenioids, such as artedidraconids, bathydraconids and channichthyids. In the Antarctic, fish species diversity, especially that of notothenioids, generally increases with latitude, although biomass decreases farther south. For example, fish biomass is 10- to 20-fold higher in the Seasonal Pack-ice Zone near Elephant Island than in the High-Antarctic southern Weddell Sea, but indices of species diversity and evenness are lower at Elephant Island than at comparable depths on the High-Antarctic shelf (Tiedtke and Kock 1989).
In high-latitude shelf areas like the Ross Sea, the overwhelming dominance by a single taxonomic group is unparalleled in the fish fauna of any other marine ecosystem. As documented by benthic trawling at depths of 107–1,191 m in the southwestern Ross Sea (73–77°S), notothenioids comprise 76.6% of species, 91.6% of abundance and 91.2% of biomass (Eastman and Hubold 1999). Non-notothenioids are represented by Rajidae, Muraenolepidae, Liparidae and Zoarcidae, accounting for 8.4% of abundance and 8.5% of frequency of occurrence (Eastman and Hubold 1999). Since notothenioids are the only fish group of significant biomass, they obviously play a major role in the food web.
There is a historical or evolutionary hypothesis for this phenomenon—notothenioids reach these levels of dominance because they occupy niches filled by taxonomically diverse groups of fishes in temperate shelf areas, groups not represented on the Antarctic shelf (Eastman 2000). The fossil record indicates that over the past 40 million years, there has been a nearly complete replacement of the Antarctic fish fauna. A diverse cosmopolitan temperate fauna from the late Eocene was replaced by the highly endemic, cold-adapted modern fauna. Although the specific factors responsible for the replacement are not completely understood, the grounding of the ice sheet on the continental shelf and changing trophic conditions may have played a role. In a habitat with few other fishes and reduced competition, notothenioids underwent a depth-related diversification directed away from the ancestral benthic habitat toward pelagic or partially pelagic zooplanktivory and piscivory. Although they lack swim bladders, in a few species, density reduction to neutral buoyancy has been achieved through a combination of reduced skeletal mineralization and lipid deposition, with paedomorphosis being one of the mechanisms facilitating such a diversification (Balushkin 1984, 2000; Voskoboinikova 1982, 2001). In the dominant family Nototheniidae, about 50% of the Antarctic species temporarily or permanently inhabit the water column rather than the ancestral benthic habitat. Referred to as pelagization (Klingenberg and Ekau 1996), this evolutionary tailoring of morphology for life in the water column has been discussed by several authors (Andersen 1984; Andriashev 1965, 1987; Eastman 1993; Ekau 1991; Hubold 1992; Klingenberg and Ekau 1996) and is the hallmark of the notothenioid radiation. Again, nototheniids have received particular attention in this regard, with ecomorphological studies documenting the evolution of the pelagic or partially pelagic mode of life (Ekau 1988, 1991), later confirmed by behavioural observations in the field (Ekau and Gutt 1991; Gutt 2002; Gutt and Ekau 1996) and by feeding habits (for example, Schwarzbach 1988).
Thus, the modern notothenioid fauna of nearly 100 species forms an adaptive radiation (Clarke and Johnston 1996; Eastman 1993, 2000) and possibly a species flock on the High-Antarctic shelf (Eastman and Clarke 1998; Eastman and McCune 2000). As products of this radiation, certain water-column species reach levels of vital importance in the food web. With its density-reducing modifications, Pleuragramma antarcticum is an exemplar of a neutrally buoyant species (DeVries and Eastman 1978), and is an important subject of this review as it is the dominant notothenioid in both abundance (92%) and biomass (74–97%) in the midwaters of the Ross Sea (DeWitt 1970).
Several authors have described different pelagic and benthic feeding communities in all three ichthyofaunal subregions, focusing mainly on fish diet and feeding strategies (Casaux et al. 1990; Daniels 1982; Gröhsler 1994; Hubold and Ekau 1990; Linkowski et al. 1983; McKenna 1991; Olaso et al. 2000; Pakhomov 1997; Pakhomov and Tseitlin 1992; Pakhomov et al. 1995; Permitin and Tarverdiyeva 1972, 1978; Richardson 1975; Schwarzbach 1988; Takahashi and Iwami 1997; Targett 1981; Tarverdiyeva and Pinskaya 1980; Vacchi et al. 1999). Nevertheless, few studies have dealt with the position of fish in the Antarctic marine food web, considering fish as both prey and predator, and these studies are restricted to the Atlantic and Indian sectors of the Southern Ocean (Barrera-Oro 2002; Blankley and Grindley 1985; Hopkins et al. 1993; Hubold 1992; Williams and Duhamel 1994). Some of this work (Barrera-Oro 2002; Hopkins et al. 1993) outlines the importance of Antarctic krill (Euphausia superba) and mesopelagic fishes (myctophids) and their key role within the food web of the southern Scotia Sea marine communities.
An assessment of the role of fishes in the food web of the Ross Sea is pertinent for several reasons. As mentioned above, there is increasing recognition that the High Antarctic is a unique evolutionary site for fishes, and this has implications for the food web. In the past decade, the largest Antarctic fish, the Antarctic toothfish Dissostichus mawsoni, has become the target of a commercial longline fishery in the northern Ross Sea (Horn 2002). The harvesting of such fishery resources over next two decades is considered the major threat to the Antarctic marine ecosystem (Clarke and Harris 2003). Furthermore, the absence of the Antarctic krill over the continental shelf of the Ross Sea, where it is replaced by the ice krill (E. crystallorophias) (Biggs 1982; Hopkins 1987; Sala et al. 2002), as well as the northernmost (with respect to the Ross Sea shelf) distribution of the mesopelagic fish families Myctophidae, Gonostomatidae, Bathylagidae and Paralepididae (DeWitt 1970), raises questions concerning the composition of the marine food web of this area. Is the role of E. crystallorophias in the Ross Sea similar to that of E. superba in other Antarctic areas? What group substitutes for the absent mesopelagic families? What is the role and the relative importance of the Antarctic silverfish (P. antarcticum) in the food web of the permanent ice zone of the Ross Sea, where it is overwhelmingly the most abundant pelagic species (DeWitt 1970)? Finally, to what extent do demersal fish inhabiting the continental shelf rely on pelagic resources?
To answer these questions, we review the published literature on the feeding habits of notothenioid fish and top-level predators. We also add some new data on the diets of artedidraconids of the genera Artedidraco, Histiodraco and Pogonophryne, and the bathydraconid, Bathydraco marri. Our emphasis is on the shelf waters of the western Ross Sea, one the most intensively studied areas in the Pacific sector of the Southern Ocean.
The Ross Sea
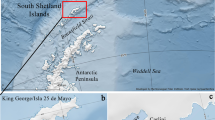
The Ross Sea is a large embayment of the Antarctic continental shelf, extending to nearly 78°S. It is surrounded by Edward VII Land, the Ross Ice Shelf and Victoria Land, with many glaciers flowing into the Ross Sea from these areas (Fig. 1). Compared to other Antarctic areas where the continental shelf is narrow, the Ross Sea has a wide shelf averaging 500 m deep, with the shelf break located at 700 m. The shelf becomes deeper from north to southwest, due to landward deepening from isostatic depression. Off Victoria Land, the shelf consists of two large N-NE shallow banks (Mawson and Pennell), about 300 m deep, separated by 500-m-deep basins. A strong cyclonic gyre characterizes the summer surface circulation on the continental shelf, with the surface current flowing westward along the Ross Ice Shelf and then northward along the Victoria Land coast (Smith and Schnack-Schiel 1990).

Map of the Ross Sea indicating most of the sites cited in the text. Dotted line shows the 500-m isobath
The Ross Sea is entirely covered by sea ice for at least 9 months of the year (Smith and Schnack-Schiel 1990). However, persistent katabatic winds off the Ross Ice Shelf (150°W–160°E) cause the formation of a large coastal polynya during spring and reduced ice concentrations in winter (Bromwich et al. 1998; Zwally et al. 1985). The front of the Ross Ice Shelf is free of sea ice during summer, and sea ice in the western part of the Ross Sea is generally advected northward into warmer water.
The fish fauna
The first ichthyological sampling in the Ross Sea was conducted during the 1839–1843 British Expedition (Richardson 1844–1848). During the first two decades of the last century, early ichthyologists described the inshore fauna off Cape Adare and McMurdo Sound, the type localities for several species of notothenioids (Boulenger 1902, 1907; Regan 1913, 1914; Waite 1911). Nearly 50 years later, between 1960 and 1980, sampling from shore and a series of cruises in the Ross Sea provided additional new species and material for taxonomic purposes (Capurro 1973; DeWitt 1962, 1964, 1970, 1971, 1985; DeWitt and Tyler 1960; Iwami and Abe 1981; Miller 1961; Miller and Reseck 1961; Reseck 1961). More recent shore- and ship-based ichthyological surveys carried out by USA and Italy (Eastman and DeVries 1982; Eastman and Hubold 1999; Vacchi et al. 1991, 1992, 1999) have greatly improved knowledge of the composition of the fish community of the Ross Sea, although the number of newly discovered species indicates our knowledge is far from complete (Chernova and Eastman 2001; Eakin and Eastman 1998; Eastman and Eakin 1999).
A fish fauna of about 50 species (Table 1) inhabits the continental shelf of the Ross Sea (i.e. the area within the shelf break at 500–700 m depth) and is composed predominantly of the endemic suborder of Notothenioidei (Eastman and Hubold 1999; Gon and Heemstra 1990; Vacchi et al. 1999). On the Ross Sea shelf, the fish fauna collected offshore with benthic trawls differs somewhat in taxonomic composition from the inshore fauna obtained by small fixed gears. This is attributable to different depth preferences of some species, although other differences are probably due to sampling artefact (Eastman and Hubold 1999). A major change occurs off Cape Adare (71–72°S), where the endemic shelf fauna receives a contribution from the mesopelagic oceanic fauna (DeWitt 1970).
