
Abstract
Salinity is one of the most important factors that limit the productivity of agricultural soils. Certain plant growth-promoting bacteria (PGPB) have the ability to stimulate the growth of crop plants even under salt stress. In the present study, we analysed the potential of PGPB Bacillus toyonensis COPE52 to improve the growth of tomato plants and its capacity to modify its membrane lipid and fatty acid composition under salt stress. Thus, strain COPE52 increased the relative amount of branched chain fatty acids (15:0i and 16:1∆9) and accumulation of an unknown membrane lipid, while phosphatidylethanolamine (PE) levels decreased during growth with 100 and 200 mM NaCl. Importantly, direct and indirect plant growth-promoting (PGP) mechanisms of B. toyonensis COPE52, such as indole-3-acetic acid (IAA), protease activity, biofilm formation, and antifungal activity against Botrytis cinerea, remained unchanged in the presence of NaCl in vitro, compared to controls without salt. In a greenhouse experiment, tomato plants (Lycopersicon esculentum ‘Saladette’) showed increased shoot and root length, higher dry biomass, and chlorophyll content when inoculated with B. toyonensis COPE52 at 0 and 100 mM NaCl. In summary, these results indicate that Bacillus toyonensis COPE52 can modify cell membrane lipid components as a potential protecting mechanism to maintain PGP traits under saline-soil conditions.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Soil salinity is one of the major abiotic stresses that adversely affect agricultural practices, constituting a major global problem and affecting almost 1 billion ha worldwide [1, 2]. For instance, salinity has caused a loss of approximately 65% in wheat yield in moderately saline soils [3], because it affects almost all developing aspects of plant biology including, germination, vegetative growth, and reproductive stages [4, 5]. Particularly, glycophyte plants like tomato plants (Lycopersicon esculentum) are forced to induce their tolerance mechanisms against salinity at salt concentrations as low as 10 mM NaCl (the salt tolerance grade also depends on the cultivar), to avoid adverse effects on growth and productivity [1, 6]. To counteract the negative effects of agricultural practices in saline soils, several strategies have been developed, including selection of genotypes resistant to salt stress, genetic engineering of hypertolerant saline plants, vegetative bioremediation, utilization of better irrigation management strategies, and the use of plant growth-promoting bacteria (PGPB) [7,8,9]. Various PGPB have been isolated and characterized by their ability to improve the growth of plants under salt stress conditions [10]. The PGPB contain their own saline stress tolerance mechanisms and in turn, can induce varying degrees of salt tolerance during interaction with plants through ion homeostasis, accumulation of osmolytes or by reducing ethylene and reactive oxygen species [11].
During growth in saline soils, PGPB and plants interact beneficially to survive. In some cases, PGPB exhibit synergistic effects of two plant growth mechanisms to counteract the salt stress in plants. For example, Orozco-Mosqueda and colleagues [12] demonstrated that 1-aminocyclopropane-1-carboxylic acid (ACC) deaminase and disaccharide trehalose production in Pseudomonas sp. UW4, act synergistically to counteract the damaging effects of salt stress and allow the growth of tomato plants.
Another mechanism of tolerance described in some PGPB is the modification of its membrane lipid components. For example, the bacterial strain L115 belonging to the genus Ochrobactrum, modifies the degree of unsaturation of fatty acids and increased its phosphatidylcholine levels under salt stress (and high temperature) growth conditions, allowing it to maintain growth-promoting effects of Arachis hypogaea plants [13]. It is important to mention that the survival of bacteria in response to environmental changes is often determined by their capacity to adapt by altering the composition of the lipid bilayer. There are two types of lipid modifications: One in which the lipids can be modified to obtain a membrane with different properties. For instance, Vences-Guzmán [14] described that the hydroxylation of ornithine lipids (OLs) is important for Rhizobium tropici to resist stress conditions such as acidic pH or high temperatures. In the second type, existing lipids are degraded and lipids with new characteristics are synthesized de novo replacing the old lipids [15]. Additional mechanisms to stabilize membrane fluidity in bacteria involve changes in fatty acid (FA) composition of membrane lipids such as the phospholipids (PL) [16].
Another genus of bacteria widely studied for being a promoter of plant growth under conditions of salt stress is Bacillus [17]. Bacillus amyloliquefaciens H-2-5 was able to enhance the growth of Chinese cabbage, radish, tomato, and mustard plants and additionally, strain H-2-5-mediated mitigation of short-term salt stress when tested on soybean plants that were affected by sodium chloride. H-2-5 did this by regulating gibberellin, abscisic acid, jasmonic acid, salicylic acid and increasing plant proline levels [18].
