
Abstract
Gram-negative bacteria have the ability to produce outer membrane-derived vesicles (OMVs) that are released into the extracellular milieu. Even though this intriguing phenomenon is well-known since many years, various aspects of bacterial OMVs are not fully described and are still in the process of being characterized in detail. One major reason for this is that depending on the bacterial species and its respective ecological niche, OMVs exhibit an enormous functional diversity. Research of the past years has clearly shown that OMVs of many pathogenic bacteria contribute to the virulence potential by enriching virulence factors and delivering them over long distances, superseding direct bacterial contact with their host. The subsequent interaction of OMVs with the host can occur at different levels regarding the type of immune response or the target cell type and may lead to different outcomes ranging from non-immunogenic activation or a pro-inflammatory response to cytotoxicity. In contrast to being virulence factors, OMVs are used for vaccination purposes in the combat against bacterial pathogens, and recent research thus is focused on to indirectly aim these versatile bacterial weapons against themselves.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Bacterial outer membrane vesicles (OMVs) are spherical, bilayered, membranous structures released from the surfaces of Gram-negative bacteria [1]. Their first description dates back more than 40 years, when Bishop et al. analyzed the culture supernatants of lysine-requiring Escherichia coli mutants [2]. The scientists were perplexed to find that these mutants under lysine-limiting conditions produced an excess of material they at that time called “extracellular lipoglycopeptide.” Further biochemical and ultrastructural studies revealed that this material was in its composition and appearance similar to the bacterial outer membrane and that the OMVs, or blebs, were produced under common laboratory culture conditions [3–6]. It has later been shown that these vesicles are not products of cell lysis but that they are generated during logarithmic cell growth as well as in stationary phase (Fig. 1). OMVs are sized between 50 and 250 nm in diameter and contain componens of the outer membrane of the bacteria, like lipopolysaccharide (LPS), lipids, adhesins, invasins, toxins, other virulence factors, and sometimes even bacterial DNA. They are associated with a broad array of functions ranging from virulence and metabolism to interspecies communication [7, 8].

Moraxella catarrhalis produces outer membrane vesicles (OMVs) or “blebs” both in vitro as well as in vivo. a Large amounts of OMVs can be seen as small blebs protruding from the bacterial cells. b Direct examination of M. catarrhalis shows secretion of OMVs carrying UspA1/A2 as revealed by a gold-conjugated rabbit anti-UspA1/A2 antiserum. c–e Secretion of OMVs during Moraxella infection in a 9-year-old child with sinusitis. In c, a bacterium and OMVs are seen in the vicinity of a polymorph. In e, UspA1/A2 expression in vivo is verified by the gold-labeled antiserum directed against UspA1/A2. All panels represent pictures obtained by transmission electron microscopy. Adapted from [21] ©2007 by the Infectious Diseases Society of America. Reproduced by permission
Biogenesis of OMVs
Despite the early discovery and the extensive research on the isolation of OMVs, the molecular mechanisms behind the production and shedding of OMVs are still not completely understood. A recent review depicts in detail this particular aspect of OMVs [9]. Hence, only an introductory overview of OMV biogenesis will be presented in this chapter.
The production of OMVs is not limited to certain growth conditions, and OMVs can be observed with bacteria growing in vitro on solid agar [10], in liquid medium [11–14], within biofilms [15, 16], during intracellular infection [17], as well as in vivo [18–21]. Nevertheless, the amount of OMV production can vary due to environmental changes. In particular, stress factors such as treatment with the antibiotics gentamicin [22] or chloramphenicol [5], limitation of lysine [2, 4], Mg2+ [23], hemin [24] or phage infections [25], as well as envelope stress due to the accumulation of unfolded proteins in the periplasm [26] cause an overproduction of OMVs.
Any structural protein machinery solely dedicated to the generation of OMVs has not been described to date. Studies in this direction are further complicated by the fact that no mutant bacteria exist, which is completely devoid of vesicle production [27, 28]. Generally, it is assumed that detachment of the outer membrane from the underlying peptidoglycan (PG) is the initial step for the formation of a vesicle [29]. This can be favored at regions where the growth of the outer membrane is faster than the cell wall. These so-called insertion sites where newly synthesized material is incorporated into the outer membrane exist along the bacterial cell membrane at distinct foci [30]. In another model, imbalance in the turnover of PG leads to accumulation of low molecular weight muramic acid, which causes a bulging of the outer membrane by the turgor generated [31]. This is supported by the fact that OMVs contain low molecular weight muramic acid [6, 22, 31]. Finally, other studies showed that also the negative charged nature of the outer membrane might influence the OMV production. For example, Pseudomonas aeruginosa possesses a higher blebbing rate when the bacterium has an outer membrane enriched in more negatively charged B-band LPS presumably due to the stronger repellent forces generated [22, 32, 33]. This is also in accordance with previous observations that limitation of Mg2+, a stabilizer of lipid membranes, enhances vesiculation [23, 34].
Although the initiation of OMV formation can be described by different models without really excluding each other, the localized absence of outer membrane proteins covalently or non-covalently bound to PG, the inner membrane, or other membrane proteins is most certainly crucial for the completion of the process. One early observation supporting this was that lipoproteins were underrepresented in OMVs compared with the outer membrane [29, 35]. Subsequent and independent studies in E. coli [28, 36], Porphyromonas gingivalis [37], and Salmonella typhimurium [38] concordantly identified OmpA, LppAB, and TolA/B as major membrane proteins that stabilize the outer membrane via protein–protein or protein–PG interactions. Deletion of these either increased the vesiculation rate or resulted in oversized vesicles. Intriguingly, Neisseria meningitidis, which is known to produce high amounts of OMVs, seems to lack homologues of the aforementioned membrane proteins [38].