The family Nototheniidae is dominant both in terms of abundance and species richness (18 species) (see Table 1). The genus Trematomus, common in the High-Antarctic Zone, is the most diverse taxon. Based on morphological characters and depth distribution (Eastman 1993), most of the nototheniids of the Ross Sea are benthic (Trematomus bernacchii, T. hansoni, T. nicolai, T. pennellii, T. scotti, T. tokarevi and Notothenia coriiceps). Given the ecomorphological trend towards pelagization mentioned previously, both epibenthic (T. eulepidotus, T. lepidorhinus and T. loennbergii) and pelagic species (Aethotaxis mitopteryx, D. mawsoni, Gvozdarus svetovidovi and P. antarcticum) are also prominent components of the shelf fauna. Furthermore, several species (Pagothenia borchgrevinki, Pagothenia brachysoma, T. newnesi) are associated with the undersurface of the ice, forming part of a cryopelagic community of organisms (Andriashev 1968, 1970).
The family Artedidraconidae, composed mostly of small benthic species (Eakin 1990), is the second most speciose group of the four families of notothenioids inhabiting the Ross Sea continental shelf. They are represented primarily by the genera Artedidraco and Pogonophryne, accounting for 88% of the total number of artedidraconid species recorded in the Ross Sea (Table 1). Among other artedidraconid genera, Dolloidraco longedorsalis is by far the most abundant species in deeper waters, whereas Histiodraco velifer represents a rare and eurybathic species (Eastman and Hubold 1999).
All members of the family Bathydraconidae occurring in East Antarctica are found in the Ross Sea shelf waters (Table 1), except for B. scotiae, which is a deep-living species inhabiting the continental slope to depths of 2,000 m. Most bathydraconids are small benthic or epibenthic fish and little is known about their biology (Gon 1990). Most species have been taken offshore with bottom trawls (Eastman and Hubold 1999; Iwami and Abe 1981), but Cygnodraco mawsoni and Gymnodraco acuticeps are the most common bathydraconids captured in inshore waters off Terra Nova Bay and McMurdo Sound (Eastman and DeVries 1982; Vacchi et al. 1999). Acanthodraco dewitti, a relatively new species from the South Shetland Islands (Skóra 1995), has been recently reported in the western Ross Sea (Vacchi et al. 2001).
The family Channichthyidae is less abundant in terms of species richness (nine species), but all the species reported from East Antarctica are found in the Ross Sea (Table 1). As a group, they are the largest notothenioids, reaching up to 75 cm TL (Iwami and Kock 1990). Most channichthyids are primarily benthic species. While Chaenodraco wilsoni, Chionodraco myersi, Dacodraco hunteri and Neopagetopsis ionah have a pelagic or partially pelagic life-style (Chen et al. 1998; Eastman 1993, 1999), most species have a combined benthic/pelagic life-style and show active vertical migration to feed on pelagic prey (Eastman and Sidell 2002; Iwami and Kock 1990; Pakhomov 1997). In the Ross Sea, Chionodraco hamatus is the most abundant and eurybathic channichthyid, both inshore and offshore (Eastman and Hubold 1999; Vacchi et al. 1999). Within the genus Cryodraco, two species, Cryodraco antarcticus and Cryodraco atkinsoni, are now recognized and they are sympatric on the continental shelf of the Ross Sea (La Mesa et al. 2002).
Prey of fish
Nototheniidae
This family is the most studied notothenioid group as far as feeding biology is concerned. Nototheniids are also diverse in terms of habitat, size, distribution and foraging modes.
In the Ross Sea, dietary data are available for the pelagic species Dissostichus mawsoni, Gvozdarus svetovidovi and Pleuragramma antarcticum. The first two are the largest species inhabiting the Southern Ocean, attaining a size of more than 200 and 100 cm, respectively (Balushkin 1994; Eastman and DeVries 2000; Yukhov 1971). The stomach of the holotype of Gvozdarus svetovidovi contains remains of Pleuragramma antarcticum, as well as bones of other fish (Balushkin 1994). Dissostichus mawsoni is predominantly piscivorous as well, feeding on Pleuragramma antarcticum, Pagothenia borchgrevinki, T. loennbergii, Gymnodraco acuticeps and Pagetopsis macropterus. However, it also relies on mysids, cephalopods and amphipods (Eastman 1985b).
As a pelagic fish, inhabiting the water column throughout its life-cycle, Pleuragramma antarcticum feeds almost exclusively on zooplankton, although it shows some dietary plasticity. Copepods (Oncaea spp., Oithona spp., Calanus propinquus, Calanoides acutus, Metridia gerlachei), both larval and adult stage of euphausiids (E. crystallorophias and E. superba), amphipods (Parathemisto gaudichaudii) and mysids (Antarctomysis ohlini) are the most abundant prey (DeWitt and Hopkins 1977; Eastman 1985a; Granata et al. 1999; Hopkins 1987; Takahashi and Nemoto 1984). Several other zooplanktonic prey, such as pelagic polychaetes (Pelagobia longicirrata), pteropods (Limacina helicina), ostracods (Conchoecia spp.), larvae of decapods and chaetognaths (Eukrohnia hamata, Sagitta spp.) are eaten occasionally (DeWitt and Hopkins 1977; Hopkins 1987). Eastman (1985a) reported that in McMurdo Sound, Pleuragramma antarcticum also feeds on fish, most of which were its own larvae (cannibalistic behaviour).
Within the cryopelagic community of the Ross Sea, feeding habits of both Pagothenia borchgrevinki and T. newnesi have been investigated in several studies. Data on Pagothenia borchgrevinki are based exclusively on specimens captured in McMurdo Sound (Eastman 1985a; Eastman and DeVries 1985; Foster and Montgomery 1993; Foster et al. 1987; Montgomery et al. 1989). This species feeds in the platelet ice and in the water beneath the ice, mainly on pteropods (L. helicina), small copepods (Calanus propinquus, Calanoides acutus, Euchaeta antarctica, M. gerlachei, Stephos longipes), gammarids (Eusirus antarcticus, Orchomene spp.) and hyperiids (Hyperiella dilatata). Secondary food consists of pelagic polychaetes, euphausiids (Euphausia crystallorophias), mysids, chaetognaths (Eukrohnia hamata) and fish (Pleuragramma antarcticum).
Feeding data are available for T. newnesi collected from both McMurdo Sound and Terra Nova Bay. The plasticity of this species is noteworthy, both in terms of body morphology and feeding habits (Eastman and DeVries 1997; La Mesa et al. 2000). In McMurdo Sound, specimens of T. newnesi sampled in different sites show some difference in diet. At Hut Point, T. newnesi was found to rely exclusively on Euphausia crystallorophias and fish (Pleuragramma antarcticum, Pagothenia borchgrevinki and Trematomus sp.), whereas at Inaccessible Island it preys on several zooplanktonic organisms (such as pteropods, copepods, hyperiids, Euphausia crystallorophias), but seemingly not on fish, even though based on few specimens (13).
Similarly, in Terra Nova Bay, the dietary composition of T. newnesi changes in relation to environmental conditions (La Mesa et al. 2000). In December, when the area is covered by sea ice, T. newnesi feeds on a few prey taxa, most characteristic of the cryopelagic habitat, such as Euphausia crystallorophias and the copepod M. gerlachei. In addition, it eats the pteropod L. helicina, other copepods (Calanoides acutus, Calanus propinquus, Euphausia antarctica) and amphipods (mainly H. dilatata and Cheirimedon fougneri). In February, when the ice cover disappears, the food spectrum of T. newnesi is more diverse, suggesting an active feeding search in the water column. Other than the above-mentioned prey, the stomach contents also reveal decapods (Chorismus antarcticus and Notocrangon antarcticus) and several species of fishes (Chionodraco sp., Pagetopsis sp., Gymnodraco acuticeps, Pagothenia borchgrevinki, Pleuragramma antarcticum, Trematomus spp.), as well as a high incidence of cannibalistic behaviour (La Mesa et al. 2000; Vacchi and La Mesa 1995).
Although they live close to the bottom, epibenthic nototheniids have a streamlined appearance and lack substrate-contact adaptations, suggesting that they are less inclined to reside directly on the substrate than other nototheniids (Eastman 1993). On the basis of the stomach contents, T. loennbergii in the Ross Sea appears more linked to the substrate than the other two epibenthic species (T. eulepidotus and T. lepidorhinus). It feeds almost exclusively on benthic organisms, such as polychaetes (mainly Eunoe anderssoni), gammarids (Eusirus spp.), isopods, decapods (Chorismus antarcticus and Notocrangon antarcticus) and fishes (Eastman 1985a; La Mesa et al. 1997).
Stomach contents of T. eulepidotus indicate that they search for food mainly in the water column, eating juvenile Euphausia. crystallorophias and Pleuragramma antarcticum (Eastman and Hubold 1999). However, amphipods (Parathemisto gaudichaudii) are the only prey of T. lepidorhinus (Takahashi and Nemoto 1984).
Generally, the benthic species of Trematomus all show high dietary diversity. They are probably on or near the bottom most of the time, displaying a series of feeding strategies such as hiding, perching, burrowing, grazing or ambushing depending on the species and bottom type. The typical benthic species, T. bernacchii, T. hansoni and T. pennellii, are all feeding generalists. They can be considered omnivorous, relying on most edible benthic organisms, such as algae, polychaetes, molluscs, gammarids, isopods, tanaids, mysids, decapods, pycnogonids, echinoderms and fishes (Eastman 1985a; Kiest 1993; La Mesa et al. 1997; Moser and Cowen 1991; Vacchi et al. 1994, 2000). However, they often ascend in the water column to feed on planktonic prey, such as pteropods, ostracods, copepods, hyperiids and euphausiids (Foster and Montgomery 1993; Montgomery et al. 1993).
Unlike the previously mentioned benthic species, T. nicolai inhabits shallow water 30–50 m deep, sometimes close to anchor ice (Eastman and Devries 1982). Its diet includes fewer polychates, fish and mysids than the benthic species, and actively moving organisms in the water column such as copepods and hyperiids (Eastman 1985a; Montgomery et al. 1993).
