
Abstract
In response to carbon and/or nitrogen limitation, diploid cells of Saccharomyces cerevisiae either sporulate or develop pseudohyphae. Although the signal transduction pathways leading to these developmental changes have been extensively studied, how nutritional signals are integrated is not clearly understood. Results of this study indicate that reducing glucose concentration from 2% (SLAD) to 0.05% (SLALD) causes an increase in the magnitude of filamentation as well as a discernible reduction in the time required for pseudohyphal development. Further, the pseudohyphal defect of gpa2, gpr1and gpa2gpr1 but not the mep2 mutant strain is overcome on SLALD. Low glucose also induced pseudohyphae in mep2gpr1 but not mep2gpa2 strain suggesting that GPR1 inhibits pseudohyphae by inhibiting GPA2 function. Accordingly, deleting GPA2 in mep2gpr1 mutant abrogated pseudohyphae formation in SLALD. Further, replenishment of glucose suppressed pseudohyphal differentiation in wild-type cells grown in SLAD medium. However, in SLALD, glucose replenishment suppressed the filamentation response of gpa2 mutants but not that of strains carrying the wild-type GPA2. Increased trehalose levels correlated with decreased pseudohyphae formation. Results of this study demonstrate that filamentation in response to nitrogen limitation occurs as glucose becomes limiting.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Fungi belonging to diverse groups exhibit dimorphic transition in response to environmental conditions. This transition from yeast to hyphal mode or vice-versa has been implicated in the pathogenecity of both plant and human fungal pathogens such as Cryptococcus neoformans, Candida albicans and Ustilago maydis (reviewed in Lengeler et al. 2000). Our current understanding of dimorphic transition in response to nutritional vagaries is mainly derived from studies conducted in Saccharomyces cerevisiae. On solid medium, upon carbon limitation haploid yeast cells become elongated, form filaments and invade agar, a process termed as invasive growth (Gimeno et al. 1992). Diploid yeast cells facing carbon limitation enter into the quiescent G0 phase (reviewed in Gray et al. 2004) and in response to limited nitrogen but abundant glucose they differentiate into pseudohyphae (Gimeno et al. 1992; reviewed in Lengeler et al. 2000). They sporulate when both carbon and nitrogen are limiting (reviewed in Engebrecht 2003; Honigberg and Purnapatre 2003). These and other diverse developmental modes such as mating in response to pheromones are due to an integration of independent biological processes (Palecek et al. 2002; Roberts and Fink 1994). For example, pseudohyphal differentiation is determined by cell elongation, change in budding pattern from bipolar to unipolar and expression of flocculin (Gimeno et al. 1992; Gancedo 2001). On the other hand, mating induced by pheromones, involves a change in cell morphology followed by cell fusion caused by flocculin expression, leading to diploidisation (Roberts and Fink 1994; Erdman and Snyder 2001). FLO11, which is regulated by diverse signals (Rupp et al. 1999), plays a critical role in both haploid invasive growth and diploid pseudohyphal differentiation (Lo and Dranginis 1998). SRB10, a cyclin dependent protein kinase of RNA PolII holoenzyme, represses a substantial fraction of genes including FLO11, when cells are grown in rich medium (Holstege et al. 1998). SNF1 alleviates glucose-mediated repression of FLO11 through NRG1 and NRG2 (Kuchin et al. 2002).
The expression of FLO11 is regulated by elements of both the cAMP-PKA pathway through FLO8 and the MAPK pathway through STE12/TEC1 (Rupp et al. 1999).
Pseudohyphal differentiation in S. cerevisiae is mainly regulated by the intracellular TOR1 (Cutler et al. 2001) and the SNF1 encoded kinases (Kuchin et al. 2002) and the cell surface receptors that receive extracellular signal from glucose represented by the G protein coupled receptor (GPCR) GPR1, the RAS signaling system (Lorenz et al. 2000) and the MEP2 encoded ammonium permease (Lorenz and Heitman 1998).
TOR proteins, members of the conserved phosphatidyl inositol 3-kinase superfamily, regulate cell growth and development in response to nutrient availability (Thomas and Hall 1997; Crespo and Hall 2002; Zurita-Martinez and Cardenas 2005). Inhibition of TOR function by rapamycin induces physiologic conditions reminiscent of carbon and nitrogen starvation (Cutler et al. 2001; Beck and Hall 1999). One of the known targets of TOR signaling pertinent to pseudohyphal differentiation is GLN3, a GATA type transcriptional activator whose activity is normally induced by nitrogen starvation. It has been shown that GLN3 is also regulated by SNF1 (Betram et al. 2002), an evolutionarily conserved protein kinase whose main role is to derepress the glucose-repressed genes. Interference with TOR1 (Cutler et al. 2001) or SNF1 (Kuchin et al. 2002) signaling results in defective pseudohyphal differentiation, but the molecular mechanism/s of these pathways is not clearly understood. Recent evidence indicates that SNF1 functions as a glucose deprivation sensor in inducing filamentation response on alternative carbon sources (Van de velde and Thevelein 2008).