In the case of the Bacillus toyonensis, there are few studies describing it as a PGPB [19,20,21]. In fact, the bacterial endophyte strain COPE52 of B. toyonensis presents diverse mechanisms of growth promotion in blueberry plants, such as the production of indole acetic acid, protease activity, and the emission of volatile compounds like acetoin, 2,3-butanediol and dimethyl disulphide, that can act as potential plant growth-promoting mechanisms [21]. However, their beneficial activities have not been analysed under conditions of saline stress.
In this study, we analysed the lipid and fatty acid composition of the bacterium B. toyonensis COPE52 while in growth under salt stress, as well as its growth-promoting mechanisms during inoculation in tomato (Lycopersicon esculentum ‘Saladette’) plants.
Materials and Methods
Bacterial Strain and Growth Conditions
B. toyonensis COPE52 was previously isolated and characterized [21]. Strain COPE52 was grown at 30 °C for 24 h on Luria Bertani (LB), and routinely maintained at 4 °C. The fungical plant pathogen Botrytis cinerea and Fusarium oxysporum were inoculated and maintained on potato dextrose agar (PDA) at 30 °C for 4–5 days in darkness and maintained at 4 °C.
Analysis of Membrane Components
Growth of the bacteria for Lipid Extraction
Twenty-five ml cultures with 0, 100, or 200 mM NaCl were adjusted to an optical density of 0.1. A 1 ml aliquot was transferred to a sterile tube and 0.5 μCi of [1-14C] acetate (Amersham Biosciences). The cultures were incubated at 30 °C while shaking for 24 h. The cells of the larger cultures were harvested by centrifugation at 6000 rpm for 10 min at 4 °C. Pellets were washed with water and re-suspended in 100 μl water. Cells from the labelled cultures were centrifuged for 1 min at 14,000 rpm, washed once and resuspended in 100 μl water.
Lipid Extraction, Separation, and Analysis of Phospholipids
Lipids were extracted using a chloroform/methanol/water extraction [22]. 375 µl methanol:chloroform (2:1) were added to the suspended cells and the mixture was vortexed. Then, 125 µl of water and 125 µl chloroform were added to obtain a separation into lower and upper phase. The lower phase, containing the lipids, was transferred to a new tube and washed once with the water, dried under N2, and dissolved in a suitable volume of chloroform/methanol 1:1 (v/v). Aliquots of the lipid extracts were spotted on HPTLC silica gel 60 plates (Merck, Pool, UK). Lipids were separated by two-dimensional TLC using chloroform/methanol/water (140:60:10, v/v/v) as solvents for the first dimension and chloroform/methanol/glacial acetic acid (130:50:20, v/v/v) as solvents for the second dimension. Unlabelled membrane lipids were visualized by iodine staining and radioactive membrane lipids were visualized by exposition to autoradiography film (Kodak) and in a PhosphorImager screen (Amersham Biosciences), respectively. Individual lipids were quantified using ImageQuant software (Amersham Biosciences).
Analysis of Fatty Acids by GC
Fatty acyl methyl esters (FAME) were prepared from total lipid extracts with 10% Methanol-BF3 (Sigma) [23] using tridecanoic acid as an internal standard and analysed using a Perkin Elmer Clarus 600 gas chromatography system coupled to a Clarus 600T mass spectrometer. The gas chromatograph was equipped with a column Elite 5-MS de Perkin Elmer (length 30 m; inner diameter 0.2 mm; film thickness 0.32 mm). Gas chromatograph conditions were as follows: 250 °C injector temperature, 300 °C detector temperature and hydrogen as the carrier gas. The temperature was programmed at 140 °C for 6 min and then increased by 10 °C/min to 240 °C for 5 min and finally to 250 °C for 5 min. Fatty acids were identified by comparing retention times to commercial standards (Sigma Chemical Co., St. Louis, MO, USA) and by MS fragmentation in the EI mode with the electron energy set to 70 eV.
Determination of the Plant Growth-Promoting Traits Under Salt Stress
Siderophore Production
The production of siderophores was evaluated on Chrome Azurol agar (CAS) medium [17] amended with 0, 100, 200 mM of NaCl. All experiments were performed in triplicate.
Colorimetric Assay for the IAA Determination
The IAA (Indole-3-acetic acid) content was determined based on the method described by Patten and Glick [24] with some modifications. Briefly, 25 ml flasks were inoculated, supplemented with a graduated series of NaCl concentrations (0, 100, 200 mM NaCl) at 30 °C on a rotary shaker at 150 rpm. Cells were then collected by centrifugation at 10,000×g for 15 min and 2 ml of Salkowski reagent were added to the supernatant. The absorbance of the pink auxin complex was read at 540 nm in a UV–Vis Spectrophotometer (JENWAY 7305). The calibration plot was constructed using dilutions of a standard total indole (Fluka, Switzerland) solution and the uninoculated medium with the reagent as a control. Experiments were performed in triplicate.