Artedidraconidae
Plunderfishes are probably the most sedentary notothenioids, living motionless on the sea bottom (Hubold 1991). Until recently, only one study reported data on feeding habits of this family in the Ross Sea (Wyanski and Targett 1981). Additional knowledge comes from some unpublished data (see Table 2). The barbel suggests that most artedidraconids are probably sit-and-wait predators, relying on actively moving organisms occurring either on or very close to the bottom (Wyanski and Targett 1981). The mental barbel, which characterizes this family, may be used as a lure to attract prey, although this has been documented only in Histiodraco velifer (Janssen et al. 1993). However, the barbel functions as an antenna or a sensor to perceive prey in Histiodraco velifer and Pogonophryne marmorata (Iwami et al. 1996).
Data indicated that all species feed on the same prey groups, but in very different proportions. Furthermore, changes in diet with increasing fish length were observed in Pogonophryne marmorata and Pogonophryne permitini. Most of the prey, such as errant and sedentary polychaetes, gastropods, gammarids, isopods, mysids, decapods (mostly Crangon antarcticus) and pycnogonids, are benthic or epibenthic organisms more or less linked to the substrate. However, some fish relied also on planktonic prey like copepods and cumaceans, although in small amounts (Table 2). Euphausiids and fishes were found occasionally in stomach contents of Pogonophryne permitini (Wyanski and Targett 1981).
Bathydraconidae
Of the ten species of dragonfishes in the Ross Sea (Table 1), feeding habits are known for only three benthic species (Table 2). In McMurdo Sound, Gymnodraco acuticeps is an ambush benthopelagic predator, feeding in order of importance on fishes (Pleuragramma antarcticum, Pagothenia borchgrevinki and T. nicolai), hyperiids, fish eggs and polychaetes (Eastman 1985a). In Terra Nova Bay, Cygnodraco mawsoni feeds exclusively on fishes (T. newnesi among them), but sample size is small (M. Vacchi, unpublished work). It is likely that a larger sample would indicate a more diversified diet, as has been reported in the Weddell Sea (Kock et al. 1984). We have observed Cygnodraco in aquaria and it is considerably more active than Gymnodraco. It employs a combination of subcarangiform and labriform locomotion that is probably effective in pursuit of its piscine prey.
The biology of the genus Bathydraco is practically unknown (Gon 1990), and data on feeding habits of B. marri are presented here for the first time. This species is both the most abundant and the most eurybathic bathydraconid sampled offshore in the Ross Sea (Eastman and Hubold 1999). Based on the stomach contents, B. marri can be considered a benthic species feeding mainly on crustaceans such as gammarids and mysids (Table 2). However, this fish may leave the bottom to feed in the water column on copepods and euphausiids (only one specimen of Euphausia superba).
Unfortunately, both specimens of Acanthodraco dewitti sampled off Terra Nova Bay had empty stomachs (Vacchi et al. 2001).
Channichthyidae
In general, icefishes are less dependent on the bottom for food than most other notothenioids (Eastman 1993) and there are, in fact, no obligatory benthivores among channichthyids (Voronina and Neelov 2001). Furthermore, the osteology of the jaws of channichthyids is not suited for feeding, either on small planktonic prey or for benthic browsing (Eastman 1993; Iwami 1985). Another common characteristic of this family is the very low feeding intensity; it is common for captured specimens of Chaenocephalus aceratus, Champsocephalus gunnari and Cryodraco antarcticus to have empty stomachs (Barrera-Oro et al. 1997; Kock 1981, 1985; Kock and Jones 2002). In the Ross Sea, Chaenodraco wilsoni, Chionodraco hamatus and Chionodraco myersi feed on euphausiids (Euphausia crystallorophias) and fish (Pleuragramma antarcticum and fish larvae) (Takahashi and Nemoto 1984; M. Vacchi, unpublished work). As is the case with Cryodraco antarcticus and Chaenocephalus aceratus (Kock and Jones 2002), they probably rest on the substrate waiting for fish, ascending sometimes to near-bottom or midwater layers to feed on ice krill.
Additional dietary data for other channichthyids collected in the Ross Sea are based on very few specimens (Table 2). All Cryodraco antarcticus, Dacodraco hunteri and Neopagetopsis ionah appear to be piscivorous, relying only on Pleuragramma antarcticum and fish larvae (Abe and Suzuki 1981; Eastman 1999; Takahashi and Nemoto 1984). Interestingly Dacodraco hunteri, probably the most pelagic channnichthyid, are able to feed on large specimens of Pleuragramma antarcticum, some approaching 50% of their own length (Eastman 1999).
Predators of fish
In the Ross Sea, published data on feeding habits of fish predators are available for two species of seabirds (south polar skua and Antarctic petrel), two species of penguins (Adélie penguin and emperor penguin), the Weddell seal and minke and killer whales (Table 3). However, these species are the most common birds and mammals of the Ross Sea (Saino and Guglielmo 1999). Moreover, it should be mentioned that, in the Southern Ocean, there are no fish top predators, such as sharks and large scombrids (tunas).
Among flying seabirds, the snow petrel, Pagodroma nivea, is quite common in the Ross Sea (Saino and Guglielmo 1999), but there are no data on its feeding habits. Off Adelie Land, the snow petrel largely relies on fish, which accounted for 95% of prey mass (Ridoux and Offredo 1989).
The Ross seal, Ommatophoca rossi, is the least abundant seal around Antarctica and in the Ross Sea (Erickson et al. 1983; Saino and Guglielmo1999); as a consequence, its importance in the food web of the Ross Sea should be negligible, although it feeds on Pleuragramma antarcticum elsewhere (Skinner and Klages 1994). However, the crabeater seal, Lobodon carcinophagus, is one of the most common seals in the Ross Sea (Saino and Guglielmo1999), but it consumes almost exclusively krill (Knox 1994).
Flying seabirds
Among flying seabirds, feeding habits of the south polar skua, Catharacta maccormicki, have been reported from Ross Island (Mund and Miller 1995; Young 1963). Apart from penguin eggs and chicks, the south polar skua largely relies on fishes, mainly pelagic and cryopelagic species such as Pleuragramma antarcticum and Pagothenia borchgrevinki. Interestingly, it is more piscivorous in the Antarctic Peninsula region, where it is sympatric with the brown skua (Catharacta antarctica lonnbergi), which is thought to monopolize the penguin food resource (Pietz 1987). In the Ross Sea, the sea-ice cover could negatively affect the availability of fish, as well as increase travel costs for feeding at sea, leading to the use of penguins as an additional food (Reinhardt et al. 2000).
The Antarctic petrel (Thalassoica antarctica) is one of the most abundant species within the bird community, associated with the pack ice and adjacent open waters of the Ross Sea (Ainley et al. 1984). In addition to euphausiids (Euphausia superba), the Antarctic petrel frequently consumes fish, mostly adult Pleuragramma antarcticum as large as 160 mm SL.
Penguins
Food habits and foraging strategies of Adélie penguin (Pygoscelis adeliae) have been studied at three colonies on Ross Island (Cape Royds, Cape Bird and Cape Crozier) (Ainley 2002; Ainley et al. 1998; Emison 1968; Polito et al. 2002; Van Heezik 1988). In most studies at these high-latitude sites, fish (mainly Pleuragramma antarcticum) represent a key prey item of the Adélie penguin, with euphausiids (mainly Euphausia crystallorophias) of secondary importance. However, the relative importance of fish and ice krill in their diet changes considerably in relation to environmental conditions, such as pack-ice cover and year of sampling. In some earlier studies, euphausiids appear to be the most important food of Adélie penguin on the Ross Sea shelf (Emison 1968; Van Heezik 1988). In later studies, Pleuragramma antarcticum proved to be the most important prey of Adélie penguin when little pack ice is present; conversely, juvenile Euphausia crystallorophias are predominantly eaten in years of heavy pack-ice cover (Ainley et al. 1998). On the basis of prey remains recovered from ornithogenic sediments in Ross Island, Polito et al. (2002) suggested that the importance of Pleuragramma antarcticum as food for Adélie penguins has decreased over the past 600 years. In this study, several other species of nototheniids of the genera Notothenia, Pagothenia and Trematomus were present as occasional prey (Polito et al. 2002). Sex differences in Adélie-penguin foraging strategies and diet have also been studied off Terra Nova Bay, where this species feeds both under the ice and in open water. During the nest-guard period, females make significantly longer foraging trips than males and consume larger quantities of ice krill (Euphausia crystallorophias), whereas males forage closer to the nesting areas and eat more fish (Pleuragramma antarcticum and Trematomus newnesi) (Clarke et al. 1998).
Ainley (2002) provides a comprehensive compilation of the diet of the Adélie penguin at locations around the continent. In agreement with the information above, his summary indicates that Pleuragramma antarcticum is an important component of the diet at all studied sites in East Antarctica, especially the high-latitude sites on Ross Island. He notes that the Adélie penguin is a dietary generalist; when feeding over the shelf, its diet is a mix of Pleuragramma antarcticum and Euphausia crystallorophias. However, in deep water over or north of the slope, the diet is more likely to consist of Euphausia superba and myctophid fish. Dietary plasticity is probably advantageous given the extent of the annual migration of Adélie penguins from some colonies in the Ross Sea. Studies of migratory movements using satellite telemetry indicate that some Adélie penguins breeding in the southern Ross Sea travel to over-winter in feeding grounds in dense pack ice west and north of the Balleny Islands (Davis et al. 2001). Here, the fish fauna is not well known and the Adélie diet has not been studied, but the fish fauna is probably unlike that of the southern Ross Sea given the different oceanographic and climatic regimes around the Ballenys.