The ammonium permease coded by MEP2, is subject to nitrogen catabolite repression (NCR) and has been implicated in transducing the signal in response to low ammonium levels. A strain defective in MEP2 is unable to form pseudohyphae (Lorenz and Heitman 1998). Pseudohyphal defect conferred by MEP2 deletion is suppressed by activating the cAMP-PKA pathway but not the MAP kinase pathway, suggesting that its site of action is upstream of PKA mediated processes (Lorenz and Heitman 1998). While it has been shown that MEP2 transcription is increased (Marini et al. 1997), the protein levels did not reflect a corresponding change in response to ammonium levels (Lorenz and Heitman 1998). Based on the observation that activated GPA2 allele overcomes the pseudohyphal defect of a mep2 mutant strain it has been suggested that MEP2 acts upstream of GPA2 (Lorenz and Heitman 1998). There is evidence to indicate that the ammonium re-supplementation induced activation of PKA is mediated by MEP2 in a cAMP independent manner and that this activation is independent of the intracellular nitrogen status (Van Nuland et al. 2006). Extensive mutational analysis of MEP2 has indicated that the above function is independent of its ability to induce pseudohyphae upon nitrogen limitation. Recent studies suggest that MEP2 plays a role in pseudohyphal differentiation through the re-uptake of secreted ammonium under nitrogen starvation conditions (Boeckstaens et al. 2007). Interestingly, of the three paralogues of ammonium permeases, only mep2 mutant exhibits enhanced invasive growth. It is clear from the forgoing that the exact mechanism of MEP2 signaling in pseudohyphae formation has remained an enigma.
The cell surface signaling systems impinge mainly on the cAMP/PKA pathway. Ras1/2, partially redundant small GTP binding proteins, not only maintain the basal cAMP levels to activate the cAMP/PKA pathway but also activate the MAP kinase pathway to cause pseudohyphal differentiation (Mosch et al. 1996; Kubler et al. 1997). A defect in pseudohyphae formation due to a mutation in RAS2 can be suppressed either by activating the MAP kinase pathway or by extraneous addition of cAMP. GPR1, the low affinity cell surface receptor of glucose (Thevelein et al. 2005), coupled to GPA2, the α subunit of the heterotrimeric G protein, induces cAMP synthesis in response to glucose (Lorenz and Heitman 1997; Lorenz et al. 2000). Mutations in GPR1 or GPA2 inhibit pseudohyphae formation (Lorenz and Heitman 1997; Kubler et al. 1997; Lorenz et al. 2000). Recently, GPA2 has been shown to regulate PKA function independent of adenylate cyclase pathway based on the evidence that the inhibition of PKA by KRH1/2 (Kelch repeat proteins) is overcome by the GTP bound form of GPA2 (Lu and Hirsch 2005; Peeters et al. 2006). Accordingly, mutations in KRH1/2 which negatively regulate PKA in a cAMP independent manner enhance filamentation (Harashima and Heitman 2002). The biological significance of PKA regulation mediated by GPA2 through a cAMP independent pathway is not clearly understood. It has been suggested that the Kelch repeat proteins might serve as integrators of different signals that converge on PKA.
While it is evident that a large number of players participate in the complex process of pseudohyphal differentiation, how various signals are integrated and eventually translated into a unique biological output has largely remained elusive. For example, the physiological significance of upregulation of GPR1, a low affinity glucose receptor in response to nitrogen limitation, (Xue et al. 1998) is not understood. Although FLO11, a key member of the pseudohyphal differentiation pathway is glucose repressed (Kuchin et al. 2002), it has been tacitly assumed that pseudohyphal differentiation is a response to limited nitrogen in the presence of abundant glucose (reviewed in Lengeler et al. 2000). Based on the above and the observation that cAMP/PKA pathway also modulates SNF1 function (Hedbacker et al. 2004), we reasoned that alleviation of glucose repression may be an integral part of signaling for pseudohyphal development. We tested this possibility using low glucose and a variety of other experimental conditions. Our results clearly suggest that filamentation is a co-ordinated response to low glucose and low nitrogen. The implications of these results are discussed in the context of the signal transduction pathway as well as the carbon-nitrogen metabolic status of the cell.
Materials and methods
Yeast strains and media
Yeast strains used in this study are isogenic derivatives of the Σ1278b strain (Table 1). Standard genetic techniques were used for strain construction (Adams et al. 1997). The strains RIY 103, RIY 104 and RIY 105 were obtained by crossing singly or doubly disrupted strains and screening the isolated tetrads for a double or triple disruptant segregant followed by diploidization using HO plasmid induced mating type switching.