Proteases Production
Proteolytic activity was determined using skimmed milk agar (pancreatic digest of casein 5 g, yeast extract 2.5 g, glucose 1 g, 7% skim milk solution 100 ml, agar 15 g dissolved in 1 l distilled water, with 0, 100 or 200 mM NaCl). After 2 days incubation at 30 °C, a clear zone around the cells indicated positive proteolytic activity [25].
Biofilm Production
Biofilm formation capacity in bacteria was analysed following the protocol by Wei and Zhang [26]. Briefly, testing strains were grown in LB medium, supplemented with or without salt (100 or 200 mM of NaCl) to an O.D. of 1 and then diluted (1:1000) with fresh LB broth. A 0.5 ml diluted culture was transferred to an Eppendorf tube. Bacteria were incubated without agitation for 24, 48, and 72 h at 30 °C and the biofilm was quantified at each time points. The biofilm was stained with 0.1% (w/v) crystal violet for 15 min at room temperature and then rinsed thoroughly with water to remove unattached cells and residual dye. Ethanol (95%) was used to solubilize the dye that had stained the biofilm cells. The absorbance of the solubilized dye (A570) was determined with a UV–Vis Spectrophotometer (JENWAY 7305). All experiments were carried out in triplicate.
In Vitro Evaluation of Fungal Antagonism
The evaluation of fungal antagonism was performed as previously reported on Petri dish bioassays [25]. Bacterial isolate COPE52 was co-inoculated, simultaneously with the pathogenic fungi on PDA agar plates, amended with 0, 100 or 200 mM NaCl. Pathogenic B. cinerea strain has been previously tested and analysed in a previous work [25], while F. oxysporum strain was isolated as a pathogenic fungus from an agricultural field and donated by the INIFAP-Mexico. The bacterium strain was streaked onto plates in a cross shape, and a mycelial plug of 4 mm was deposited in the centre of each of the quadrants formed. The plates were incubated in the dark at 30 °C (BOD incubator), and the mycelial growth diameter was measured at day 6. Antifungal effects of volatile compounds emitted by the Bacillus were evaluated in divided Petri plates as follows: A bacterial inoculum of each strain (1 × 106 CFU) was simultaneously deposited on one side of the Petri plate and in the other section a mycelial plug of B. cinerea and Fusarium oxysporum (4 mm) was inoculated. The plates were incubated in the dark at 30 °C, and mycelial growth diameter was measured at day 6. All experiments were carried out in triplicate. The percentage of growth inhibition was measured using the following formula: % of growth inhibition = [(Ac − Ab) /Ac] × 100, where Ac is the control mycelial area and Ab is the mycelial area with treatment.
Evaluation of Plant Growth Promotion by Bacillus toyonensis COPE52 in a Greenhouse Under Salt Stress Conditions
Greenhouse pot experiments with tomato plants (Lycopersicon esculentum ‘Saladette’) were performed with 0, 100, and 200 mM NaCl in sterile peat moss. Greenhouse experiments were carried out as previously reported [12]. Briefly, tomato seeds were germinated, and after one week, seedlings of the same size were selected and transplanted into pots (one plant was transplanted in each pot). The experimental design included the following treatments: 24 control plants (1 plant per pot) without NaCl and/or COPE52 and other 24 plants were inoculated with COPE52 strain under normal conditions (no-salt addition). Another set of plants were irrigated with 100 (24 plants) or 200 mM NaCl (24 plants) and without bacterial inoculants, while other two groups of 24 plants each, were inoculated with COPE52 strain and watered with a 100 or 200 mM NaCl. Throughout the experiment, the plants were irrigated every third day with deionized water or saline solution, while constantly controlling the salt concentration by measuring electrical conductivity (Field Scout. Mod. 2265FS). Bacterial inoculants, adjusted to 1 × 108 Colony Forming Units (CFU)/mL, were applied every week with exception of the control groups. After 5 weeks of plant growth, the effect of adding the bacterial inoculum on the root and shoot length, fresh weight, weight dry and chlorophyll concentration was evaluated. The chlorophyll concentration was measured in at least three leaves from each plant [12].
Statistical Analysis
The results were analysed using Statistica 8.0 software, and analysis of variance and Duncan's test for mean comparison was used for multiple comparisons (P < 0.05). The fungal antagonism experiments were statistically analysed by Student’s t-test (P < 0.05).The greenhouse test results were analysed using a factorial design, comprising two factors and three levels (no-salt, medium and high salt stress conditions, and the presence and absence of B. toyonensis), followed by Duncan’s multiple range test (P ≤ 0.05).