Feeding habits of the emperor penguin (Aptenodytes forsteri) in the western Ross Sea have been investigated by analysis of stomach contents at Cape Washington, Coulman Island and Cape Roget (Burns and Kooyman 2001; Cherel and Kooyman 1998). At all sites, fishes are the most important food, accounting for 88–95% of diet by mass. Pleuragramma antarcticum is the most common fish prey, occurring in 95–100% of samples (Cherel and Kooyman 1998). Other fish prey includes the nototheniids (Pagothenia borchgrevinki, Trematomus lepidorhinus and Trematomus newnesi), the channichthyids (mostly Chaenodraco wilsoni and Pagetopsis sp.) and a bathydraconid (Gerlachea australis). At Coulman Island, one large specimen of the rare channichthyid Dacodraco hunteri was recorded in a regurgitation. Crustaceans are the second important taxa in the diet of emperor penguins at Cape Washington. Euphausia crystallorophias accounts for 24% of the total number of crustaceans, whereas Euphausia superba is preyed upon in lesser amounts (1%) (Cherel and Kooyman 1998).
Since emperor penguins dive to depths of 400–500 m (Kooyman and Kooyman 1995), they are able to forage in most of the water column over the Antarctic shelf, as indicated by the composition of their diet. However, the predominance of pelagic prey, such as euphausiids and juvenile Pleuragramma antarcticum, indicates that emperor penguins feed mostly in mid-water, with the majority of dives at <200 m depths (Kooyman and Kooyman 1995). During deep dives, it feeds on other nototheniids and large channichthyids, as well as on adult and subadult Pleuragramma antarcticum found at >200 m depths (Eastman 1993; Gon and Heemstra 1990). At McMurdo Sound, emperor penguins probably dive under the sea ice, feeding on cryopelagic fish such as Pagothenia borchgrevinki and Trematomus newnesi (Cherel and Kooyman 1998; Kooyman et al. 1989).
Weddell seals
Several studies consider the diet and foraging behavior of the Weddell seal (Leptonychotes weddellii) in the Ross Sea, mostly in the area of McMurdo Sound (Burns and Kooyman 2001; Burns et al. 1998; Calhaem and Christoffel 1969; Castellini et al. 1992; Davis et al. 1999; Dearborn 1965; Green and Burton 1987; Testa et al. 1985). All of these studies, based on faecal or stomach samples, indicate Weddell seals are almost exclusively piscivorous. Adult Pleuragramma antarcticum are by far the most important prey of Weddell seals, being recorded from between 70 and 100% of stomach/faecal samples. Occasionally, the Weddell seals consume some benthic species, such as Trematomus bernacchii and Trematomus pennellii (Burns et al. 1998; Castellini et al. 1992; Green and Burton 1987; Testa et al. 1985). In the Weddell Sea, Weddell seals foraged mainly on Pleuragramma antarcticum in summer (Plötz 1986), whereas in spring they fed on other species of notothenioids, such as Chionodraco myersi, Trematomus eulepidotus, Pagetopsis maculatus, Racovitzia glacialis and Cryodraco antarcticus, probably as a consequence of temporal changes in the local abundance of Pleuragramma antarcticum (Plötz et al. 1991, 2001).
Based on data from scat collections and stable-isotope analysis, Burns et al. (1998) reported two different feeding strategies in Weddell seals. Adults and most yearlings feed primarily in the water column on Pleuragramma antarcticum, while the shallow-diving yearlings forage mainly on the benthic Trematomus species. In McMurdo Sound, Weddell seals flush Pagothenia borchgrevinki, which represents an occasional prey item (Castellini et al. 1992), by blowing air into subice crevices or by pursuing them into the platelet ice (Davis et al. 1999). A video camera attached to Weddell seals as they forage under the ice of McMurdo Sound also provides some insight into previously unknown aspects of the behaviour and ecology of Pleuragramma antarcticum (Fuiman et al. 2002). Pleuragramma aggregate in loose shoals, with densities estimated as 1 fish per 7–43 m3. They also undergo diel vertical migration; they occur at mean depths of 252 m at night versus 346 m during the day. When pursued by a seal, Pleuragramma employed subcarangiform locomotion to reach an estimated speed of 4.9 body-lengths s−1 which was similar to that of other nototheniids, but slower than in temperate species. Other accounts (Calhaem and Christoffel 1969; Dearborn 1965; Wohlschlag 1968) report that Weddell seals eat large specimens of Antarctic cod (Dissostichus mawsoni), and a strict prey-predator relationship between the two species has been hypothesized (Testa et al. 1985). Using a video camera attached to Weddell seals, Fuiman et al. (2002) documented encounters and pursuit of Dissostichus mawsoni, but not captures. This interaction took place at depths of 12–180 m.
Whales
Excluding crabeater seals (Lobodon carcinophagus), the marine-mammal community of the Ross Sea is dominated in biomass by minke whales (Balaenoptera acutorostrata) (Ainley 1985). All over the Southern Ocean, they are important consumers of Antarctic krill, with estimates that, after the depletion of the large whales, they are responsible for 95% of the total biomass of krill consumed by all baleen whales in the Antarctic (Armstrong and Siegfried 1991). In the northern Ross Sea, large aggregations of minke whales are observed close to the continental shelf breaks and bank tops in proximity to high concentrations of Antarctic krill (Euphausia superba) (Ichii 1990; Kasamatsu et al. 1998). Nevertheless, in the southern part of the Ross Sea, the neritic euphausiid Euphausia crystallorophias is exclusively the dominant food species of minke whales on the shelf, as well as in the vicinity of the Bay of Whales (Ichii et al. 1998). In summer 1990–1991, Pleuragramma antarcticum was found to be an important food resource of minke whales on the southeast shelf of the Ross Sea, in addition to Euphausia crystallorophias (Ichii et al. 1998).
Finally, killer whales (Orcinus orca) are abundant off Ross Island and have been observed feeding on Antarctic cod (Dissostichus mawsoni) (Thomas et al. 1981).
Key prey species in the food web
As most of studies on feeding habits are based on the frequency of occurrence of prey, we have summarized these quantitative data for the two most likely key species, Pleuragramma antarcticum and Euphausia crystallorophias, with the aim of estimating their importance in the food web of the Ross Sea (Table 4).
With regard to the nototheniid Pleuragramma antarcticum as food for other fish, several species of channichthyids (Chionodraco hamatus, Chionodraco myersi, Dacodraco hunteri, N. ionah) rely almost exclusively on this species, although most data are based on few specimens. Similarly, the large mesopelagic predators Dissostichus mawsoni and Gvozdarus svetovidovi both feed heavily on Pleurogramma antarcticum. It is also consumed in small amounts (5–20%) by the cryopelagic species Pagothenia borchgrevinki and Trematomus newnesi, possibly when it swims close to the sea-ice cover. Top predators such as south polar skua, Adélie and emperor penguins and Weddell seal all feed on Pleuragramma antarcticum, with frequency of occurrences ranging from 26 to 100% (see Table 4). Unfortunately, no quantitative data for the diet of minke whales are available, but they probably rely on Pleuragramma antarcticum as food in periods of particular abundance and/or in areas where Euphausia superba are absent (Ichii et al. 1998).
In comparison to Pleuragramma antarcticum, the ice krill (Euphausia crystallorophias) is preyed upon by few fish species. It is found in very different amounts in the stomach contents of the channichthyids Chaenodraco wilsoni and Chaenodraco myersi (Table 4), which both frequently inhabit and feed in the water column (Eastman 1993). In addition, the ice krill has a 42–63% frequency of occurrence in the diets of cryopelagic species, such as Pagothenia borchgrevinki and Trematomus newnesi, and 20% in the zooplanktivorous Pleuragramma antarcticum.
Both Adélie and emperor penguins feed heavily on Euphausia crystallorophias (Table 4); however, the relative importance of the ice krill changes considerably in the diet of Adélie penguins both seasonally and yearly (Ainley 2002; Ainley et al. 1998). Finally, as previously mentioned, Euphausia crystallorophias represents the dominant food of minke whales on the continental shelf of the Ross Sea, although quantitative data are not available (Ichii et al. 1998).
Discussion
Compared to other Antarctic areas, the Ross Sea has several unusual environmental and biological features. The Ross Sea continental shelf is covered by sea ice for most of the year, with decay and growth starting near the ice-shelf front in late October and late February, respectively (Jacobs and Giulivi 1999). In addition, strong katabatic winds maintain two main ice-free areas, the large Ross Sea polynya close to the ice-shelf front (Zwally et al. 1985) and a small and persistent coastal polynya in the waters off Terra Nova Bay (Kurtz and Bromwich 1985). Jacobs and Comiso (1989) also describe an annually persistent polynya along the continental slope of the Ross Sea during winter.
The marginal ice zone and polynyas, as well as the continental shelf-break front, all represent key areas in terms of pelagic biological production and carbon flux to the upper trophic levels (Ainley and DeMaster 1990). In the Ross Sea, most of the top predators, such as flying seabirds, penguins, seals and minke whales, are more or less linked to the pack-ice edge, to polynyas or to the shelf-break front, where they concentrate for foraging or moulting (Ainley 1985; Ainley et al. 1984, 1990; Kasamatsu et al. 1998; Laws 1977; Saino and Guglielmo 1999; Veit and Braun 1984).
One of the most significant biological features of the Ross Sea is the absence of the Antarctic krill Euphausia superba, at least within the continental-shelf waters (Marr 1962). This is the key species in the East Wind Drift waters elsewhere in the Southern Ocean (El-Sayed 1985; Everson 1977). Instead, Euphausia crystallorophias is the dominant species over the entire Ross Sea shelf area, but it is absent north of the shelf-slope front, where it is replaced by Euphausia superba (Biggs 1982; Smith and Schnack-Schiel 1990). The distribution of the two species rarely overlaps: Euphausia crystallorophias is widespread over the continental shelf, close to the Ross Ice shelf and in neritic waters, whereas Euphausia superba is dominant offshore, from the continental slope northward (Azzali and Kalinowski 1999; Sala et al. 2002). Furthermore, data on size and sexual maturity indicate the absence of juveniles of Euphausia superba in the Ross Sea, suggesting that it is unlikely this area is used as nursery grounds by Antarctic krill (Ichii et al. 1998; Sala et al. 2002).