Bacteria produce OMVs in vivo during infection
Several examples exist of interactions in vitro between OMVs and both gastric and mucosal epithelium, as well as several parts of the immune system. The relevance of these studies depends on the assumption that OMVs are produced in vivo, which in fact has been shown in a number of studies. Probably, the earliest proof for the in vivo production of OMVs came from an infant suffering from N. meningitidis. Meningococci and their blebs were observed in negatively stained cerebrospinal fluid of this patient and were proposed to contribute to the disease [20]. In another study, LPS in plasma from a lethal shock patient was quantified using gas chromatography and mass spectrometry. Some of the LPS circulated in plasma as complexes with high sedimentation coefficients, almost always in combination with outer membrane fragments, indicating the presence of OMVs in the serum [18]. Also in another case study where high levels of endotoxins were found in the serum of a severe shock patient with multiple organ failure, cultures were positive for N. meningitidis serogroup B and the in vivo release of OMVs could be proven by electron microscopy. Again the authors concluded that LPS-carrying OMVs contributed to the fatal endotoxin levels in the serum of the patient [39]. Furthermore, it was shown that OMVs were produced by clinical Helicobacter pylori isolates in vivo, and that these contained serologically recognizable Lewis antigens on their surface [19]. Another example for the in vivo production of OMVs comes from a study, where nasal swaps of a child suffering from Moraxella sinusitis were analyzed by electron microscopy (Fig. 1c–e) [21]. In this case, the co-existence of OMVs together with extracellular bacteria and host immune cells could clearly be shown. Also the presence of ubiquitous surface proteins (Usp) A1 and A2, two major virulence factors of Moraxella catarrhalis, on these vesicles was demonstrated by using gold-labeled antibodies. In a more recent publication from our laboratory, it was also demonstrated that the superantigen Moraxella IgD-binding protein (MID) exists on OMVs seen in children [40].
OMVs deliver virulence factors and contribute to bacterial colonization
The discovery that bacteria are able to produce considerably large amounts of OMVs was initially puzzling, since this must come along with high rates of lipid and protein biosynthesis, which should cause a homeostatic disadvantage for the bacterium in question. However, the more the biological functions associated with OMVs were unraveled the more it became apparent that OMVs are crucial for the successful establishment of a bacterial species in its anatomical niche. Thus, in the past, much research focused on OMVs of bacterial pathogens and their functions relevant for virulence.
In order to colonize the host, it is essential for the bacteria to adhere to the epithelial cell membrane in the gut or in the respiratory tract in a way that it also resists physical removal. A wide array of bacterial virulence factors is used by the bacteria, which help them to invade cells, compete with the host for iron and other nutrients and evade or counteract the innate immune defense system of the host. But what is the function of OMVs in the context of bacterial infection?
It has been hypothesized that OMVs can act as an alternative secretion system, similar to type III secretion systems (TIIISS) of bacteria, like Salmonella and Yersinia, or type IV (TIVSS) secretion systems of Legionella or Helicobacter, which are utilized by these species to colonize their hosts or establish intracellular infection [41, 42]. Similarly, OMVs can be considered as a secretion system, delivering proteins and virulence factors to target cells. Clearly exemplifying the role of OMVs as a transport system, the OMVs from the Gram-negative pathogen Shigella flexneri was shown to pick up and entrap the antibiotic gentamicin, and subsequently deliver it to the cytoplasm of host epithelial cells [43]. In contrast to TIIISS or TIVSS, OMVs can act over a long distance, allowing the pathogen to hide at a distance from the site of infection, still allowing it to evade or distract the host immune response. By using OMVs, the pathogen is also able to inject virulence factors across host membranes and into compartments of cells, leading to destruction of the target cells.
Another advantage of OMVs is that they may provide means for bacterial proteins to be transported in complex with other bacterial factors while being protected from degradation. In OMVs, bacteria can transport toxins, enzymes, as well as DNA to eukaryotic cells in a protected manner. Several studies have aimed at comparing the protein composition of OMVs and the outer membranes of the pathogen they derive from (Table 1). For example, a proteomic analysis of H. pylori OMVs showed that the adhesins BabA and SabA were more abundant in the outer membrane compared to the OMVs [44]. In contrast, the chaperone and serine protease HtrA was enriched in the vesicles compared to the outer membrane. In another recent study, H. pylori HtrA was shown to be a potentially important virulence factor since this protein cleaves the cell adhesion protein E-cadherin. This leads to a disruption of the barrier functions of the epithelium, allowing the pathogen to access the intercellular space [45].
An important aspect of OMVs is that they have been found to contain RNA as well as circular or linear DNA [46–49]. Vesicles from P. aeruginosa, Haemophilus influenzae, M. catarrhalis, and Neisseria gonorrhoeae were all found to contain DNA in various forms, either surface-associated or inside the lumen of the OMVs. Vesicles from N. gonorrhoeae can transfer plasmid DNA both from inside the bacteria as well as exogenous DNA taken up from the extracellular space to other gonococci [50]. Renelli et al. in turn showed that in P. aeruginosa OMVs, DNA could be derived from the periplasm of the cell or the extracellular space. However, the authors were not able to transform neither Pseudomonas nor E. coli cells under a variety of transforming conditions [49]. In the case of Moraxella, this species spreads CpG-motif containing bacterial DNA attached on the surface of OMVs [40]. Any DNA cannot, however, be found inside the vesicles that are derived from M. catarrhalis.
OMVs can also be found in biofilms in natural environments and may contribute to the establishment of biofilms by mediating adherence, delivering extracellular material, or utilizing growth factors [1, 15, 16, 51]. The heme-utilization protein HmuY of P. gingivalis was shown to be associated with OMVs and beneficial for the biofilm formation [52]. Kulp and Kuehn hypothesized that OMVs are an important part of the microflora as part of the biofilm [53]. However, these authors also claimed that OMVs lyse surrounding foreign bacteria in the biofilm, in a situation when nutrients are scarce and during hostile growth conditions. Using a peptidoglycan hydrolase, as it is the case for P. aeruginosa, this would give the OMV-producing bacterial strain an important survival advantage at the expense of the surrounding bacteria [53, 54]. On the other hand, OMVs from species such as P. aeruginosa and M. catarrhalis have been established to carry proteins of the β-lactamase family, which break down β-lactam rings such as those found in amoxicillin [55] [Schaar et al., in preparation]. By this mechanism, the bacteria might transfer antibiotic resistance in biofilms, as well as protect other bacteria from antibiotic killing.