Results
Effect of Salinity on Phospholipid Metabolism and Fatty Acid Composition
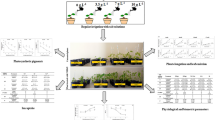
The cell viability of Bacillus toyonensis COPE52 was evaluated under salt concentrations of up to 200 mM NaCl. Here, no significant decrease in the growth of COPE52 was observed (Supplementary Table 1). Thin layer chromatography (TLC) analysis revealed that the presence of salt in the growth medium induced substantial changes in membrane lipid (ML) composition. The predominant membrane lipid was phosphatidylethanolamine (PE), followed in descending order by phosphatidylglycerol (PG), an unknown lipid (NI), and cardiolipin (CL) under non-saline conditions (Table 1 and Fig. 1). The unknown lipid NI migrated as phosphatidylcholine (PC), but it did not stain with Dragendorff reagent, so we exclude that it is PC (not shown). ML patterns for COPE52 were qualitatively similar for all experimental conditions, but quantitative changes were observed for individual ML. The addition of salt to the growing medium of COPE52 caused an increase in NI from 11 to 21% with 100 mM, and with 200 mM NaCl the NI increased from 11.5% to 20%. A decrease was observed for PE under saline conditions from 60 to 49% with 100 mM NaCl, and from 60 to 47% with 200 mM of NaCl.

Membrane lipids of Bacillus toyonensis COPE52 growth in LB medium supplemented (or not) with different salt concentrations: 0 mM (a), 100 mM (b), and 200 mM NaCl (c)
The FA composition of COPE52 strain is shown in Table 2. The major FA was stearic acid (18:0), palmitoleic acid (16:1 ∆ 9) and 14-methyl pentadecanoic acid (16:0i) in control conditions without salt. The FA composition changed in response to increasing salt concentrations: with 100 mM NaCl, the relative amount of stearic acid (18: 0) decreased from 30.3 to 16.09%, whereas myristic acid (14: 0) increased from 10.87% to 16.23% and the branched chain FA 13-methyl tetradecanoic acid (15: 0i) and 15-methyl hexadecanoic acid (17: 0i) increased from 7.89 to 12.07%, and from 5.81 to 16.09%, respectively. At 200 mM NaCl, 14-methyl pentadecanoic acid decreased from 18.58 to 7.84%, while palmitoleic acid (16: 1 ∆ 9) declined from 19.21 to 8.44%, myristic acid (14: 0) and stearic acid (18: 0) declined from 10.87 to 6.47% and from 30.03 to 12.28%, respectively. On the other hand, palmitic acid increased from 7.61 to 16.46%, as did the branched chain FA 13-methyl tetradecanoic acid (15: 0i) and 15-methyl hexadecanoic acid (17: 0i) whose percentages increased from 7.89 to 24.24%, and from 5.81 to 24.27%, respectively. In summary, changing growth conditions applied to COPE52 caused a shift in the ratio of saturated to branched chain FA (B/S in Table 2), which increased from 0.66 to 1.02 under 100 mM NaCl condition and from 0.66 to 1.6 in 200 mM NaCl.
Biocontrol and Potential PGP Traits of Bacillus toyonensis COPE52
The potential of biocontrol and plant growth-promoting determinants was analysed in COPE52, such as siderophore excretion, protease activity and biofilm formation, in addition to the production of IAA (Table 3). Bacillus toyonensis COPE52 did not produce siderophores, and the production of IAA was maintained without significant difference with or without NaCl, while the biofilm formation was slightly increased in the presence of additional salt, specifically at 200 mM of NaCl. The proteolytic activity was the only activity that suffered a decrease (Table 3).
The antagonism experiment in no-salt conditions showed that the strain COPE52 inhibits the growth of the mycelium of Botrytis cinerea through diffusible and volatile compounds, by 10.89% and 18.12%, respectively. This degree of antagonism was maintained in saline conditions with 100 mM NaCl (10.71%), but not at 200 mM during the direct co-inoculation experiment (3.06%). Interestingly, volatile compounds of COPE52 were able to significantly inhibit the diameter of the B. cinerea mycelium with or without salt. With respect to the phytopathogen F. oxysporum, the strain B. toyonensis COPE52 restricted the diameter of the mycelium only through diffusible compounds and under control conditions (8.23%) and with salt at 100 mM (10.61%) (Table 4). These results indicate that strain COPE52 maintains its biocontrol and PGP in saline conditions, mainly at the concentration of 100 mM.