Other than Euphausia crystallorophias, the midwater ecosystem over the Ross Sea continental shelf is largely dominated by the nototheniid fish Pleuragramma antarcticum, representing more than 90% of both the abundance and biomass of midwater fish fauna (DeWitt 1970). In addition, as a consequence of the sharp decrease in temperature of deep water (from +0.5 to −1.8°C) in the northern area of the Ross Sea, almost all mesopelagic myctophids, gonostomatids, bathylagids and paralepidids are excluded from the continental shelf (DeWitt 1970).
With these considerations in mind, the traditional concept of the Antarctic food web as a simple chain consisting of phytoplankton, krill and consumers of krill (Tranter 1982) is not valid in the high-latitude shelf waters. In particular, on the basis of the present analysis and previous studies, the food web is non-linear with complex prey-predator interactions in the shelf waters of the Ross Sea (Fig. 2).

Diagram showing the position of notothenioid fish in the food web of the Ross Sea continental shelf
In an area where Antarctic krill is scarce or absent, such as the Ross Sea shelf, the key role of midwater fish (particularly Pleuragramma antarcticum) and the neritic euphausiid Euphausia crystallorophias is documented or hypothesized in many studies on the feeding habits of fish and top predators such as sea birds, penguins, seals and whales (Ainley et al. 1998; Burns et al. 1998; Cherel and Kooyman 1998; Eastman 1985a, 1985b; Hempel 1985; Hopkins 1987; Ichii et al. 1998; Takahashi and Nemoto 1984; Testa et al. 1985). Considering all data published to date, we now have a reasonably detailed view of the structure of the marine food web of the Ross Sea continental shelf, especially the role of certain notothenioid fish.
Information on feeding habits and life-style indicates that Antarctic notothenioids are primarily benthic (see Table 2), and this is likely the ancestral condition as well. However, an evolutionary departure from the benthic habitat occurred in several species, which became pelagic, cryopelagic or epibenthic (Eastman 1993). In the Southern Ocean, the notothenioids, and particularly the nototheniids, fill a wide variety of niches, so that they can be considered the equivalent of the taxonomically and ecologically diverse fish faunas from other shelf areas of the world. The phenomenon of pelagization (Klingenberg and Ekau 1996) allowed several nototheniid species to depart from the bottom and to live and feed in the water column. The same is also true for some channichthyids. The absence of competition from mesopelagic species on the high-latitude continental shelves of Antarctica probably facilitated this trend. The notothenioids permanently inhabiting the water column, such as pelagic or cryopelagic species, became the most important food for several top predators, as discussed below.
The position of notothenioid fish within the marine food web of the Ross Sea shelf waters is summarized in Fig. 2. Of the 31 fish species considered, 19 are not preyed upon by any of the top predators. Most of these are small benthic or epibenthic fish (see Table 2), which may not be easily available to predators foraging in deep waters near the bottom, such as Weddell seals and emperor penguins (Kooyman and Kooyman 1995; Plötz et al. 2001). Based on the analysis of stomach contents, it is also noteworthy that almost all benthic notothenioid species feed on zooplanktonic organisms (Fig. 2, Table 2). As an example, planktivory in benthic nototheniid fish has been reported in McMurdo Sound (Foster and Montgomery 1993), where Trematomus bernacchi, Trematomus hansoni and Trematomus pennellii rely in varying degrees on copepods (M. gerlachei), pteropod molluscs (Limacina helicina) and hyperiids (Hyperiella dilatata), which are the most abundant species in the midwater food web of this area (Hopkins 1987). Most benthic nototheniids can, therefore, be considered as opportunistic feeders, undertaking occasional vertical migrations to forage on locally and seasonally abundant planktonic prey.
At the top of the food web, predators feed on relatively few species of notothenioids, most of which are pelagic, cryopelagic or epibenthic (Fig. 2). In particular, the cryopelagic Pagothenia borchgrevinki and Trematomus newnesi both represent significant prey for Weddell seals and emperor penguins (Castellini et al. 1992; Cherel and Kooyman 1998): their role as a link between zooplankton and top predators may be more important than previously thought. A similar case can be made for Dissostichus mawsoni, whose importance in the diet of Weddell seals and killer whales has probably been underestimated.
Pleuragramma antarcticum, however, dominates the diet of all top predators, from the Antarctic petrel to the Weddell seal (Fig. 2, Table 4). In the southeast shelf of the Ross Sea, Pleuragramma antarcticum is preyed upon heavily by minke whales as well (Ichii et al. 1998). Moreover, at McMurdo Sound, they represent the principal food item of several fishes, including pelagic and cryopelagic nototheniids and some channichthyids (Table 4; Eastman 1985a).
Interestingly, compared to Pleuragramma antarcticum, a smaller number of predators rely on Euphausia crystallorophias (Table 4), which is less important as prey than the Antarctic krill in other Antarctic areas. In the Ross Sea, ice krill is consumed in large amounts only by few nototheniids (Pagothenia borchgrevinki and Trematomus newnesi), one icefish (Chaenodraco wilsoni) and Adélie and emperor penguins. Finally, Euphausia crystallorophias is the dominant species in the diet of minke whales, although only when foraging on the continental shelf (Ichii et al. 1998).
In conclusion, notothenioid fish play a key role in the high-Antarctic food web of the Ross Sea: as predators, they occupy most of the trophic niches available in the marine ecosystem, relying on benthic, zooplanktonic and nektonic organisms; as prey, they are important food resources for each other and most of the top predators living and foraging on the shelf. They also constitute the major link between lower (invertebrates) and higher (birds and mammals) levels of the food web. This is especially true for Pleuragramma antarcticum. Along with Euphausia crystallorophias, its ecological role in the Ross Sea is equivalent to that of myctophids and Euphausia superba elsewhere in the Southern Ocean.
References
Abe T, Suzuki M (1981) Notes on some fishes associated with the Antarctic krill. 2. On Xenocyttus nemotoi Abe and again on Neopagetopsis ionah Nybelin. Antarct Rec 71:121–129
Ainley DG (1985) Biomass of birds and mammals in the Ross Sea. In: Siegfried WR, Condy PR, Laws RM (eds) Antarctic nutrient cycles and food webs. Proc. 4th SCAR symposium on Antarctic biology. Springer, Berlin, pp 498–515
Ainley DG (2002) The Adélie penguin: bellwether of climate change. Columbia University Press, New York
Ainley DG, DeMaster DP (1990) The upper trophic levels in polar marine ecosystems. In: Smith WO (ed) Polar oceanography, Part B. chemistry, biology and geology. Academic, San Diego, pp 599–630
Ainley DG, O’Connor EF, Boekelheide RJ (1984) Marine ecology of birds in the Ross Sea, Antarctica. Ornithol Monogr 32:1–97
Ainley DG, Ribic CA, Wood RC (1990) A demographic study of the south polar skua Catharacta maccormicki at Cape Crozier. J Anim Ecol 59:1–20
Ainley DG, Wilson PR, Barton KJ, Ballard G, Nur N, Karl B (1998) Diet and foraging effort of Adélie penguins in relation to pack-ice conditions in the southern Ross Sea. Polar Biol 20:311–319
Andersen NC (1984) Genera and subfamilies of the family Nototheniidae (Pisces, Perciformes) from the Antarctic and Subantarctic. Steenstrupia 10:1–34
Anderson JB (1999) Antarctic marine geology. Cambridge University Press, Cambridge
Andriashev AP (1965) A general review of the Antarctic fish fauna. In: Oye P van, Mieghem J van (ed) Biogeography and ecology in Antarctica, Monographiae biologicae, vol XV. Junk, The Hague, pp 491–550
Andriashev AP (1968) The problem of the life community associated with the Antarctic fast ice. In: Currie RI (ed) Symposium on Antarctic oceanography. Scott Polar Research Institute, Cambridge, pp 147–155
Andriashev AP (1970) Cryopelagic fishes of the Arctic and Antarctic and their significance in polar ecosystems. In: Holdgate MW (ed) Antarctic ecology, vol 1. Academic, London, pp 297–304
Andriashev AP (1987) A general review of the Antarctic bottom fish fauna. In: Kullander SO, Fernholm B (eds) Proceedings of the fifth congress of European ichthyologists. Swedish Museum of Natural History, Stockholm, pp 357–372
Armstrong AJ, Siegfried WR (1991) Consumption of Antarctic krill by minke whales. Antarct Sci 3:13–18
Azzali M, Kalinowski J (1999) Spatial and temporal distribution of krill Euphausia superba biomass in the Ross Sea (1989–1990 and 1994). In: Faranda F, Guglielmo L, Ianora A (eds) Ross Sea ecology. Italiantartide Expeditions (1987–1995). Springer, Berlin Heidelberg New York, pp 434–455