Synthetic low ammonia dextrose (SLAD) medium for scoring pseudohyphal growth was prepared as described (Gimeno et al. 1992; Lorenz and Heitman 1997). SLAD was modified to synthetic low ammonia low dextrose (SLALD) medium by decreasing the glucose concentration from 2 to 0.05%. Other modifications in SLAD are as mentioned in the figure legends. Strains were scored for pseudohyphal growth on SLAD, SLALD and SLAD with other modifications after incubation at 30°C for 6 days unless otherwise mentioned. Rapamycin was purchased from Calbiochem (Cat # 553210) and the stock solution was prepared as described (Cutler et al. 2001).
For glucose replenishment experiments, the cells were spread on one half of the plate with SLAD or SLALD medium. After incubation for 48 h, a trough measuring 5 × 3 cm was cut in the other half of the plate at a distance of 5 mm from the colonies along the diameter of the plate. A saturated solution of glucose (0.5 ml) was then added into the trough. This would result in an overall concentration of approximately 2% glucose in the plate after diffusion. As a control on a duplicate plate, sterile double distilled water was added into the trough. After this, the plates were incubated for 4 days before capturing images of the colonies.
Image analysis
Yeast colonies were photographed at 10× magnification using either a Leica Image Analysis System or a Nikon coolpix 8400 camera attached to a Nikon TS 100 microscope. The experiments were repeated several times and the images of representative colonies are shown. In case of differences in pseudohyphae formation in response to a change in the environmental condition, colonies exhibiting the least difference are shown in order to minimize variability. Additional images are provided as supplementary material as indicated in the legend.
β-galactosidase assay
β-galactosidase activity was measured using a FLO11::LacZ reporter plasmid (Rupp et al. 1999). Assays were done in triplicates and repeated at least five times.
Trehalose estimation
Cells were pre-grown in Synthetic complete (SC) medium for 20 h, washed twice with either 2% glucose or 0.05% glucose solution, diluted 1:5 in either SLAD or SLALD medium, respectively, and grown for 4–6 h. Trehalose content was estimated as described previously (Kienle et al. 1993). Trehalose content of cells grown to exponential phase in SC medium was estimated as a control and was found to be negligible. Assays were done in triplicates and repeated thrice.
Statistical analysis
The student’s t test was used to determine the statistical significance of the results of the above assays.
Results
Low glucose promotes pseudohyphal differentiation
To determine the role of glucose in pseudohyphal differentiation, we compared the filamentation of wild-type cells in SLAD and SLALD media. SLALD is modified SLAD with 0.05% glucose, a concentration at which glucose mediated repression is absent (Adams 1972; Carlson et al. 1984). We observed that wild-type cells developed pseudohyphae in SLALD after 2 days as compared to 4–6 days in SLAD (Fig. 1, panels a, b). The extent of pseudohyphae formed by wild-type cells at the end of 6 days was significantly enhanced in SLALD as compared to SLAD (Fig. 1, panels a, b). We also observed a gradual decrease in the filamentation response as the glucose concentration was increased gradually from 0.05 to 2% (Fig. 1, panel c).

Reducing glucose concentration results in early and enhanced filamentation. Panel a Pseudohyphae are observed either at the end of 2 or 6 days as indicated the figure. Panel b A time course showing the development of pseudohyphae of the same colony from day 3 to day 7 as indicated. Three independent colonies are shown. Panel c Pseudohyphae observed at the end of 6 days on varying glucose concentrations in Synthetic low ammonium (SLA). Glucose concentrations are indicated in percentages (0.05% corresponds to SLALD while 2% corresponds to SLAD). Three independent colonies are shown
Since the GPR1 mediated pathway has been shown to respond to abundant glucose, we determined the role of this pathway in filamentation in low glucose. We observed that the pseudohyphal differentiation defect of gpa2, gpr1 and gpa2gpr1 mutants was suppressed in SLALD (Fig. 2, compare panels a, b). These preliminary results suggest that low glucose favors pseudohyphal differentiation.

Effect of glucose limitation on mutant strains. Refer to figure S1 of supplementary material for additional colony images
A prediction of the above result was that the replenishment of glucose should delay or suppress filamentation. To test this idea, glucose was replenished after growing cells on SLAD and SLALD for 2 days. The colonies were observed after a further incubation of 4 days (see Sect. “Materials and methods” for details). As expected, wild-type cells did not develop pseudohyphae at the end of 4 days of incubation after glucose replenishment in SLAD medium (Fig. 3, panel a). On the other hand, colonies on the control plate that did not receive glucose developed normal pseudohyphae (Fig. 3, panel b). To determine the role of GPA2 and GPR1 in this process, we carried out the glucose replenishment experiments in strains bearing the corresponding mutations in SLALD (these mutants do not put forth pseudohyphae in SLAD medium). Replenishment of glucose did not suppress pseudohyphal differentiation in the wild-type and gpr1 mutant. However, glucose replenishment resulted in decreased pseudohyphae formation in gpa2 and gpa2gpr1 mutants (Fig. 4, compare panels a, b). This means that, once a strain with wild-type GPA2 forms pseudohyphae, it becomes refractory to inhibition by glucose. However, GPR1 did not seem to play a role in this process.