Effect of Bacillus toyonensis COPE52 Inoculation on Tomato Plants Under Salt Stress
To further evaluate the plant growth-promoting capacity of B. toyonensis COPE52, tomato plants were inoculated with strain COPE52 under three different conditions (0, 100 and 200 mM NaCl) (Fig. 2a). The inoculation of COPE52 significantly improved the root and shoot length, as well as the chlorophyll concentration, while the dry weight remained unchanged in control conditions. In the case of plants subjected to salt stress at 100 mM, it was observed that the length of the root and the stem were significantly increased. The chlorophyll concentration was similar but interestingly, in the presence of COPE52 the biomass (dry weight) of the tomato plants under these salinity conditions was increased. At 200 mM, the strain of B. toyonensis alone increased the length of the root significantly (Fig. 2). The factor analysis confirmed that in all cases the NaCl and COPE52 are highly significant, as well as in the interaction, compared to the controls without salt and uninoculated (Table 5). It is also clear that COPE52 protected plant growth, measured as dry weight, mainly at a concentration of 100 mM NaCl, while at 200 mM NaCl the effect was discreet or absent (Table 5 and Fig. 2).This result suggests that COPE52 have the ability to promote certain growth parameters of tomato plants under saline conditions (100 mM of NaCl).

Effect of inoculation of isolate COPE52 on plant growth and biomass content under different treatments; 0 mM, 100 mM, and 200 mM NaCl. Representative plants of the experiment are shown in Panel (a). Graphics show the root length (b), shoot length (c), chlorophyll concentration (d) and total dry weight (e). The bars represent the values of the mean ± SE. Letters indicate that the means differ significantly according to Duncan's multiple range test (P < 0.05)
Discussion
Bacteria regulate the fluidity of their membrane in response to several abiotic factors, including saline conditions [27]. Here, we analysed the phospholipid (PL) and fatty acid (FA) concentrations in the plant growth-promoting endophytic bacterium Bacillus toyonensis COPE52, and how its direct and indirect PGP mechanisms remain active under salt stress conditions.
Phospholipids are a major component of bacterial membranes, and changes in membrane lipid composition can affect important cellular processes such as metabolism, stress response, antimicrobial resistance, and virulence [28]. The COPE52 strain modified the phospholipids composition under tested conditions by increasing an unknown lipid (NI) and decreasing phosphatidylethanolamine (PE) synthesis. This unknown lipid migrated similarly to PC, but did not stain with Dragendorff reagent which is specific for betaine groups, meaning that the accumulated lipid is not PC. Paulicci and collaborators [13], showed that Ochrobactrum intermedium L115 presents with increased PC levels under high temperature and salinity (37 °C plus 300 mM NaCl), and this increase of PC could indicate an important adaptive mechanism for maintaining the structure and function of the membrane under stress conditions. However, changes in the amount of CL in the membranes, is perhaps the most important mechanism for bacterial adaptation to environmental stress [29]. For example, CL accumulates in E. coli cells in response to osmotic stress, which in turn promotes the polar localization of transporter ProP. ProP is denoted an osmosensory transporter because it is activated by increasing osmolarity and regulates the concentrations of organic osmolytes as proline in the cytoplasm [15]. López et al. [30], described a CL-deficient Bacillus subtilis strain, finding that this mutant is unable to grow in high salt concentrations. This result clearly indicates that the presence of CL in the membranes of B. subtilis is important for high salinity adaptation. The other adaptive mechanism often used by bacteria is alteration of membrane fatty acids (FA) [31]. Bacillus toyonensis COPE52 increased its proportion of branched chain FA in the presence of 100 and 200 mM NaCl. The increase in the proportion of branched FA increased the fluidity and permeability of the membrane in saline conditions [32], therefore, it is important to present other mechanisms that allow the cells to adapt to salinity. Haque and Russell [33], determined changes at the level of FA in seven strains of Bacillus cereus, finding that strain BR2853-5, when grown in a medium with NaCl (7%, w/v), increases to more than double the amount of the branched FA 13-methyl tetradecanoic acid (15: 0i). Such a report is consistent with the results obtained in this study. However, the changes that occur at the level of membrane FA in bacteria are generally conditioned by the type of stress to which they are subjected [34], that determined the composition of fatty acids of Bacillus thuringiensis under triclosan stress, where results showed increased concentrations of myristic acid, palmitoleic acid, palmitic acid and linoleic acid in the treatment samples with triclosan, as opposed to results in COPE52 wherein, the unsaturation level decreased. This indicates that the observed adaptative mechanism presented at the FA level are different under several stress conditions. It is important to mention that, myristic acid increased its production at 100 mM NaCl, but decreased when COPE52 cells were subjected to 200 mM NaCl. It has been observed with other bacilli strains that myristic acid similarly decreases its proportion as cell grow in media supplemented with salt (200 mM) (Rojas-Solís et al., 2020. Unpublished results). An explanation for this result is not known at this point.