Balushkin AV (1984) Morphological bases of the systematics and phylogeny of the nototheniid fishes. Acad Sci USSR Zool Inst Leningrad 1–140
Balushkin AV (1994) Gvozdarus svetovidovi gen. et sp. n. (Pisces, Nototheniidae) from Ross Sea (Antarctica). J Ichthyol 34:152–158
Balushkin AV (2000) Morphology, classification, and evolution of notothenioid fishes of the Southern Ocean (Notothenioidei, Perciformes). J Ichthyol 40 [Suppl 1]:74–109
Barrera-Oro E (2002) Review: the role of fish in the Antarctic marine food web: differences between inshore and offshore waters in the southern Scotia Arc and west Antarctic Peninsula. Antarct Sci 14:293–309
Barrera-Oro E, Casaux R, Roux A (1997) Diet composition of Champsocephalus gunnari (Pisces, Channichthyidae) around South Georgia Islands, R/V Eduardo L. Homberg survey, February/March 1994. Rev Invest Desarr Pesq 11:53–62
Biggs DC (1982) Zooplankton excretion and NH4 + cycling in near-surface waters of the Southern Ocean. I. Ross Sea, austral summer 1977–1978. Polar Biol 1:55–67
Blankley WO, Grindley JR (1985) The intertidal and shallow subtidal food web at Marion Island. In: Siegfried WR, Condy PR, Laws RM (eds) Antarctic nutrient cycles and food webs. Proc 4th SCAR symposium on Antarctic biology. Springer, Berlin Heidelberg New York, pp 630–636
Boulenger GA (1902) Pisces. Report on the collections of natural history made in Antarctic regions during the voyage of the “Southern Cross”. British Museum of Natural History, London
Boulenger GA (1907) Fishes. National Antarctic Expedition 1901–1904. Nat Hist Zool 2:1–5
Bromwich DH, Liu Z, Rogers AN, Van Woert ML (1998) Winter atmospheric forcing of the Ross Sea Polynya. In: Jacobs SS, Weiss RF (eds) Ocean, ice and atmosphere: interactions at the Antarctic continental margin. Antarct Res Ser 75. American Geophysical Union, Washington, DC, pp 101–133
Burns JM, Kooyman GL (2001) Habitat use by Weddell seals and emperor penguins foraging in the Ross Sea, Antarctica. Am Zool 41:90–98
Burns JM, Trumble SJ, Castellini MA, Testa JW (1998) The diet of Weddell seals in McMurdo Sound, Antarctica as determined from scat collections and stable isotope analysis. Polar Biol 19:272–282
Calhaem I, Christoffel DA (1969) Some observations of the feeding habits of a Weddell seal, and measurements of its prey, Dissostichus mawsoni, at McMurdo Sound, Antarctica. NZ J Freshwater Res 3:181–190
Capurro LRA (1973) USNS Eltanin’s 55 cruises—scientific accomplishments. Antarct J US 8:57–61
Casaux RJ, Mazzotta AS, Barrera-Oro ER (1990) Seasonal aspects of the biology and diet of nearshore nototheniid fish at Potter Cove, South Shetland Islands, Antarctica. Polar Biol 11:63–72
Castellini MA, Davis RW, Kooyman GL (1992) Annual cycles of diving behavior and ecology of the Weddell seal. Bull Scripps Inst Oceanogr 28:1–54
Chen WJ, Bonillo C, Lecointre G (1998) Phylogeny of the Channichthyidae (Notothenioidei, Teleostei) based on two mitochondrial genes. In: Prisco G di, Pisano E, Clarke A (eds) Fishes of Antarctica: a biological overview. Springer, Milan, pp 287–298
Cherel Y, Kooyman GL (1998) The food of emperor penguins (Aptenodytes forsteri) in the western Ross Sea (Antarctica). Mar Biol 130:335–344
Chernova NV, Eastman JT (2001) Two new species of snailfish genus Paraliparis (Pisces: Liparidae) from the Ross Sea, Antarctica. J Fish Biol 59:92–104
Clarke A, Harris CM (2003) Polar marine ecosystems: major threats and future change. Environ Conserv 30:1–25
Clarke A, Johnston IA (1996) Evolution and adaptive radiation of Antarctic fishes. Trends Ecol Evol 11:212–218
Clarke J, Manly B, Kerry K, Gardner H, Franchi E, Corsolini S, Focardi S (1998) Sex differences in Adélie penguin foraging strategies. Polar Biol 20:248–258
Daniels RA (1982) Feeding ecology of some fishes of the Antarctic Peninsula. Fish Bull US 80:575–588
Davis LS, Harcourt RG, Bradshaw CJA (2001) The winter migration of Adelie penguins breeding in the Ross Sea sector of Antarctica. Polar Biol 24:593–597
Davis RW, Fuiman LA, Williams TM, Collier SO, Hagey WP, Kanatous SB, Kohin S, Horning M (1999) Hunting behavior of a marine mammal beneath the Antarctic fast ice. Science 283:993–996
Dearborn JH (1965) Food of Weddell seals at McMurdo Sound, Antarctica. J Mammal 46:37–43
DeVries AL, Eastman JT (1978) Lipid sacs as a buoyancy adaptation in an Antarctic fish. Nature 271:352–353
DeWitt HH (1962) A new Antarctic nototheniid fish with notes on two recently described nototheniiformes. Copeia 4:826–833
DeWitt HH (1964) A revision of the Antarctic genus Racovitzia (Pisces, Bathydraconidae). Copeia 3:496–506
DeWitt HH (1970) The character of the midwater fish fauna of the Ross Sea, Antarctica. In: Holdgate MW (ed) Antarctic ecology, vol 1. Academic, London, pp 305–314
DeWitt HH (1971) Coastal and deep-water benthic fishes of the Antarctic. In: Bushnell VC (ed) Antarctic Map Folio Series 15. American Geographical Society, New York, pp 1–10
DeWitt HH (1985) Reports on fishes of the University of Southern California Antarctic Research Program, 1962–1968. 1. A review of the genus Bathydraco Günther (Family Bathydraconidae). Cybium 9:295–314
DeWitt HH, Hopkins TL (1977) Aspects of the diet of the antarctic silverfish, Pleuragramma antarcticum. In: Llano GA (ed) Adaptations within Antarctic ecosystems. Proc 3rd SCAR symposium on Antarctic biology. Smithsonian Institution, Washington, DC, pp 557–567
DeWitt HH, Tyler JC (1960) Fishes of the Stanford Antarctic Biological Research Program, 1959–1959. Stanford Ichthyol Bull 7:162–199
Eakin RR (1990) Artedidraconidae. In: Gon O, Heemstra PC (eds) Fishes of the Southern Ocean. JLB Smith Institute of Ichthyology, Grahamstown, pp 332–356
Eakin RR, Eastman JT (1998) New species of Pogonophryne (Pisces, Artedidraconidae) from the Ross Sea, Antarctica. Copeia 4:1005–1009
Eastman JT (1985a) Pleuragramma antarcticum (Pisces, Nototheniidae) as food for other fishes in McMurdo Sound, Antarctica. Polar Biol 4:155–160
Eastman JT (1985b) The evolution of neutrally buoyant Notothenioid fishes: their specializations and potential interactions in the Antarctic marine food web. In: Siegfried WR, Condy PR, Laws RM (eds) Antarctic nutrient cycles and food webs. Proc 4th SCAR symposium on Antarctic biology. Springer, Berlin Heidelberg New York, pp 430–436
Eastman JT (1993) Antarctic fish biology: evolution in an unique environment. Academic, San Diego
Eastman JT (1999) Aspects of the biology of the icefish Dacodraco hunteri (Notothenioidei, Channichthyidae) in the Ross Sea, Antarctica. Polar Biol 21:194–196
Eastman JT (2000) Antarctic notothenioid fishes as subjects for research in evolutionary biology. Antarct Sci 12:276–287
Eastman JT, Clarke A (1998) A comparison of adaptive radiations of Antarctic fish with those of non-Antarctic fish. In: Prisco G di, Pisano E, Clarke A (ed) Fishes of Antarctica: a biological overview. Springer, Milan, pp 3–26
Eastman JT, DeVries AL (1982) Buoyancy studies of Notothenioid fishes in McMurdo Sound, Antarctica. Copeia 2:385–393
Eastman JT, DeVries AL (1985) Adaptations for cryopelagic life in the Antarctic notothenioid fish Pagothenia borchgrevinki. Polar Biol 4:45–52
Eastman JT, DeVries AL (1997) Biology and phenotypic plasticity of the Antarctic nototheniid fish Trematomus newnesi in McMurdo Sound. Antarct Sci 9:27–35
Eastman JT, DeVries AL (2000) Aspects of body size and gonadal histology in the Antarctic toothfish, Dissostichus mawsoni, from McMurdo Sound, Antarctica. Polar Biol 23:189–195
Eastman JT, Eakin RR (1999) Fishes of the genus Artedidraco (Pisces, Artedidraconidae) from the Ross Sea, Antarctica, with the description of a new species and a colour morph. Antarct Sci 11:13–22
Eastman JT, Eakin RR (2000) An updated species list for notothenioid fish (Perciformes; Notothenioidei), with comments on Antarctic species. Arch Fish Mar Res 48:11–20
Eastman JT, Hubold G (1999) The fish fauna of the Ross Sea, Antarctica. Antarct Sci 11:293–304
Eastman JT, McCune AR (2000) Fishes on the Antarctic continental shelf: evolution of a marine species flock? J Fish Biol 57 [Suppl A]:84–102
Eastman JT, Sidell BD (2002) Measurements of buoyancy for some Antarctic notothenioid fishes from the South Shetland Islands. Polar Biol 25:753–760
Ekau W (1988) Ecomorphology of nototheniid fish from the Weddell Sea, Antarctica. Ber Polarforsch 51:1–140
Ekau W (1991) Morphological adaptations and mode of life in High Antarctic fish. In: Prisco G di, Maresca B, Tota B (eds) Biology of Antarctic fish. Springer, Berlin Heidelberg New York, pp 23–29
Ekau W, Gutt J (1991) Notothenioid fishes from the Weddell Sea and their habitat, observed by underwater photography and television. Proc NIPR Symp Polar Biol 4:36–49
El-Sayed SZ (1985) Plankton of the Antarctic seas. In: Bonner WN, Walton DWH (eds) Antarctica. Pergamon, Oxford, pp 135–153