OMV-dependent interaction with the mucosa and epithelial cells
Outer membrane vesicles have a protein composition reflecting that of the outer membrane of the parent bacteria. Considering this, OMVs have the potential to bind to epithelial cells in the respiratory tract as well as in the mucosa, anatomical sites where bacteria interact with their host.
H. pylori is an illustrative example of a pathogen that is residing in the mucosa and colonizes the gut of about 50% of the human population. In most patients, this infection is asymptomatic; but in about 20% of patients, the infection leads to chronic gastritis, as well as, an increased risk of developing peptic ulcer disease, gastric lymphoma, or gastric carcinoma. Surprisingly, it has been found that H. pylori are a non-invasive pathogen and even though the species has the ability to adhere to epithelial cells it is often found to remain unattached. However, LPS-enriched OMVs secreted from the bacteria bind to and invade gastric epithelial cells partly through clathrin-mediated endocytosis. By adherence through adhesins BabA and SabA and the vacuolating cytotoxin VacA, OMV can help cause inflammation by the persistent delivery of antigens like proteases and urease to the gastric mucosa [44, 56]. OMVs have been found to modulate epithelial cell proliferation and apoptosis and stimulate secretion of inflammatory cytokines.
Another example where OMVs bind to epithelial cells is found with M. catarrhalis, a respiratory pathogen commonly causing otitis media in children and exacerbations in patients with COPD. Moraxella OMVs bind to TLR2 in lipid rafts of the epithelial cells, which are subdomains of the membrane with distinct protein and lipid compositions and enriched in cholesterol and sphingolipids [57]. The OMVs are subsequently internalized, causing a pro-inflammatory response resulting in increased IL-8 secretion and ICAM-1 expression. OMVs from M. catarrhalis contain among other proteins two of the main virulence factors of Moraxella, the adhesin UspA1 and the superantigen MID [58]. Slevogt et al. have previously shown that UspA1 interacts with the adhesion molecule CEACAM-1 on human cells, thereby interrupting the TLR2-dependent activation of the transcription factor NF-κB, which in turn leads to a pro-inflammatory response by epithelial cells [59]. Supporting these findings, our experiments revealed that OMVs deficient of UspA1 enhance the pro-inflammatory response. UspA1 in OMVs might thus have an important role in helping Moraxella to evade the host immune response [58]. Hence, OMVs can be seen as bacterial tools to interact with and regulate the inflammatory response of epithelial cells at a distant site from the localization of the parent bacteria.
Attachment of Legionella pneumophila OMVs to A549 lung alveolar epithelial cells has been observed, although the molecular mechanisms remain to be solved [17]. Following attachment of OMVs, the production of IL-6, 7, 8, and 13 as well as GM-CSF, IFN-γ, and MCP-1 was observed. Interestingly, IL-7 and the anti-inflammatory IL-13 were specific to the OMVs and were not produced when epithelial cells were incubated with whole bacteria. It was postulated that this could be due to the altered LPS composition of the OMVs, since EDTA and protease treatment did not alter the cytokine profile. OMVs were also observed in the phagosomes of internalized Legionella suggesting that OMVs might play a role for the intracellular survival of the bacteria. Once taken up, L. pneumophila typically circumvents phagolysosomal degradation with the help of virulence factors, which are secreted by the TIVSS into the cytosol of the host cell. However, also OMVs were shown to inhibit the fusion of the Legionella-containing phagosome with lysosomes independent of the TIVSS [60].
OMVs from several species including P. aeruginosa, Borrelia burgdorferi, Vibrio cholerae, and Vibrio vulnificus have all been shown to bind epithelial cells in vitro, delivering bacterial virulence factors to the membrane of host cells [55, 61–64]. Moreover, OMVs from V. vulnificus were found to cause cytotoxicity and induce cell death, mediated by the cytolysin VvhA [63]. In H. pylori, N. meningitidis and P. aeruginosa OMVs were shown to deliver peptidoglycan into epithelial cells, where it is recognized by the intracellular immune receptor NOD1. This in turn initiated an inflammatory response by the epithelial cells in form of IL-8 production (Fig. 2c). In H. pylori, similar to L. pneumophila, vesicles were identified as an alternative to the TIVSS, in this case for delivering peptidoglycan [65].

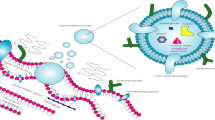
Bacterial OMVs mount various immune responses. a M. catarrhalis OMVs interact via the IgD-binding protein MID with surface IgD on B cells and are taken up by the additional interaction of lipoproteins with TLR2. This leads to lipid raft formation and subsequent uptake. TLR9, which recognizes OMV-associated DNA, together with TLR2 activates the B cells non-immunogenically resulting in IL-6 secretion. Additionally, B cells start producing non-specific IgM, which are not directed against Moraxella [40]. b Salmonella vesicles can be taken up by macrophages or dendritic cells (DC). This leads to activation and production of TNF-α, IL-12, and nitric oxide (NO). The subsequent endosomal processing of OMVs leads to upregulation of MHC class II and CD86 on the surface of macrophages and DC and triggers cell-mediated immunity [27]. c The uptake of OMVs by epithelial cells is in many bacterial species dependent on lipid raft formation and leads to a general pro-inflammatory response via NF-κB and p38-mediated signaling and IL-8 production. OMVs were shown to promote this response partly by the delivery of peptidoglycan, which is recognized by the intracellular PAMP-receptor NOD1 [65]. For further references see also Table 1
Enterotoxigenic E. coli, a major cause of diarrhea and infant mortality in third world countries, was also analyzed for OMV binding to epithelial cells. The heat-labile enterotoxin LT is secreted by the outer membrane of the bacteria as well as OMVs. LT-containing vesicles and OMVs lacking the toxin were compared for their capacity to invade adrenal and intestinal epithelial cells. It was clearly shown that only LT-carrying vesicles have the ability to invade cells in a time-, temperature-, and receptor-dependent manner. The vesicles attached to the cells at 4°C, and at 37°C, the LT was internalized [66]. This was mediated through cholesterol-rich lipid rafts, and OMVs sometimes co-localized with the endocytosis protein caveolin. Kesty et al. concluded that the OMVs are targeted transport vesicles of the enterotoxin.