The ability of several Bacillus species to antagonize phytopathogens, as well as promoting the growth of crop plants has been widely documented [35]. However, there are few reports about the beneficial effects exerted by strains of B. toyonensis on plants [19,20,21]. For example, Lopes and colleagues [19] described in silico, several genes for the synthesis of antimicrobial compounds, including bacteriocins, non-ribosomal peptides and chitinases during an in silico analysis of the B. toyonensis BAC315 genome, but no experimental evidence was evaluated. Recently, the draft genome sequence of the B. toyonensis COPE52 was reported, which presented its stimulating growing effects on blueberry plants (Vaccinium spp. var. Biloxi) in a greenhouse experiment [21]. The authors also detected the production of indoleacetic acid and volatile compounds like acetoin, 2,3-butanediol, as potential direct plant growth-promoting mechanisms. Interestingly, the strain COPE52 also produced the volatile compound and dimethyl disulfide, which has been associated with antifungal action, in particular, against the grey mould phytopathogen B. cinerea. Similar results were observed here, as the strain COPE52 exhibited better antagonism against B. cinerea and F. oxysporum through emission of volatile compounds. The ability of B. toyonensis COPE52 to promote the growth of tomato plants under saline conditions, mainly observed in growing plants at 100 mM NaCl, could be attributed to production of IAA and the volatile compounds acetoin and 2,3-butanediol. Several studies have shown that these compounds are responsible of stimulating growth of Arabidopsis and Canola plants [36, 37]. In fact, overproduction of Indole acetic acid in the rhizobacterium Pseudomonas sp. UW4 showed enhanced root elongation under gnotobiotic conditions [37]. In recent reports, the role of dimethyl disulfide as a modulator compound for the root system in Arabidopsis via auxin signalling pathway was elucidated [38].
Interestingly, the formation of a biofilm in B. toyonensis COPE52 was increased under saline conditions. Biofilm production has been proposed as a strategy adopted by bacterial strains for their successful biocontrol and survival in a plant rhizosphere and attachment to its roots. Biofilm production is a highly conserved mechanisms within the genus Bacillus under stressing growing conditions [39, 40]. In a report by Kasim et al.[39], the biofilm production of 20 bacterial isolates, mostly belonging to Bacilli, was evaluated under different salt concentrations. The authors observed that a large number of bacteria increased the production of biofilm presence of salt in the medium. Similar results were observed here, since B. toyonensis COPE52 increases the amount of biofilm as the salt was increased in the medium. Thus, biofilm production is also a potential strategy for protection and survival against toxic effects of the salt. In addition, it has been suggested that biofilm production is important to perform better plant growth-promoting activities in diverse rhizobacterial species [41].
Conclusion
The plant growth-promoting bacterial endophyte Bacillus toyonensis COPE52 modified the membrane profile of its PL, which potentially contributes to its adaptation to saline conditions and protection of plant growth-promoting mechanisms. Future research priorities include identifying the unknown membrane lipid NI and constructing COPE52 mutants in genes involved in the synthesis of specific phospholipids and fatty acids production, in order to unveil specific roles of each component of the cell membrane.
References
Bui E (2013) Soil salinity: a neglected factor in plant ecology and biogeography. J Arid Environ 92:14–25. https://doi.org/10.1016/j.jaridenv.2012.12.014
Yensen P (2008) Halophyte uses for the twenty-first century. In: Khan MA, Weber DJ (eds) Ecophysiology of high salinity tolerant plants. Springer, Dordrecht, pp 367–396
Shafi M, Guoping Z, Bakht J, Khan M, Islam E, Khan M, Raziuddin GZ (2010) Effect of cadmium and salinity stresses on root morphology of wheat. Pak J Bot 4:2747–2754. https://doi.org/10.1007/s00128-009-9707-7
Pierzynski M, Sims T, Vance F (2005) Soils and environmental quality. CRC Press, Florida
Paul D (2013) Osmotic stress adaptations in rhizobacteria. J Basic Microbiol 53:101–110. https://doi.org/10.1002/jobm.201100288
Orcutt M (2000) The physiology of plants under stress: soil and biotic factors. Wiley, New York