Effect of glucose replenishment on wild-type strain in SLAD. Images of colonies after addition of a saturated solution of glucose (a) or no glucose addition (b). Refer to figure S2 of supplementary material for additional colony images

Effect of glucose replenishment on the mutant strains in SLALD. Images of colonies after addition of a saturated solution of glucose (a) or no glucose (b). Refer to figure S3 of supplementary material for additional colony images
If low glucose is indeed a physiological signal, MEP2, a gene involved in ammonium signaling is not expected to participate in this process. As expected, the mep2 mutant did not show filamentation on SLALD. Surprisingly, GPR1 disruption overcame the filamentation defect of mep2 mutant in SLALD but not in SLAD (Fig. 2). During the course of this study, it was reported that mep2gpr1 mutant did not putforth pseudohyphae at the end of 24 h of growth in SLAD with 2 or 0.2% glucose (Van de velde and Thevelein 2008). We also did not observe pseudohyphae in mep2gpr1 mutant in SLALD at the end of 24 h (data not shown). This is not unexpected as the wild-type strain showed filamentation only after 48 h in SLALD (Fig. 1, panel b).
Why does GPR1 disruption overcome the inability of mep2 mutant to put forth pseudohyphae in SLALD but not SLAD medium? We invoke the following idea to explain the above epistatic relationship. Since GPR1 is a low affinity glucose receptor, it is possible that it exists in the unliganded form in low glucose, and that this unliganded form inhibits GPA2. Accordingly, inability of the mep2 mutant to put forth pseudohyphae under these conditions may be overcome by removing the inhibitory effect on GPA2 as a result of deletion of GPR1. This idea is supported by our observation that GPR1 deletion consistently resulted in enhanced filamentation in SLALD as compared to the wild-type strain (see Fig. S1 and S3 of supplementary material), suggesting a probable negative regulation of GPA2 by GPR1 under conditions of low glucose. The above explanation predicts that GPA2 is essential for the epistatic relationship between gpr1 and mep2 mutations in SLALD. If so, gpa2 mutation should abrogate the ability of mep2 gpr1 mutant to putforth pseudohyphae in SLALD. As expected, the mep2gpa2gpr1 triple mutant does not put forth pseudohyphae on SLALD (Fig. 2).
Analysis of Flo11p expression
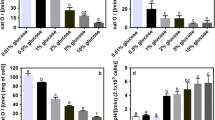
We monitored FLO11 expression by determining the β-galactosidase activity in cells transformed with a FLO11::LacZ reporter plasmid in SLAD as well as SLALD medium (Table 2). The data correlated well with pseudohyphae formation. FLO11 expression was substantially elevated in the gpr1 mutant strain grown in SLALD as compared to SLAD. This is consistent with the profuse filamentation exhibited by the gpr1 mutant strain in SLALD medium (see Fig. S1 and S3 of supplementary material). This data is also consistent with the idea that GPR1 may inhibit GPA2 under conditions of low glucose.
Lower trehalose content correlates with pseudohyphal differentiation
Increased trehalose content in response to stress is well documented (reviewed by Gasch 2003). Our reasoning that cells choose between a filamentation response and sporulation depending on the carbon/nitrogen signals, led us to determine trehalose levels in SLAD as well as SLALD media (Table 3). It is clear from our data that wild-type cells grown in SLAD have a lower trehalose level as compared to strains with gpr1 and gpa2 mutations, either alone or in combination. Therefore, it is possible that mutants unable to put forth pseudohyphae experience a metabolic stress. We consistently observed that the trehalose content in SLALD, a condition that favors pseudohyphae formation, is less than that in SLAD. This suggests that lowering glucose under conditions of low ammonium may favor filamentation and not stress response even in cells defective in GPR1 and GPA2. In contrast to the above results, although trehalose levels in a mep2 mutant were low in SLALD, pseudohyphae were not observed. Recent mutational analysis has shown that the MEP2 affects trehalase levels and pseudohyphal development differentially (van Nuland et al. 2006). Thus, our results indicate that alleviation of stress response may be necessary but not sufficient for filamentation.
It is to be noted that in the above experiments, trehalose content was measured after 6 h of exposure to SLAD and SLALD, but filamentation is a relatively long-term response. Nevertheless, lower trehalose content correlated well with pseudohyphal growth. It is conceivable that stress in SLAD could be due to a severe imbalance between the nitrogen and carbon availability. It is likely that as this imbalance is reduced, by reduction in glucose concentration, cells experience less stressful conditions. This is consistent with the glucose replenishment experiments in that re-exposure of cells to glucose prevented pseudohyphal differentiation.