Further investigating the mechanism behind uptake of OMVs, Furuta et al. characterized in detail how OMVs from P. gingivalis are taken up and sorted by human epithelial cells [67]. They established that OMVs bound to epithelial cell membranes in a fimbria-dependent manner and were endocytosed through actin-dependent lipid raft-mediated pathways, involving caveolin and dynamin. It was established that OMVs intracellularly were transported to lysosomal compartments where they were eventually degraded. However, OMVs persisted inside the cells for up to 24 h, and caused a large induction of acidic cellular compartments, suggesting cellular stress and impairment. In the case of P. gingivalis, this might suggest a possible mechanism for destruction of the peridontal tissue in patients with chronic peridontitis, eventually leading to tooth exfoliation. OMVs from P. gingivalis thus also exemplify a general mechanism whereby bacteria use OMVs as a powerful offensive weapon to survive within and in some cases fight their host.
Interaction of OMVs with the innate and adaptive immune system
Bacterial colonization of the human body is always accompanied by a host immune response. As a principle, bacteria interact with the human immune system in a number of ways and also use different proteins for binding the same protein or receptor, an efficient strategy resulting in several backup systems. Important parts of this immune response are phagocytic cells such as neutrophils and macrophages, activation of cells of the adaptive immune response including B and T cells, as well as activation of the complement system. In turn, bacteria have evolved different ways to survive inside the host in order to initiate colonization. Outer membrane vesicles have an important role in interacting with the innate immune system, and act both as pro- and anti-inflammatory mediators, perhaps depending on the specific strain and environmental circumstances (Fig. 2).
Several studies have shown that OMVs are decoys for the immune system by redirecting the inflammatory response. A specific example of OMVs interacting directly with immune cells has been shown with M. catarrhalis that activates B cells through a T cell independent mechanism. The interaction between the superantigen MID expressed on OMVs and IgD B cell receptors on the cell surface leads to receptor clustering in lipid rafts, and subsequently, uptake of the OMVs [40]. The membrane of the OMVs interacts with TLR2 and TLR9 sensing lipoproteins and DNA, respectively. This MID-dependent cross-linking of the IgD B cell receptor as well as TLR activation results in IL-6 synthesis and polyclonal IgM production (Fig. 2a). Interestingly, Moraxella OMVs containing MID as well as DNA could be found in specimens from a 6-year-old child, also proving this concept in vivo [40].
In the case of Moraxella, we demonstrated that OMVs interact with the alternative pathway of the complement system, a central part of the innate immunity. Three major pathways are recognized when the complement system is activated [68, 69]. These are (a) the classical pathway, activated by bacterial LPS, nucleic acids, and the antigen–antibody complex, (b) the lectin-mediated pathway, activated by binding of mannose-binding lectin to mannose residues on the bacterial surface, and finally, (c) the alternative pathway, activated by deposition of C3b after recognizing pathogen-associated molecular patterns at the bacterial surface and LPS. Pathogen recognition leads to opsonization of the pathogen and subsequently phagocytosis [70, 71]. In addition, all three pathways eventually lead to the common terminal pathway resulting in formation of a membrane-attack complex [69]. We found that Moraxella OMVs contain the virulence factors UspA1/A2. This multifunctional family of proteins in a dose-dependent manner binds and inactivates complement factor C3, one of the major players in the complement system, and hereby, restraining complement activation [21, 69, 72, 73]. Interestingly, we also observed that Moraxella OMVs carrying UspA1/A2 protected the respiratory pathogen H. influenzae from complement-mediated killing by a mechanism comprising inactivation of C3 [21]. These results suggest a novel, OMV-mediated strategy by which bacteria collaborate with each other in order to defeat innate immunity. Considering the importance of the complement system as an essential part of the innate immune system, leading to lysis of pathogens, opsonization, and an increase in the general inflammatory response, the described mechanisms using OMVs as a “first line of attack” give the bacteria an important advantage in evading the early innate immune response.
S. typhimurium produces OMVs that stimulate professional antigen presenting cells, like macrophages and dendritic cells in vitro [27, 74]. These cells showed an increased expression in MHC class II and induced production of inflammatory cytokines like TNF-α and IL-12 as well as the toxic nitric oxide through a TLR4-dependent and independent signaling pathway. The antigens on Salmonella OMVs were also found to have the capacity to prime both B cells and T helper cells (Fig. 2b).
H. pylori represents another way of OMV-dependent interaction with the immune system. In initial immunization studies with H. pylori LPS, the antibodies that were produced were found to be reactive against Lewis antigen structures on human blood cells. These antigens also commonly occur in the gastric mucosa [75]. Hence, H. pylori seems to express a protein similar to the Lewis structures in some cases of gastritis, thereby creating an autoimmune response in the host via molecular mimicry [75]. Later, it was determined that these antigens could also be found on OMVs secreted from the bacteria, specifically in cases of chronic gastritis and gastric cancer [19]. Additionally, Helicobacter OMVs, like shown for other bacteria, can be taken up by epithelial cells via lipid rafts. This leads to a general inflammatory response and the production of IL-8 (Fig. 2c).
Potential of OMVs as non-replicating vaccines
Bacterial infections are still one of the major causes for hospitalization and death worldwide [76–78]. The treatment of bacterial diseases is hampered due to an increased and rapid spreading of antibiotic resistant bacteria. Hence, vaccines are considered as one of the most straightforward strategies of the post-antibiotic era in the treatment of bacterial diseases [79, 80]. Soon after the discovery that OMVs carry antigenetically active virulence factors, their potential as non-replicating vaccines has become a major interest of immunotherapeutic research [81–85]. The number of studies focusing on OMVs as vaccination tools is steadily growing, and examples from many laboratories including the already existing OMV vaccines against N. meningitidis support the feasibility of this approach (Table 2). A closer look at the development of neisserial vaccines gives also an insight into the potentials and challenges of OMV-based vaccines.