Dimkpa C, Weinand T, Ash F (2009) Plant-rhizobacteria interactions alleviate abiotic stress conditions. Plant Cell Environ 32:1682–1694. https://doi.org/10.1111/j.1365-3040.2009.02028.x
Wang W, Vinocur B, Altman A (2003) Plant responses to drought, salinity and extreme temperatures: towards genetic engineering for stress tolerance. Planta 218:1–14. https://doi.org/10.1007/s00425-003-1105-5
Qadir M, Oster D (2004) Crop and irrigation management strategies for saline-sodic soils and waters aimed at environmentally sustainable agriculture. Sci Total Environ 323:1–19. https://doi.org/10.1016/j.scitotenv.2003.10.012
Shrivastava P, Kumar R (2016) Soil salinity: a serious environmental issue and plant growth promoting bacteria as one of the tools for its alleviation. Saudi J Biol Sci 22:123–131. https://doi.org/10.1016/j.sjbs.2014.12.001
Paul D, Lade H (2014) Plant-growth-promoting rhizobacteria to improve crop growth in saline soils: a review. Agron Sustain Dev 4:737–752. https://doi.org/10.1007/s13593014-0233-6
Ma O-M, Duan J, DiBernardo M, Zetter E, Glick C-G et al (2019) The production of ACC deaminase and trehalose by the plant growth promoting bacterium Pseudomonas sp. UW4 synergistically protect tomato plants against salt stress. Front Microbiol 10:1392. https://doi.org/10.3389/fmicb.2019.01392
Paulucci N, Gallarato A, Reguera B, Vicario C, Cesari B, García B, Dardanelli S (2015) Arachis hypogaea PGPR isolated from Argentina soil modifies its lipids components in response to temperature and salinity. Microbiol Res 173:1–9. https://doi.org/10.1016/j.micres.2014.12.012
Vences-Guzmán MA, Guan Z, Ormeño-Orrillo E, González-Silva N, López-Lara I, Martínez-Romero E, Geiger O, Sohlenkamp C (2011) Hidroxylated ornithine lipids increase stress tolerance in Rhizobium tropici CIAT899. Mol Microbiol 79:1496–1514. https://doi.org/10.1111/j.1365-2958.2011.07535.x
Sohlenkamp C, Geiger O (2016) Bacterial membrane lipids: diversity in structures and pathways. FEMS Microbiol Rev 40:133–159. https://doi.org/10.1093/femsre/fuv008
Paulucci N, Medeot D, Dardanelli S, de Lema M (2011) Growth temperature and salinity impact fatty acid composition and degree of unsaturation in peanut-nodulating rhizobia. Lipids 46:435–441. https://doi.org/10.1007/s11745-011-3545-1
Santoyo G, Sánchez-Yáñez J, de los Santos-Villalobos S (2019) Methods for detecting biocontrol and plant growth-promoting traits in rhizobacteria. In: Reinhardt D, Sharma A (eds) Methods in rhizosphere biology research. Rhizosphere Biology. Springer, Singapore, pp 133–149
Kim M, Radhakrishnan R, Kang S, You Y, Jeong E, Kim J, Lee J (2017) Plant growth promoting effect of Bacillus amyloliquefaciens H-2-5on crop plants and influence on physiological changes in soybean under soil salinity. Physiol Mol Biol Plants 23:571–580. https://doi.org/10.1007/s12298-017-0449-4
Lópes R, Cerdeira L, Tavares S, Ruiz C, Blom J, Horácio A, Mantovani C, Queiroz V (2017) Genome analysis reveals insights of the endophytic Bacillus toyonensis BAC3151 as a potentially novel agent for biocontrol of plant pathogens. World J Microbiol Biotechnol 33:185. https://doi.org/10.1007/s11274-017-2347-x
Rocha F, Oliveira C, Silva P, Melo L, Carmo M, Baldani J (2017) Taxonomical and functional characterization of Bacillus strains isolated from tomato plants and their biocontrol activity against races 1, 2 and 3 of Fusarium oxysporum f. sp. Lycopersici. Appl Soil Ecol 120:8–19. https://doi.org/10.1016/j.apsoil.2017.07.025
Contreras-Perez M, Hernández-Salmerón J, Rojas-Solis D, Rocha-Granados C, Ma O-M, Parra-Cota FI, Santos-Villalobos S, Santoyo G (2019) Draft genome analysis of the endophyte, Bacillus toyonensis COPE52, a blueberry (Vaccinium spp. Var. Biloxi) growth-promoting bacterium. 3 Biotech 9:370. https://doi.org/10.1007/s13205-019-1911-5
Bligh G, Dyer W (1959) A rapid method of total lipid extraction and purification. Can J Biochem Physiol 37:911–917
Morrison W, Smith L (1964) Preparation of fatty acid methyl esters and dimethylacetals from lipids with boron fluoride. J Lipid Res 5:600–608
Patten C, Glick B (2002) Role of Pseudomonas putida indole acetic acid in development of the host plant root system. Appl Environ Microbiol 68:3795–3801. https://doi.org/10.1128/aem.68.8.3795-3801.2002
Hernández-León R, Rojas-Solís D, Contreras-Pérez M, Ma O-M, Macías-Rodríguez I, Reyes-de la Cruz H, Valencia-Cantero E, Santoyo G (2015) Characterization of the antifungal and plant growth-promoting effects of diffusible and volatile organic compounds produced by Pseudomonas fluorescens strains. Biol Control 81:8–92. https://doi.org/10.1016/j.biocontrol.2014.11.011