Rapamycin does not prevent filamentation at low concentrations
Treatment of cells with rapamycin induces the transcription of nitrogen repressible genes and accordingly it is expected to enhance pseudohyphae formation. However, Cutler et al. (2001) observed that at the concentration tested, rapamycin inhibited pseudohyphal differentiation. This inhibition could not be reversed by changing the ammonium concentration (Cutler et al. 2001). They explained this paradox, by suggesting that rapamycin inhibits TOR above a threshold resulting in constitutive expression of NCR genes which in turn inhibits pseudohyphae formation. The above idea predicts that exposing the cells to concentrations of rapamycin lower than that used by Cutler et al. should result in filamentation. Accordingly, rapamycin concentration was varied from 10 to 0.01 ng/ml in SLAD (Fig. 5, panel a), modified SLAD with 5 μM ammonium (Fig. 5, panel b) and SLALD (Fig. 5, panel c). Rapamycin at 10 ng/ml inhibited pseudohyphae formation consistent with the previous report (Cutler et al. 2001). However, we observed that rapamycin concentration at or below 0.1 ng/ml did not prevent pseudohyphal development in the wild-type strain in SLAD medium, a result consistent with our prediction. The above effect was more pronounced when the concentration of ammonium was reduced to 5 μM. In fact, we observed hyper-filamentation at a concentration of 0.01 ng/ml of rapamycin at 5 μM ammonium concentration. We observed that reducing glucose concentration from 2 to 0.05% overcomes the inhibition of filamentation imposed by rapamycin on the wild-type strain at 1ng/ml in SLAD (Fig. 5, compare panels a, c). Also, rapamycin did not prevent pseudohyphae formation in the gpr1 mutant at or below 1 ng/ml and in the gpa2 mutant at or below 0.1 ng/ml in SLALD medium (Fig. 6). The mep2 strain, however, did not respond to changes in rapamycin concentrations under the conditions tested (data not shown). These results point out to the possibility that above response is mediated by MEP2 in a GPA2 independent manner.

Effect of changing rapamycin concentrations on the wild-type strain. Images of colonies on SLAD (a), modified SLAD with 5μM NH4 + (b) and SLALD (c), containing different rapamycin concentrations (ng/ml)

Effect of changing rapamycin concentrations on the mutant strains. Images of colonies on SLALD containing different rapamycin concentrations (ng/ml) as indicated
Discussion
Glucose signaling
There is extensive body of evidence indicating that S. cerevisiae has optimized rapid growth in yeast form by fermenting glucose, an inefficient process in terms of energy production (Piskur et al. 2006). Glucose mediated repression is one of the key elements of this optimization. It is generally believed that abundant glucose in conjunction with limited nitrogen is the trigger for pseudohyphal differentiation (Lengeler et al. 2000). However, expression of FLO11, one of the targets of the pseudohyphal signaling is glucose repressed (Kuchin et al. 2002). This apparent paradox has not been clearly understood. Based on our results, we suggest that the wild-type cell grows initially in the yeast form on SLAD and as alleviation of glucose repression occurs due to a decrease in glucose levels, cells switch over to pseudohyphal mode of growth, provided the cells experience nitrogen depletion.
What is the threshold concentration of glucose below which pseudohyphal differentiation is favored? Yeast cells growing on 2% glucose in liquid culture start utilizing ethanol when glucose concentration drops to 0.5%, due to the alleviation of glucose repression (Piskur et al. 2006). It is possible that this occurs on solid medium as well. It is to be noted that unlike liquid medium, ethanol produced near the colony may prevent the availability of glucose to the cells resulting in the alleviation of glucose repression at a concentration above 0.5%. In fact, cells unable to utilize ethanol due to mitochondrial dysfunction remain very small, suggesting that ethanol utilization starts quite early during growth on solid media with glucose (Chen and Clark-Walker 1999).
That the alleviation of glucose repression is an important signal for pseudohyphal differentiation is also supported by the recent observation that filamentation is enhanced in microcolonies of hxk2 mutants as compared to wild-type cells in SLAD (Van de velde and Thevelein 2008). In contrast, in sucrose, a medium that does not confer glucose repression, hxk2 mutants as well as wild-type cells show enhanced filamentation (Van de velde and Thevelein 2008). The above observation that the effect of hxk2 mutation manifests only under conditions of glucose repression reinforces the idea that pseudohypal differentiation is controlled by glucose repression. Based on the above and the results presented in this communication, we suggest that pseudohyphal differentiation observed on SLAD occurs after glucose concentration drops to a level sufficient to alleviate glucose repression.
We suggest that in a wild-type strain, pseudohyphal differentiation occurs primarily due to nitrogen limitation in conjunction with a decrease in the concentration of glucose. We propose (Fig. 7) that, the primary event is the concommitant activation of SNF1 mediated by low glucose levels (Carlson 1999) as well as the cAMP-PKA pathway (Hedbacker et al. 2004). Activation of FLO11 also occurs by PKA mediated inhibition of SFLI (a FLO11 repressor) as well as activation of FLO8 (Pan and Heitman 2002). This reciprocal regulation of FLO11 appears to be critically dependent on the relative levels of glucose and ammonium and is expected to play a key role in regulating filamentation. Thus, the metabolic transition coupled to the signal transduction process may lead to the physiological and morphological events associated with pseudohyphal differentiation.