N. meningitidis is an encapsulated Gram-negative diplococcus, which colonizes the nasopharyngeal mucosa of 5–10% of human beings without causing disease. However, N. meningitidis is also the major cause for bacterial meningitis epidemics affecting especially children beyond the neonatal period and young adults up to the age of 29 [86]. Its polysaccharide capsule allows the distinction between 13 different serogroups, of which the groups A, B, C, W135, and Y account for 90% of the cases. Out of these, group A meningococci are mostly associated with broad epidemics especially in the sub-Saharan region. On the other hand, in temperate and industrialized countries of North America and Europe serogroup B meningococci are the main cause for N. meningitidis accounting for 30–80% of the cases followed by serogroup C [86, 87]. Another striking feature of serogroup B epidemics is that they can persist for a decade and thus much longer than the epidemics of other serogroups.
The polysaccharide capsule proved to be a valuable tool for vaccination purposes in case of meningococci except for the serogroup B. Introduction of the polysaccharide conjugate vaccine against serogroup C (MenC) reduced the incidence by 90% [88, 89]. Similarly, a tetravalent vaccine (MCV4), where polysaccharide of the serogroups A, C, W135, and Y were covalently linked to diphtheria toxin, has been successfully applied in the USA for the age group 11–55 years old [90–92]. Hence, conjugate vaccines are effective in controlling meningococcal disease by inducing the production of bactericidal antibodies with a long lasting immunity due to the elicited T cell-dependent immune response [86].
The success of conjugate vaccines could, however, not be repeated in case of serogroup B meningococci. In early attempts, it was shown that polysaccharide of this serogroup fails to boost an immunological response [93]. Also the coupling to tetanus toxoid did not improve the antigenicity [94]. The reason for this lies in the nature of the capsular polysaccharide of serogroup B, which contains homopolymers of α2-8 linked sialic acid and by this closely resembles the oligosaccharide side chains of human brain tissue [95]. Also the approach of using outer membrane proteins as vaccines had to be abandoned due to low immunogenic response [85]. These drawbacks together with the increase in epidemics in Cuba, Norway, and New Zealand catalyzed the development of OMVs from epidemic disease strains as a last resort against serogroup B meningococci [96–99]. This finally led to the approval of the first OMV-based vaccine in Cuba, which achieved vaccination efficacies ranging from 83% to 94% with low reactogenicity and long-term protection [98], followed by trials in Norway and Brazil as well as the introduction of the MeNZB vaccine in New Zealand [100].
The initial OMV vaccine formulations based on the preparation of culture supernatants of the regional epidemic strain, and applied detergent extraction to remove excess LPS [87, 100]. Although this method aimed at reducing the endotoxicity and increasing tolerability during vaccination, it also had the unwanted side effect of removing most of the lipoproteins leaving basically PorA as the predominant antigenic molecule of the vaccine [101]. However, this neisserial porin is highly variable, and therefore, the bactericidal antibody response generated by a wild-type OMV vaccine confers protection only to one epidemic serotype [102, 103], which is currently the biggest obstacle in the development of a globally effective serogroup B vaccine. The next possible approach of mixing OMV preparations of different disease causing serotypes is due to the high variability of PorA not feasible. There are over 600 PorA variants, and, e.g., a vaccine covering 80% of serogroup B disease causing strains in the USA would need to include 20 different variants [104, 105]. Additionally, the protection of children under the age of 4 cannot be achieved with PorA dominant OMV vaccines [86]. Therefore, much effort is put into improving the efficacy of OMV vaccines by several approaches. Given that PorA alone is not enough for mounting a good protection, the search for alternative OMV-associated antigens has led to the identification of the neisserial adhesin A (NadA), the factor H binding protein (fHBP), neisserial heparin binding antigen (NHBA), or the neisserial surface protein A (NspA) [106, 107]. The combination of recombinantly expressed NadA, fHBP, and NHBA with OMVs of the New Zealand epidemic strain proved to be more efficient in the immunization of infants compared to OMVs or recombinant proteins alone [106].
Another recombinant approach aims at improving the isolation process of OMVs so that the usage of detergents for depleting LPS can be avoided, since this removes also most of the lipoproteins and by this, potential antigens. In three recent publications, these efforts towards the generation of native OMV vaccines were presented [101, 108, 109]. Deletion of the LPS-biosynthesis gene lpxL1 led in both cases to the production of shorter LPS with less toxicity, nevertheless, retaining its adjuvant activity. Additional deletion of rmpM increased the OMV yield, since this mutation causes an easier detachment of OMVs from the outer membrane [101, 110]. In a phase 1 of the clinical trial, the safety and immunogenicity of OMVs isolated form a lpxL2 and synX negative mutant epidemic strain 44/76 was assessed, and the tolerability of native vesicles with a due to lpxL2 deletion modified LPS in human volunteers was shown [108]. In the same study, serum bactericidal activity together with antibodies against LPS and the outer membrane protein OpcA could be detected. Deletion of the sialic acid synthesis gene synX was also applied in the third study, where the authors in addition to the aforementioned lpxL1 deleted a further LPS modification gene lgtA [109]. Interfering with the sialic acid synthesis, although it negatively affected the purification process and OMV yield, was introduced in order to prevent the production of human cross-reactive antibodies. Again LPS was the dominant antigen leading to the production of bactericidal antibodies in mice. The authors also went a step further and introduced recombinant fHBP and NadA variants into the tested vaccine strain and could confirm that NadA was also a potent antigen while antibody response towards fHBP was comparably lower [109].