Emison WB (1968) Food preferences of Adélie penguin at Cape Crozier, Ross Island. Antarct Res Ser 12:191–212
Erickson AW, Hanson MB, Kehoe DM (1983) Population densities of seals and whales observed during the 1983 circumnavigation of Antarctica by the USCGC Polar Star. Antarct J US 18:163–166
Everson I (1977) The living resources of the Southern Ocean. (GLO/SO/77/1). FAO, Rome
Foster BA, Montgomery JC (1993) Planktivory in benthic nototheniid fish in McMurdo Sound, Antarctica. Environ Biol Fish 36:313–318
Foster BA, Cargill JM, Montgomery JC (1987) Planktivory in Pagothenia borchgrevinki (Pisces: Nototheniidae) in McMurdo Sound, Antarctica. Polar Biol 8:49–54
Fuiman LA, Davis RW, Williams TM (2002) Behavior of midwater fishes under Antarctic ice: observations by a predator. Mar Biol 140:815–822
Gon O (1990) Bathydraconidae. In: Gon O, Heemstra PC (eds) Fishes of the Southern Ocean. JLB Smith Institute of Ichthyology, Grahamstown, pp 364–380
Gon O, Heemstra PC (eds) (1990) Fishes of the Southern Ocean. JLB Smith Institute of Ichthyology, Grahamstown
Granata A, Guglielmo L, Greco S, Vacchi M, Sidoti O, Zagami G, La Mesa M (1999) Spatial distribution and feeding habits of larval and juvenile Pleuragramma antarcticum in the western Ross Sea (Antarctica). In: Faranda F, Guglielmo L, Ianora A (eds) Ross Sea ecology. Italiantartide Expeditions (1987–1995). Springer, Berlin Heidelberg New York, pp 369–393
Green K, Burton HR (1987) Seasonal and geographical variation in the food Weddell seals, Leptonychotes weddellii, in Antarctica. Aust Wildl Res 14:475–489
Gröhsler T (1994) Feeding habits as indicators of ecological niches: investigations of Antarctic fish conducted near Elephant Island in late autumn/winter 1986. Arch Fish Mar Res 42:17–34
Gutt J (2002) The Antarctic ice shelf: an extreme habitat for notothenioid fish. Polar Biol 25:320–322
Gutt J, Ekau W (1996) Habitat partitioning of dominant high Antarctic demersal fish in the Weddell Sea and Lazarev Sea. J Exp Mar Biol Ecol 206:25–37
Hempel G (1985) Antarctic marine food webs. In: Siegfried WR, Condy PR, Laws RM (eds) Antarctic nutrient cycles and food webs. Proc. 4th SCAR symposium on Antarctic biology. Springer, Berlin Heidelberg New York, pp 266–270
Hopkins TL (1987) Midwater food web in McMurdo Sound, Ross Sea, Antarctica. Mar Biol 96:93–106
Hopkins TL, Ainley DG, Torres JJ, Lancraft TM (1993) Trophic structure in open waters of the marginal ice zone in the Scotia-Weddell confluence region during spring (1983). Polar Biol 13:389–397
Horn PL (2002) Age and growth of Patagonian toothfish (Dissostichus eleginoides) and Antarctic toothfish (D. mawsoni) in waters from the New Zealand subantarctic to the Ross Sea, Antarctica. Fish Res 56:275–287
Hubold G (1991) Ecology of notothenioid fish in the Weddell Sea. In: Prisco G di, Maresca B, Tota B (eds) Biology of Antarctic fish. Springer, Berlin Heidelberg New York, pp 3–22
Hubold G (1992) Ecology of Weddell Sea fishes. Ber Polarforsch 103:1–157
Hubold G, Ekau W (1990) Feeding patterns of post-larval and juvenile Notothenioids in the Southern Weddell Sea (Antarctica). Polar Biol 10:255–260
Hureau JC (1994) The significance of fish in the marine Antarctic ecosystems. Polar Biol 14:307–313
Ichii T (1990) Distribution of antarctic krill concentrations exploited by Japanese krill trawlers and minke whales. Proc NIPR Symp Polar Biol 3:36–56
Ichii T, Shinohara N, Fujise Y, Nishiwaki S, Matsuoka K (1998) Interannual changes in body fat condition index of minke whales in the Antarctic. Mar Ecol Prog Ser 175:1–12
Iwami T (1985) Osteology and relationships of the family Channichthyidae. Mem Natl Inst Polar Res Ser E 36:1–69
Iwami T, Abe T (1981) The collection of the fishes trawled in the Ross Sea. Antarct Rec 71:130–141
Iwami T, Kock K-H (1990) Channichthyidae. In: Gon O, Heemstra PC (eds) Fishes of the Southern Ocean. JLB Smith Institute of Ichthyology, Grahamstown, pp 381–399
Iwami T, Numanami H, Naito Y (1996) Behavior of three species of the family Artedidraconidae (Pisces, Notothenioidei), with reference to feeding. Proc NIPR Symp Polar Biol 9:225–230
Jacobs SS, Comiso JC (1989) Sea ice and oceanic processes on the Ross Sea continental shelf. J Geophys Res 94:18195–18211
Jacobs SS, Giulivi CF (1999) Thermohaline data and ocean circulation on the Ross Sea continental shelf. In: Spezie G, Manzella GMR (eds) Oceanography of the Ross Sea, Antarctica. Springer, Milan, pp 3–16
Janssen J, Montgomery JC, Tien R (1992) Social rotational feeding in Pagothenia borchgrevinki (Pisces: Nototheniidae). Copeia 2:559–562
Janssen J, Jones W, Slattery M (1993) Locomotion and feeding responses to mechanical stimuli in Histiodraco velifer (Artedidraconidae). Copeia 3:885–889
Kasamatsu F, Ensor P, Joyce GG (1998) Clustering and aggregations of minke whales in the Antarctic feeding grounds. Mar Ecol Prog Ser 168:1–11
Kiest KA (1993) A relationship of diet to prey abundance and the foraging behavior of Trematomus bernacchii. Polar Biol 13:291–296
Klingenberg CP, Ekau W (1996) A combined morphometric and phylogenetic analysis of an ecomorphological trend: pelagization in Antarctic fishes (Perciformes: Nototheniidae). Biol J Linn Soc 59:143–177
Knox GA (1994) The biology of the Southern Ocean. Cambridge University Press, Cambridge
Kock K-H (1981) Fischereibiologische Untersuchungen an drei antarktischen Fischarten: Champsocephalus gunnari Lönnberg, 1905, Chaenocephalus aceratus (Lönnberg, 1906) und Pseudochaenichthys georgianus Norman, 1937 (Notothenioidei, Channichthyidae). Mitt Inst Seefisch Hamb 32:1–226
Kock K-H (1985) Krill consumption by Antarctic notothenioid fish. In: Siegfried WR, Condy PR, Laws RM (eds) Antarctic nutrient cycles and food webs. Proc. 4th SCAR symposium on Antarctic biology, Springer, Berlin Heidelberg New York, pp 437–444
Kock K-H (1992) Antarctic fish and fisheries. Cambridge University Press, Cambridge
Kock K-H, Jones CD (2002) The biology of the icefish Cryodraco antarcticus Dollo, 1900 (Pisces, Channichthyidae) in the southern Scotia Arc (Antarctica). Polar Biol 25:416–424
Kock K-H, Schneppenheim R, Siegel V (1984) A contribution to the fish fauna of the Weddell Sea. Arch FischWiss 34:103–120
Kooyman GL, Kooyman TG (1995) Diving behavior of emperor penguins nurturing chicks at Coulman Island, Antarctica. Condor 97:536–549
Kooyman GL, Ponganis PJ, Castellini MA, Ponganis E, Eckert S, Le Maho Y (1989) Physiology of diving in emperor penguins and Weddell seals. Antarct J US 24:204
Kurtz DD, Bromwich DH (1985) A recurring, atmospherically forced polynya in Terra Nova Bay. In: Jacobs SS (ed) Oceanology of the Antarctic continental shelf. Antarct Res Ser 43. American Geophysical Union, Washington DC, pp 177–201
La Mesa M, Vacchi M, Castelli A, Diviacco G (1997) Feeding ecology of two nototheniid fishes, Trematomus hansoni and Trematomus loennbergii, from Terra Nova Bay, Ross Sea. Polar Biol 17:62–68
La Mesa M, Vacchi M, Zunini Sertorio T (2000) Feeding plasticity of Trematomus newnesi (Pisces, Nototheniidae) in Terra Nova Bay, Ross Sea, in relation to environmental conditions. Polar Biol 23:38–45
La Mesa M, Vacchi M, Iwami T, Eastman JT (2002) Taxonomic studies of the Antarctic icefish genus Cryodraco Dollo, 1900 (Notothenioidei: Channichthyidae). Polar Biol 25:384–390
Laws RM (1977) The significance of vertebrates in the Antarctic marine ecosystems. In: Llano GA (ed) Adaptations within Antarctic ecosystems. Proc 3rd SCAR symposium on Antarctic biology. Smithsonian Institution, Washington DC, pp 411–438
Linkowski TB, Presler P, Zukowski C (1983) Food habits of nototheniid fishes (Nototheniidae) in Admiralty Bay (King George Island, South Shetland Islands). Pol Polar Res 4:79–95
Lutjeharms JRE (1990) The oceanography and fish distribution of the Southern Ocean. In: Gon O, Heemstra PC (eds) Fishes of the Southern Ocean. JLB Smith Institute of Ichthyology, Grahamstown, pp 6–27
Marr JWS (1962) The natural history and geography of the Antarctic krill (Euphausia superba Dana). Discovery Rep 32:33–464
McKenna JE (1991) Trophic relationships within the Antarctic demersal fish community of South Georgia Island. Fish Bull US 89:643–654
Miller RG (1961) Notothenioid fishes from Cape Hallet and Ross Sea, Antarctica. N Z J Sci 4:664–668
Miller RG, Reseck R (1961) Chionodraco markami, a new Antarctic fish of the family Chaenichthyidae. Copeia 1:50–53
Montgomery JC, Foster BA, Cargill JM (1989) Stomach evacuation rate in the planktivorous Antarctic fish Pagothenia borchgrevinki. Polar Biol 9:405–408
Montgomery JC, Foster BA, Milton RC, Carr E (1993) Spatial and temporal variations in the diet of nototheniid fish in McMurdo Sound, Antarctica. Polar Biol 13:429–431
Moser M, Cowen RK (1991) Effects of periodic eutrophication on parasitism and stock identification of Trematomus bernacchii (Pisces: Nototheniidae) in McMurdo Sound, Antarctica. J Parasitol 77:551–556