Schematic illustration of the cross talk between metabolic and morphological events during pseudohyphal differentiation in response to low glucose and ammonia. In presence of low nitrogen MEP2 is expressed due to the alleviation of NCR mediated by the activation of SNF1 and TOR. Also, GPR1 is upregulated in low nitrogen. A concomitant reduction in glucose levels results in FLO11 expression due to the relief of inhibition from SRB10, NRG1,2 and SFL1 in conjunction with the activation mediated by FLO8. According to our model, GPR1 probably exists in the unliganded form in low glucose, which inhibits GPA2. Also, MEP2 may activate PKA independent of GPA2. The coupling of the extracellular nutrient depletion to the intracellular metabolic state ensures that pseudohyphal differentiation does not occur fortuitously but occurs only when all the necessary conditions are met. Evolution of these “co-incidence circuits” increases the signal to noise ratio.  , activation;
, activation;  , inhibition;
, inhibition;  , extracellular signaling;
, extracellular signaling;  , novel interactions proposed based on our data,
, novel interactions proposed based on our data,  , GLN3 mediated signaling
, GLN3 mediated signaling
SNF1 appears to play a critical role in filamentation in addition to alleviating glucose repression. For example, it has been observed that disruption of SNF4 abolishes pseudohyphae not only in SLAD but also in low ammonium sucrose medium, where glucose repression is absent (Van de velde and Thevelein 2008). The observations that SNF1 is involved in nitrogen signaling (Betram et al. 2002; Orlova et al. 2006) and that SNF1 disruption abrogates filamentation (Kuchin et al. 2002) are consistent with the above.
A key observation of our study is that the disruption of GPR1 overcomes the pseudohyphal differentiation defect of a mep2 mutant in SLALD but not in SLAD. This raises the possibility that GPR1 inhibits GPA2 as glucose levels fall. In this context, GPR1 may be a “true” low affinity glucose receptor and its function may be to ensure that the cells grow in yeast form as long as sufficient glucose is available. It is likely that induction of GPR1 in response to nitrogen depletion (Xue et al. 1998) is to allow the switch over to pseudohyphal mode of growth only after glucose also becomes limiting.
We suggest that like budding, filamentation is vegetative growth but with a metabolic as well as morphological difference. We observed that lower trehalose levels correlated well with pseudohyphal mode of growth, suggesting that the cells have opted to enter into filamentation and not sporulation. It has been demonstrated that meiosis and filamentation are two mutually exclusive developmental processes regulated by the availability of carbon and nitrogen (Donzeau and Bandlow 1999). Accordingly, although filamentation occurs in SLALD as glucose and ammonium deplete, the cells may not sporulate as sporulation is a specific response to complete starvation in the presence of acetate.
TOR signaling
The involvement of TOR in pseudohyphal regulation is much more difficult to evaluate as TOR has been shown to regulate many downstream events in response to both carbon and nitrogen limitations (Thomas and Hall 1997; Crespo and Hall 2002; Cutler et al. 2001; Beck and Hall 1999). Based on our observation that rapamycin can induce pseudohyphal differentiation at low concentrations, it is likely that varying the concentration of rapamycin could modulate TOR, leading to different biological consequences. Alternatively, TOR could exist in functional states having different susceptibilities to rapamycin, depending upon the constellation of nutritional signals. That the effect of rapamycin is at the metabolic level is supported by the observation that the loss of function of 396 genes that are involved in cell growth and metabolism alter the cellular response to rapamycin (Xie et al. 2005).
The possibility that the degree or extent of activation or repression of a protein kinase such as TOR1 can cause opposite biological effects opens new avenues in the design and interpretation of experiments. Regardless of the above, our observation that rapamycin promotes cellular differentiation under specific environmental conditions should allow manipulation of signaling pathways for inducing cellular proliferation or differentiation.
Our observations have far reaching consequences in the nutrient dependent regulation of cellular differentiation. The evolutionary conservation of regulation mediated by TOR1, SNF1 and PKA protein kinases in response to carbon/nitrogen availability underscores the significance of our observation. Given an intricate connection between metabolic and genetic regulation in pseudohyphal differentiation, it is not surprising that the filamentation response is susceptible to minute changes in the environmental stimulus, making it difficult to interpret experimental results. As demonstrated here, phenomenological evidence coupled with existing genetic and biochemical data may be necessary to provide a better understanding of pseudohyphal differentiation. Clearly, the response of the genetic regulatory network to physiological status governed by glucose and nitrogen availability is the key determinant of pseudohyphal differentiation.