The recent developments in neisserial OMV vaccines support the potential of OMVs as an alternative way of vaccination and also give an insight into the basics for OMV-based vaccinology, which then can be transferred to other bacterial pathogens. It is therefore not surprising that in case of many other pathogens the interest in identifying the vaccine potential of OMVs or OMV delivered antigens is growing.
While the exact immunological mechanisms still need to be elucidated in detail for each tested pathogen, a general observation is that the immune protection is mainly based upon bactericidal antibodies directed against one dominant or multiple antigens delivered by the OMVs (Table 2). This is clearly shown by a study, where S. enterica Typhimurium was genetically modified, so that it produced less amounts of toxic LPS. In addition, a viral epitope of the canine parvovirus (VP2) was introduced. When mice were immunized with these genetically modified OMVs bactericidal antibodies against Salmonella as well as neutralizing antibodies against VP2 could be detected [111]. In case of V. cholerae, several consecutive studies have proven that immunizing mice or rabbits by administering OMVs via several routes conferred long-term protection against the pathogen by the production of bactericidal IgA and IgG1. In this case, the main antigenic molecule was found to be LPS [62, 112–114]. Bordetella pertussis and B. burgdorferi are two other pathogens, where the protective effect of OMVs in challenged mice was experimentally demonstrated [115, 116]. In case of B. burgdorferi, the major protective antigens were OspA and DbpA, two adhesins responsible for colonization. In case of OspA, it could even be shown that the OMV-associated form is more immunoprotective than the purified protein [115]. In Treponema pallidum OMVs had a much higher killing titer compared with serum from rabbits immunized with whole bacteria [117].
Concluding remarks
OMVs of Gram-negative bacteria that were initially observed under peculiar growth conditions are now an inherent part of microbiology. In the last 40 years of research on OMVs, it has become clear that these “magic bullets” are a general phenomenon rather than an exception. The research on OMVs now covers bacterial physiology, pathogenesis, and ecology with OMVs as virulence factors certainly being one of the most interesting properties. OMVs clearly represent an alternative secretion and transport mechanism for many different proteins, especially for those where no already known and described secretion system can be accounted for. OMVs also allow the bacteria to act on and interact with their environment over a longer distance without risking the disadvantages of direct contact. This is particularly true in cases where bacteria might be able to induce unspecific immune reactions via OMVs, and thereby, circumvent a specific host response, or where bacteriocidal factors might be beneficial for the competition in complex polymicrobial niches.
OMVs have evolved from being an exceptional observation to an important tool of immunology by means of alternative vaccination agents against one of the most epidemic bacterial diseases, namely neisserial meningitis. The benefits of OMV-based vaccines for the control of serogroup B meningococci combined with more than 20 years of experience in clinical trials and approved vaccination programs strongly support the potential of these vaccines. Genetic manipulation, which aims at improving vesicle production and antigenicity as well as decreasing adverse effects, already shows first promising results in the case of neisserial vaccines, and points out the plasticity as well as future potential of this approach. OMV-based vaccines will hence certainly be an important strategy in combating further endemic or epidemic diseases, like cholera, where OMVs are shown to be effective immunogenic vehicles.
Proteomics is a great help in characterizing OMV contents of many bacteria and shows that they can be enriched by certain components when compared to the periplasm or even the bacterial outer membrane they originate from. This further suggests that the biogenesis of OMVs is a relatively controlled process. However, the precise mechanisms of OMV biogenesis and specific targeting of proteins to OMVs are still only poorly understood and will certainly constitute an important part of future OMV research.
Abbreviations
- IL:
-
Interleukin
- LPS:
-
Lipopolysaccharide
- MHC:
-
Major histocompatibility complex
- OMV(s):
-
Outer membrane vesicle(s)
- TIII or TIVSS:
-
Type III or IV secretion system
- TNF-α:
-
Tumor necrosis factor-α
- COPD:
-
Chronic obstructive pulmonary disease
- MCP-1:
-
Macrophage chemoattractant protein-1
- IFN-γ:
-
Interferon-γ
References
Beveridge TJ (1999) Structures of Gram-negative cell walls and their derived membrane vesicles. J Bacteriol 181(16):4725–4733
Bishop DG, Work E (1965) An extracellular glycolipid produced by Escherichia coli grown under lysine-limiting conditions. Biochem J 96(2):567–576
Chatterjee SN, Das J (1967) Electron microscopic observations on the excretion of cell-wall material by Vibrio cholerae. J Gen Microbiol 49(1):1–11
Knox KW, Vesk M, Work E (1966) Relation between excreted lipopolysaccharide complexes and surface structures of a lysine-limited culture of Escherichia coli. J Bacteriol 92(4):1206–1217