Wei H, Zhang L (2006) Quorum-sensing system influences root colonization and biological control ability in Pseudomonas fluorescens 2P24. Antonie Van Leeuwenhoek 89:267–280. https://doi.org/10.1007/s10482-005-9028-8
Gunde-Cimerman N, Plemenitaš A, Oren A (2018) Strategies of adaptation of microorganisms of the three domains of life to high salt concentrations. FEMS Microbiol Rev 42:353–375. https://doi.org/10.1093/femsre/fuy009
Kuhn S, Slavetinsky C, Peschel A (2015) Synthesis and function of phospholipids in Staphylococcus aureus. Int J Med Microbiol 305:196–202. https://doi.org/10.1016/j.ijmm.2014.12.016
Ti-Yu L, Douglas W (2016) Organization and function of anionic phospholipids in bacteria. Appl Microbiol Biotechnol 100:4255–4267. https://doi.org/10.1007/s00253-016-7468-x
Lópes C, Alice A, Heras H, Rivas E, Sánchez-Rivas C (2006) Role of anionic phospholipids in the adaptation of Bacillus subtilis to high salinity. Microbiology 152:605–616. https://doi.org/10.1099/mic.0.28345-0
Ramos J, Duque E, Gallegos M, Godoy P, Ramos-González M, Rojas A, Teran W, Segura A (2002) Mechanisms of solvent tolerant in gram negative bacteria. Ann Rev Microbiol 56:743–768. https://doi.org/10.1146/annurev.micro.56.012302.161038
Murínova S, Decová K (2014) Response mechanisms of bacterial degraders to environmental contaminants on the level of cell walls and cytoplasmic membrane. Int J Microbiol 2014:873081. https://doi.org/10.1155/2014/873081
Haque A, Russell J (2004) Strains of Bacillus cereus vary in the phenotypic adaptation of their membrane lipid composition in response to low water activity, reduced temperature and growth in rice starch. Microbiology 150:1397–1404. https://doi.org/10.1099/mic.0.26767-0
Li Y, Li C, Qin H, Yang M, Ye J, Long Y, Ou H (2018) Proteome and phospholipid alteration reveal metabolic network of Bacillus thuringiensis under triclosan stress. Sci Total Environ 615:508–516. https://doi.org/10.1016/j.scitotenv.2017.10.004
Santoyo G, Ma O-M, Govindappa M (2012) Mechanisms of biocontrol and plant growth-promoting activity in soil bacterial species of Bacillus and Pseudomonas: a review. Biocontrol Sci Technol 22:855–872. https://doi.org/10.1080/09583157.2012.694413
Ryu M, Farag A, Hu H, Reddy S, Wei X, Paré W, Kloepper W (2003) Bacterial volatiles promote growth in Arabidopsis. Proc Natl Acad Sci USA 100:4927–4932. https://doi.org/10.1073/pnas.0730845100
Duca R, Rose R, Glick B (2018) Indole acetic acid overproduction transformants of the rhizobacterium Pseudomonas sp. UW4. Antonie Van Leeuwenhoek 111:1645–1660. https://doi.org/10.1007/s10482-018-1051-7
Tyagi S, Kim K, Cho M, Lee J (2019) Volatile dimethyl disulfide affects root system architecture of Arabidopsis via modulation of canonical auxin signaling pathways. Environ Sustain 2:211–216. https://doi.org/10.1007/s42398-019-00060-6
Kasim A, Gaafar R, Abou-Ali M, Omar N, Hewait M (2016) Effect of biofilm forming plant growth promoting rhizobacteria on salinity tolerance in barley. Ann Agric Sci 61:217–227. https://doi.org/10.1016/j.aoas.2016.07.003
Bais H, Fall P, Vivanco M (2004) Biocontrol of Bacillus subtilis against infection of Arabidopsis roots by Pseudomonas syringae is facilitated by biofilm formation and surfactin production. Plant Physiol 134:307–319. https://doi.org/10.1104/pp.103.028712
Seneviratne G, Weerasekara MLMAW, Seneviratne KACN, Zavahir JS, Kecskés ML, Kennedy IR (2010) Importance of biofilm formation in plant growth promoting rhizobacterial action. In: Maheshwari D (ed) Plant Growth and health promoting bacteria. Springer, Berlin, pp 81–95
Acknowledgements
We thank Eduardo Valencia-Cantero for the help with the statistical analyses. This study was funded by Consejo Nacional de Ciencia y Tecnología, México (Grant number: A1-S-15956) and CIC-UMSNH (2019–2020). DR-S thanks a Ph.D. scholarship from Consejo Nacional de Ciencia y Tecnología, México. We also thank the anonymous reviewers for helpful comments.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Rojas-Solis, D., Vences-Guzmán, M.A., Sohlenkamp, C. et al. Bacillus toyonensis COPE52 Modifies Lipid and Fatty Acid Composition, Exhibits Antifungal Activity, and Stimulates Growth of Tomato Plants Under Saline Conditions. Curr Microbiol 77, 2735–2744 (2020). https://doi.org/10.1007/s00284-020-02069-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00284-020-02069-1