References
Adams BG (1972) Induction of galactokinase in Saccharomyces cerevisiae: kinetics of induction and glucose effects. J Bacteriol 111:308–315
Adams A, Gottschling DE, Kaiser CA, Stearns T (1997) Methods in yeast genetics. Cold Spring Harbor Laboratory, New York, pp 1–157
Beck T, Hall MN (1999) The TOR signaling pathway controls nuclear localization of nutrient-regulated transcription factors. Nature 402:689–692
Betram PG, Choi JH, Carvalho J, Chan T, Ai W, Steven Zheng XF (2002) Convergence of TOR-nitrogen and Snf1-glucose signaling pathways onto Gln3. Mol Cell Biol 22:1246–1252
Boeckstaens M, Andre B, Marini AM (2007) The yeast ammonium transport protein Mep2 and its positive regulator, the Npr1 kinase, play an important role in normal and pseudohyphal growth on various nitrogen media through retrieval of excreted ammonium. Mol Microbiol 64:534–546
Carlson M (1999) Glucose repression in yeast. Curr Opin Microbiol 2:202–207
Carlson M, Osmond BC, Neigeborn L, Botstein D (1984) A suppressor of SNF1 mutations causes constitutive high-level invertase synthesis yeast. Genetics 107:19–32
Chen XJ, Clark-Walker GD (1999) The petite mutation in yeasts: 50 years on. Int Rev Cytol 194:197–237
Crespo JL, Hall MN (2002) Elucidating TOR signaling and rapamycin action: lessons from Saccharomyces cerevisiae. Microbiol Mol Bio Rev 66:579–591
Cutler NS, Pan X, Heitman J, Cardenas M (2001) The TOR signal transduction cascade controls cellular differentiation in response to nutrients. Mol Biol Cell 12:4103–4113
Donzeau M, Bandlow W (1999) The yeast trimeric guanine nucleotide-binding protein α subunit, Gpa2p, controls the meiosis-specific kinase Ime2p activity in response to nutrients. Mol Cell Biol 19:6110–6119
Engebrecht J (2003) Cell signaling in yeast sporulation. BBRC 306:325–328
Erdman S, Snyder M (2001) A filamentous growth response mediated by the yeast mating pathway. Genetics 159:919–928
Gancedo JM (2001) Control of pseudohyphae formation in Saccharomyces cerevisiae. FEMS Microbiol Rev 25:107–123
Gasch AP (2003) The environmental stress response: a common yeast response to diverse environmental stress. In: Hohmann S, Mager WH (eds) Yeast stress response. Springer-Verlag, Berlin, pp 11–57
Gimeno CJ, Ljungdahl PO, Styles CA, Fink GR (1992) Unipolar cell divisions in the yeast S. cerevisiae lead to filamentous growth: regulation by starvation and RAS. Cell 68:1077–1090
Gray JV, Petsko GA, Johnston GC, Ringe D, Singer RA, Werner-Wahsburne M (2004) “Sleeping beauty”: quiescence in Saccharomyces cerevisiae. Microbiol Mol Biol Rev 68:187–206
Harashima T, Heitman J (2002) The Gα protein Gpa2 controls yeast differentiation by interacting with kelch repeat proteins that mimic Gβ subunits. Mol Cell 10:163–173
Hedbacker K, Townley R, Carlson M (2004) Cyclic AMP-dependent protein kinase regulates the subcellular localization of Snf1-Sip1 protien kinase. Mol Cell Biol 24:1836–1843
Holstege FC, Jennings EG, Wyrick JJ, Lee TI, Hengartner CJ, Green MR, Golub TR, Lander ES, Young RA (1998) Dissecting the regulatory circuitry of a eukaryotic genome. Cell 95:717–728
Honigberg SM, Purnapatre K (2003) Signal pathway integration in the switch from the mitotic cell cycle to meiosis in yeast. J Cell Sci 116:2137–2147
Kienle I, Burgert M, Holzer H (1993) Assay of trehalose with acid trehalase purified from Saccharomyces cerevisiae. Yeast 9:607–611
Kubler E, Mosch H, Rupp S, Lisanti MP (1997) Gpa2p, a G-protein α-subunit, regulates growth and pseudohyphal development in Saccharomyces cerevsiae via a cAMP-dependent mechanism. J Biol Chem 272:20321–20323
Kuchin S, Vyas VK, Carlson M (2002) Snf1 protein kinase and the repressors Nrg1 and Nrg2 regulate FLO11, haploid invasive growth and diploid pseudohyphal growth. Mol Cell Biol 22:3994–4000
Lengeler KB, Davidson RC, D’Souza C, Harashima T, Shen W, Wang P, Pan X, Waugh M, Heitman J (2000) Signal transduction cascades regulating fungal development and virulence. Microbiol Mol Biol Rev 64:746–785
Lo W, Dranginis AM (1998) The cell surface flocculin Flo11 is required for pseudohyphae formation and invasion by Saccharomyces cerevisiae. Mol Biol Cell 9:161–171
Lorenz MC, Heitman J (1997) Yeast pseudohyphal growth is regulated by GPA2, a G protein α homolog. EMBO J 16:7008–7018