Rothfield L, Pearlman-Kothencz M (1969) Synthesis and assembly of bacterial membrane components. A lipopolysaccharide–phospholipid–protein complex excreted by living bacteria. J Mol Biol 44(3):477–492
Work E, Knox KW, Vesk M (1966) The chemistry and electron microscopy of an extracellular lipopolysaccharide from Escherichia coli. Ann NY Acad Sci 133(2):438–449
Ellis TN, Kuehn MJ (2010) Virulence and immunomodulatory roles of bacterial outer membrane vesicles. Microbiol Mol Biol Rev 74(1):81–94. doi:10.1128/MMBR.00031-09
Mashburn-Warren L, McLean RJ, Whiteley M (2008) Gram-negative outer membrane vesicles: beyond the cell surface. Geobiology 6(3):214–219. doi:10.1111/j.1472-4669.2008.00157.x
Kulp A, Kuehn MJ (2010) Biological functions and biogenesis of secreted bacterial outer membrane vesicles. Annu Rev Microbiol 64:163–184. doi:10.1146/annurev.micro.091208.073413
Tetz VV, Rybalchenko OV, Savkova GA (1990) Ultrastructural features of microbial colony organization. J Basic Microbiol 30(8):597–607
Devoe IW, Gilchrist JE (1973) Release of endotoxin in the form of cell wall blebs during in vitro growth of Neisseria meningitidis. J Exp Med 138(5):1156–1167
Hozbor D, Rodriguez ME, Fernandez J, Lagares A, Guiso N, Yantorno O (1999) Release of outer membrane vesicles from Bordetella pertussis. Curr Microbiol 38(5):273–278
Shoberg RJ, Thomas DD (1995) Borrelia burgdorferi vesicle production occurs via a mechanism independent of immunoglobulin M involvement. Infect Immun 63(12):4857–4861
Wai SN, Takade A, Amako K (1995) The release of outer membrane vesicles from the strains of enterotoxigenic Escherichia coli. Microbiol Immunol 39(7):451–456
Schooling SR, Beveridge TJ (2006) Membrane vesicles: an overlooked component of the matrices of biofilms. J Bacteriol 188(16):5945–5957. doi:10.1128/JB.00257-06
Yonezawa H, Osaki T, Kurata S, Fukuda M, Kawakami H, Ochiai K, Hanawa T, Kamiya S (2009) Outer membrane vesicles of Helicobacter pylori TK1402 are involved in biofilm formation. BMC Microbiol 9:197. doi:10.1186/1471-2180-9-197
Galka F, Wai SN, Kusch H, Engelmann S, Hecker M, Schmeck B, Hippenstiel S, Uhlin BE, Steinert M (2008) Proteomic characterization of the whole secretome of Legionella pneumophila and functional analysis of outer membrane vesicles. Infect Immun 76(5):1825–1836. doi:10.1128/IAI.01396-07
Brandtzaeg P, Bryn K, Kierulf P, Ovstebo R, Namork E, Aase B, Jantzen E (1992) Meningococcal endotoxin in lethal septic shock plasma studied by gas chromatography, mass-spectrometry, ultracentrifugation, and electron microscopy. J Clin Invest 89(3):816–823. doi:10.1172/JCI115660
Hynes SO, Keenan JI, Ferris JA, Annuk H, Moran AP (2005) Lewis epitopes on outer membrane vesicles of relevance to Helicobacter pylori pathogenesis. Helicobacter 10(2):146–156. doi:10.1111/j.1523-5378.2005.00302.x
Stephens DS, Edwards KM, Morris F, McGee ZA (1982) Pili and outer membrane appendages on Neisseria meningitidis in the cerebrospinal fluid of an infant. J Infect Dis 146(4):568
Tan TT, Morgelin M, Forsgren A, Riesbeck K (2007) Haemophilus influenzae survival during complement-mediated attacks is promoted by Moraxella catarrhalis outer membrane vesicles. J Infect Dis 195(11):1661–1670. doi:10.1086/517611
Kadurugamuwa JL, Beveridge TJ (1995) Virulence factors are released from Pseudomonas aeruginosa in association with membrane vesicles during normal growth and exposure to gentamicin: a novel mechanism of enzyme secretion. J Bacteriol 177(14):3998–4008
Suzuki H, Nishimura Y, Yasuda S, Nishimura A, Yamada M, Hirota Y (1978) Murein-lipoprotein of Escherichia coli: a protein involved in the stabilization of bacterial cell envelope. Mol Gen Genet 167(1):1–9
Smalley JW, Birss AJ, McKee AS, Marsh PD (1991) Haemin-restriction influences haemin-binding, haemagglutination and protease activity of cells and extracellular membrane vesicles of Porphyromonas gingivalis W50. FEMS Microbiol Lett 69(1):63–67
Loeb MR (1974) Bacteriophage T4-mediated release of envelope components from Escherichia coli. J Virol 13(3):631–641
McBroom AJ, Kuehn MJ (2007) Release of outer membrane vesicles by Gram-negative bacteria is a novel envelope stress response. Mol Microbiol 63(2):545–558. doi:10.1111/j.1365-2958.2006.05522.x
Alaniz RC, Deatherage BL, Lara JC, Cookson BT (2007) Membrane vesicles are immunogenic facsimiles of Salmonella typhimurium that potently activate dendritic cells, prime B and T cell responses, and stimulate protective immunity in vivo. J Immunol 179(11):7692–7701
McBroom AJ, Johnson AP, Vemulapalli S, Kuehn MJ (2006) Outer membrane vesicle production by Escherichia coli is independent of membrane instability. J Bacteriol 188(15):5385–5392. doi:10.1128/JB.00498-06
Wensink J, Witholt B (1981) Outer-membrane vesicles released by normally growing Escherichia coli contain very little lipoprotein. Eur J Biochem 116(2):331–335
de Leij L, Kingma J, Witholt B (1979) Nature of the regions involved in the insertion of newly synthesized protein into the outer membrane of Escherichia coli. Biochim Biophys Acta 553(2):224–234
Zhou L, Srisatjaluk R, Justus DE, Doyle RJ (1998) On the origin of membrane vesicles in Gram-negative bacteria. FEMS Microbiol Lett 163(2):223–228
Nguyen TT, Saxena A, Beveridge TJ (2003) Effect of surface lipopolysaccharide on the nature of membrane vesicles liberated from the Gram-negative bacterium Pseudomonas aeruginosa. J Electron Microsc (Tokyo) 52(5):465–469
Sabra W, Lunsdorf H, Zeng AP (2003) Alterations in the formation of lipopolysaccharide and membrane vesicles on the surface of Pseudomonas aeruginosa PAO1 under oxygen stress conditions. Microbiology 149(Pt 10):2789–2795