Mund MJ, Miller GD (1995) Diet of the south polar skua Catharacta maccormicki at Cape Bird, Ross Island, Antarctica. Polar Biol 15:453–455
Olaso I, Rauschert M, De Broyer C (2000) Trophic ecology of the family Artedidraconidae (Pisces: Osteichthyes) and its impact on the eastern Weddell Sea benthic system. Mar Ecol Prog Ser 194:143–158
Pakhomov EA (1997) Feeding and exploitation of the food supply by demersal fishes in the Antarctic Part of the Indian Ocean. J Ichthyol 37:360–380
Pakhomov EA, Tseitlin VB (1992) Feeding patterns of nine species of antarctic fish and assessment of their daily food consumption. Sel Sci Pap CCAMLR 1991:321–333
Pakhomov EA, Pankratov SA, Gorelova TA (1995) Food web of juvenile Antarctic fish. Oceanology 34:521–532
Permitin YY, Tarverdiyeva MI (1972) The food of some Antarctic fish in the South Georgia area. J Ichthyol 12:104–114
Permitin YY, Tarverdiyeva MI (1978) Feeding of fishes of the families Nototheniidae and Chaenichthyidae in the South Orkney Islands. Sov J Mar Biol 4:619–622
Pietz PC (1987) Feeding and nesting ecology of sympatric South Polar and brown skuas. Auk 104:617–627
Plötz J (1986) Summer diet of Weddell seals (Leptonychotes weddellii) in the eastern and southern Weddell Sea, Antarctica. Polar Biol 6:97–102
Plötz J, Ekau W, Reijnders PJH (1991) Diet of Weddell seals Leptonychtes weddellii at Vestkapp, eastern Weddell Sea (Antarctica), in relation to local food supply. Mar Mammal Sci 7:136–144
Plötz J, Bornemann H, Knust R, Schröder A, Bester M (2001) Foraging behaviour of Weddell seals, and its ecological implications. Polar Biol 24:901–909
Polito M, Emslie SD, Walker W (2002) A 1000-year record of Adélie penguin diets in the southern Ross Sea. Antarct Sci 14:327–332
Regan CT (1913) The Antarctic fishes of the Scottish National Antarctic Expedition. Trans R Soc Edinburgh 49:229–292
Regan CT (1914) Fishes. British Antarctic (“Terra Nova”) Expedition 1910, Natural History Report. Zoology 1:1–54
Reinhardt K, Hahn S, Peter HU, Wemhoff H (2000) A review of the diets of Southern Hemisphere skuas. Mar Ornithol 28:7–19
Reseck J (1961) A note on fishes from the Ross Sea, Antarctica. N Z J Sci 4:107–115
Richardson J (1844–48) Ichthyology. In: Richardson J, Gray JS (eds) The zoology of the voyage of HMS Erebus and Terror under the command of Captain Sir James Clark Ross during 1839–43. Janson, London, pp 1–139
Richardson MG (1975) The dietary composition of some Antarctic fish. Br Antarct Surv Bull 41–42:113–120
Ridoux V, Offredo C (1989) The diets of five summer breeding seabirds in Adelie Land, Antarctica. Polar Biol 9:137–145
Saino N, Guglielmo L (1999) ROSSMIZE Expedition: distribution and biomass of birds and mammals in the western Ross Sea. In: Faranda F, Guglielmo L, Ianora A (eds) Ross Sea ecology. Italiantartide Expeditions (1987–1995). Springer, Berlin Heidelberg New York, pp 469–478
Sala A, Azzali M, Russo A (2002) Krill of the Ross Sea: distribution, abundance and demography of Euphausia superba and Euphausia crystallorophias during the Italian Antarctic Expedition (January-February 2000). Sci Mar 66:123–133
Schwarzbach W (1988) The demersal fish fauna of the eastern and southern Weddell Sea: geographical distribution, feeding of fishes and their trophic position in the food web. Ber Polarforsch 54:1–94
Skinner JD, Klages NTW (1994) On some aspects of the biology of the Ross seal Ommatophoca rossii from King Haakon VII Sea, Antarctica. Polar Biol 14:467–472
Skóra KE (1995) Acanthodraco dewitti gen. et sp. n. (Pisces, Bathydraconidae) from Admiralty Bay (King George Island, South Shetland Islands, Antarctica). Arch Fish Mar Res 42:283–289
Smith SL, Schnack-Schiel (1990) Polar zooplankton. In: Smith WO (ed) Polar oceanography, part B: chemistry, biology and geology. Academic, San Diego, pp 527–598
Takahashi M, Iwami T (1997) The summer diet of demersal fish at the South Shetland Islands. Antarct Sci 9:407–413
Takahashi M, Nemoto T (1984) The food of some Antarctic fish in the western Ross Sea in summer 1979. Polar Biol 3:237–239
Targett TE (1981) Trophic ecology and structure of coastal Antarctic fish communities. Mar Ecol Prog Ser 4:243–263
Tarverdiyeva MI, Pinskaya IA (1980) The feeding of fishes of the families Nototheniidae and Chaenichthyidae on the shelves of the Antarctic Peninsula and the South Shetlands. J Ichthyol 20:50–60
Testa JW, Siniff DB, Ross MJ, Winter JD (1985) Weddell seal—Antarctic cod interactions in McMurdo Sound, Antarctica. In: Siegfried WR, Condy PR, Laws RM (eds) Antarctic nutrient cycles and food webs. Proc 4th SCAR Symposium on Antarctic biology, Springer, Berlin Heidelberg New York, pp 561–565
Thomas JA, Leatherwood S, Evans WE, Jehl JR, Awbrey FT (1981) Ross Sea killer whale distribution, behavior, color pattern, and vocalizations. Antarct J US 16:157–158
Tiedtke JE, Kock K-H (1989) Structure and composition of the demersal fish fauna around Elephant Island. Arch FischWiss 39:143–169
Tranter DJ (1982) Interlinking of physical and biological processes in the Antarctic ocean. Oceanogr Mar Biol 20:11–35
Vacchi M, La Mesa M (1995) Diet of the Antarctic fish Trematomus newnesi Boulenger, 1902 (Nototheniidae) from Terra Nova Bay, Ross Sea. Antarct Sci 7:37–38
Vacchi M, Greco S, La Mesa M (1991) Ichthyological survey by fixed gears in Terra Nova Bay (Antarctica). Fish list and first results. Mem Biol Mar Oceanogr 19:197–202
Vacchi M, La Mesa M, Tarulli T (1992) Investigation by fixed gears on ichthyofauna of Terra Nova Bay (Ross Sea, Antarctica). In: Albertelli G, Ambrosetti W, Picazzo M, Ruffoni Riva T (eds) Atti IX Congr Ass Ital Oceanol Limnol. Lang, Genoa, pp 659–664
Vacchi M, La Mesa M, Castelli A (1994) Diet of two coastal nototheniid fish from Terra Nova Bay, Ross Sea. Antarct Sci 6:61–65
Vacchi M, Greco S, La Mesa M (1999) The coastal fish fauna of Terra Nova Bay, Ross Sea. In: Faranda F, Guglielmo L, Ianora A (eds) Ross Sea ecology. Italiantartide Expeditions (1987–1995). Springer, Berlin Heidelberg New York, pp 457–468
Vacchi M, Cattaneo-Vietti R, Chiantore M, Dalù M (2000) Predator-prey relationship between the nototheniid fish Trematomus bernacchii and the Antarctic scallop Adamussium colbecki at Terra Nova Bay (Ross Sea). Antarct Sci 12:64–68
Vacchi M, La Mesa M, Ozouf-Costaz C (2001) First occurrence of Acanthodraco dewitti Skóra, 1995 (Notothenioidei, Bathydraconidae) in the Ross Sea. Polar Biol 24:471–472
Van Heezik Y (1988) Diet of Adélie penguins during the incubation period at Cape Bird, Ross Island, Antarctica. Notornis 35:23–26
Veit RR, Braun BM (1984) Hydrographic fronts and marine bird distribution in Antarctic and subantarctic waters. Antarct J US 19:165–167
Voronina EP, Neelov AV (2001) Structural traits of alimentary tract of fishes of the family Channichthyidae (Notothenioidei). J Ichthyol 41:778–788
Voskoboinikova OS (1982) Reduction of the pterygoid bones of the visceral skeleton during evolution of the suborder Notothenioidei (Perciformes). J Ichthyol 22:105–111
Voskoboinikova OS (2001) Evolutionary significance of heterochronies in the development of the bony skeleton in fishes of the suborder Notothenioidei (Perciformes). J Ichthyol 41:415–424
Waite ER (1911) Antarctic fishes. British Antarctic Expedition 1907–1909, Reports on the Scientific Investigations. Biology 2:11–16
Williams R, Duhamel G (1994) Studies on fish of the Indian Ocean sector of the Southern Ocean during the BIOMASS Programme. In: El-Sayed SZ (ed) Southern Ocean ecology: the BIOMASS perspective. University Press, Cambridge, pp 211–229
Wohlschlag DE (1968) Fishes beneath Antarctic ice. Aust Nat Hist 16:45–48
Wöhrmann APA (1998) Aspects of eco-physiological adaptations in Antarctic fish. In: Prisco G di, Pisano E, Clarke A (eds) Fishes of Antarctica: a biological overview. Springer, Milan, pp 119–128
Wyanski DM, Targett TE (1981) Feeding biology of fishes in the endemic antarctic Harpagiferidae. Copeia 3:686–693
Young EC (1963) Feeding habits of the South Polar skua Catharacta maccormicki. Ibis 105:301–318
Yukhov VL (1971) The range of Dissostichus mawsoni Norman and some features of its biology. J Ichthyol 11:8–18
Zwally HJ, Comiso C, Gordon AL (1985) Antarctic offshore leads and polynyas and oceanographic effects. In: Jacobs SS (ed) Oceanology of the Antarctic continental shelf. Antarct Res Ser 43. American Geophysical Union, Washington DC, pp 203–226
Acknowledgements
This study was financially supported by the PNRA (Italian National Antarctic Research Program). J.T.E. was supported by National Science Foundation grant OPP 94-16870 and an Ohio University Presidential Research Scholar Award.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
La Mesa, M., Eastman, J.T. & Vacchi, M. The role of notothenioid fish in the food web of the Ross Sea shelf waters: a review. Polar Biol 27, 321–338 (2004). https://doi.org/10.1007/s00300-004-0599-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00300-004-0599-z