Lorenz MC, Heitman J (1998) The MEP2 ammonium permease regulates pseudohyphal differentiation Saccharomyces cerevisiae. EMBO J 17:1236–1247
Lorenz MC, Pan X, Harashima T, Cardenas ME, Xue Y, Hirsch JP, Heitman J (2000) The G protein-coupled receptor Gpr1 is a nutrient sensor that regulates pseudohyphal differentiation in Saccharomyces cerevisiae. Genetics 154:609–622
Lu A, Hirsch JP (2005) Cyclic AMP-independent regulation of protein kinase: a substrate phosphorylation by Kelch repeat homologues. Eukaryot Cell 4:1794–1800
Marini A, Soussi-Boudekou S, Vissers S, Andre B (1997) A family of ammonium transporters in Saccharomyces cerevisiae. Mol Cell Biol 17:4282–4283
Mosch H, Roberts RL, Fink G (1996) Ras2 signals via the Cdc42/Ste20/mitogen-activated protein kinase module to induce filamenatous growth in Saccharomyces cerevisiae. Proc Natl Acad Sci USA 93:5352–5356
Orlova M, Kanter E, Krakovich D, Kuchin S (2006) Nitrogen availability and TOR regulate the Snf1 protein kinase in Saccharomyces cerevisiae. Eukaryot Cell 5:1831–1837
Palecek SP, Parikh AS, Kron SJ (2002) Sensing, signaling and integrating physical processes during Saccharomyces cerevisiae invasive and filamentous growth. Microbiol 148:893–907
Pan X, Heitman J (2002) Protein kinase A operates a molecular switch that governs yeast pseudohyphal differentiation. Mol Cell Biol 22:3981–3993
Peeters T, Louwet W, Gelade R, Nauwelaers D, Thevelein JM, Versele M (2006) Kelch-repeat proteins interacting with the G{alpha} protein Gpa2 bypass adenylate cyclase for direct regulation of protein kinase A in yeast. Proc Natl Acad Sci USA 103:13034–13039
Piskur J, Rozpedowska E, Polakava S, Merico A, Compagno A (2006) How did Saccharomyces cerevisiae become a good brewer? Trends Genet 22:183–187
Roberts RL, Fink GR (1994) Elements of a single MAP kinase cascade in Saccharomyces cerevisiae mediate two developmental programs in the same cell type: mating and invasive growth. Genes Dev 8:2974–2985
Rupp S, Summers E, Lo HJ, Madhani H, Fink G (1999) MAP kinase and cAMP filamentation signaling pathways converge on the unusually large promoter of the yeast FLO11 gene. EMBO J 18:1257–1269
Thevelein JM, Gelade R, Holsbeeks I, Legatie O, Popova Y, Rolland F, Stolz F, Van de Velde S, Van Dijck P et al (2005) Nutrient sensing systems for rapid activation of the protein kinase A pathway in yeast. Biochem Soc Trans 33:253–256
Thomas G, Hall MN (1997) TOR signaling and control of cell growth. Curr Opin Cell Biol 9:782–787
Van de Velde S, Thevelein JM (2008) cAMP-PKA and Snf1 signaling mechanisms underlie the superior potency of sucrose for induction of filamentation in yeast. Eukaryot Cell 7:286–293
van Nuland A, Vandormael P, Donaton M, Alenquer M, Lourenco A, Quintino E, Versele M, Thevelein JM (2006) Ammonium permease-based sensing mechanism for rapid ammonium activation of the protein kinase: a pathway in yeast. Mol Microbiol 59:1485–1505
Xie MW, Jin F, Hwang H, Hwang S, Anand V, Duncan MC, Huang J (2005) Insights into TOR function and rapamycin response: chemical genomic profiling by using a high-density cell array method. Proc Natl Acad Sci USA 102:7215–7220
Xue Y, Batlle M, Hirsch JP (1998) GPR1 encodes a putative G protein-coupled receptor that associates with the Gpa2 Gα subunit and functions in a Ras-independent pathway. EMBO J 17:1996–2007
Zurita-Martinez SA, Cardenas ME (2005) Tor and cyclic AMP-Protein kinase A: two parallel pathways regulating expression of genes required for cell growth. Eukaryot Cell 4:63–71
Acknowledgments
This work was supported by financial assistance provided by Department of Science and Technology (India) under the WOS-‘A’ scheme awarded to Dr. Revathi Iyer. We thank Prof. J. Heitman for graciously providing the strains. We thank the Sophisticated Analytical Instruments Facility (SAIF), IIT Bombay, for providing the imaging facility.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by G. Braus.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Iyer, R.S., Das, M. & Bhat, P.J. Pseudohyphal differentiation defect due to mutations in GPCR and ammonium signaling is suppressed by low glucose concentration: a possible integrated role for carbon and nitrogen limitation. Curr Genet 54, 71–81 (2008). https://doi.org/10.1007/s00294-008-0202-1
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00294-008-0202-1