Hirota Y, Suzuki H, Nishimura Y, Yasuda S (1977) On the process of cellular division in Escherichia coli: a mutant of E. coli lacking a murein-lipoprotein. Proc Natl Acad Sci USA 74(4):1417–1420
Hoekstra D, van der Laan JW, de Leij L, Witholt B (1976) Release of outer membrane fragments from normally growing Escherichia coli. Biochim Biophys Acta 455(3):889–899
Sonntag I, Schwarz H, Hirota Y, Henning U (1978) Cell envelope and shape of Escherichia coli: multiple mutants missing the outer membrane lipoprotein and other major outer membrane proteins. J Bacteriol 136(1):280–285
Iwami J, Murakami Y, Nagano K, Nakamura H, Yoshimura F (2007) Further evidence that major outer membrane proteins homologous to OmpA in Porphyromonas gingivalis stabilize bacterial cells. Oral Microbiol Immunol 22(5):356–360. doi:10.1111/j.1399-302X.2007.00363.x
Deatherage BL, Lara JC, Bergsbaken T, Rassoulian Barrett SL, Lara S, Cookson BT (2009) Biogenesis of bacterial membrane vesicles. Mol Microbiol 72(6):1395–1407. doi:10.1111/j.1365-2958.2009.06731.x
Namork E, Brandtzaeg P (2002) Fatal meningococcal septicaemia with “Blebbing” Meningococcus. Lancet 360(9347):1741. doi:10.1016/S0140-6736(02)11721-1
Vidakovics ML, Jendholm J, Morgelin M, Mansson A, Larsson C, Cardell LO, Riesbeck K (2010) B cell activation by outer membrane vesicles—a novel virulence mechanism. PLoS Pathog 6(1):e1000724. doi:10.1371/journal.ppat.1000724
Alvarez-Martinez CE, Christie PJ (2009) Biological diversity of prokaryotic type IV secretion systems. Microbiol Mol Biol Rev 73(4):775–808. doi:10.1128/MMBR.00023-09
Cornelis GR (2000) Type III secretion: a bacterial device for close combat with cells of their eukaryotic host. Philos Trans R Soc Lond B Biol Sci 355(1397):681–693. doi:10.1098/rstb.2000.0608
Kadurugamuwa JL, Beveridge TJ (1998) Delivery of the non-membrane-permeative antibiotic gentamicin into mammalian cells by using Shigella flexneri membrane vesicles. Antimicrob Agents Chemother 42(6):1476–1483
Olofsson A, Vallstrom A, Petzold K, Tegtmeyer N, Schleucher J, Carlsson S, Haas R, Backert S, Wai SN, Grobner G, Arnqvist A (2010) Biochemical and functional characterization of Helicobacter pylori vesicles. Mol Microbiol 77(6):1539–1555. doi:10.1111/j.1365-2958.2010.07307.x
Hoy B, Lower M, Weydig C, Carra G, Tegtmeyer N, Geppert T, Schroder P, Sewald N, Backert S, Schneider G, Wessler S (2010) Helicobacter pylori HtrA is a new secreted virulence factor that cleaves E-cadherin to disrupt intercellular adhesion. EMBO Rep 11(10):798–804. doi:10.1038/embor.2010.114
Deich RA, Hoyer LC (1982) Generation and release of DNA-binding vesicles by Haemophilus influenzae during induction and loss of competence. J Bacteriol 152(2):855–864
Dorward DW, Garon CF (1989) DNA-binding proteins in cells and membrane blebs of Neisseria gonorrhoeae. J Bacteriol 171(8):4196–4201
Dorward DW, Garon CF (1990) DNA is packaged within membrane-derived vesicles of Gram-negative but not Gram-positive bacteria. Appl Environ Microbiol 56(6):1960–1962
Renelli M, Matias V, Lo RY, Beveridge TJ (2004) DNA-containing membrane vesicles of Pseudomonas aeruginosa PAO1 and their genetic transformation potential. Microbiology 150(Pt 7):2161–2169. doi:10.1099/mic.0.26841-0
Dorward DW, Garon CF, Judd RC (1989) Export and intercellular transfer of DNA via membrane blebs of Neisseria gonorrhoeae. J Bacteriol 171(5):2499–2505
Beveridge TJ, Makin SA, Kadurugamuwa JL, Li Z (1997) Interactions between biofilms and the environment. FEMS Microbiol Rev 20(3–4):291–303
Olczak T, Wojtowicz H, Ciuraszkiewicz J, Olczak M (2010) Species specificity, surface exposure, protein expression, immunogenicity, and participation in biofilm formation of Porphyromonas gingivalis HmuY. BMC Microbiol 10:134. doi:10.1186/1471-2180-10-134
Kulp A, Kuehn MJ (2010) Biological functions and biogenesis of secreted bacterial outer membrane vesicles. Annu Rev Microbiol 64:163–184. doi:10.1146/annurev.micro.091208.073413
MacDonald KL, Beveridge TJ (2002) Bactericidal effect of gentamicin-induced membrane vesicles derived from Pseudomonas aeruginosa PAO1 on Gram-positive bacteria. Can J Microbiol 48(9):810–820
Bomberger JM, Maceachran DP, Coutermarsh BA, Ye S, O’Toole GA, Stanton BA (2009) Long-distance delivery of bacterial virulence factors by Pseudomonas aeruginosa outer membrane vesicles. PLoS Pathog 5(4):e1000382. doi:10.1371/journal.ppat.1000382
Parker H, Chitcholtan K, Hampton MB, Keenan JI (2010) Uptake of Helicobacter pylori outer membrane vesicles by gastric epithelial cells. Infect Immun 78(12):5054–5061
Pike LJ (2003) Lipid rafts: bringing order to chaos. J Lipid Res 44(4):655–667. doi:10.1194/jlr.R200021-JLR200
Schaar V, De Vries SP, Vidakovics ML, Bootsma HJ, Larsson L, Hermans PW, Bjartell A, Morgelin M, Riesbeck K (2010) Multicomponent Moraxella catarrhalis outer membrane vesicles induce an inflammatory response and are internalized by human epithelial cells. Cell Microbiol. doi:10.1111/j.1462-5822.2010.01546.x
Slevogt H, Zabel S, Opitz B, Hocke A, Eitel J, N’Guessan PD, Lucka L, Riesbeck K, Zimmermann W, Zweigner J, Temmesfeld-Wollbrueck B, Suttorp N, Singer BB (2008) CEACAM1 inhibits toll-like receptor 2-triggered antibacterial responses of human pulmonary epithelial cells. Nat Immunol 9(11):1270–1278. doi:10.1038/ni.1661
Fernandez-Moreira E, Helbig JH, Swanson MS (2006) Membrane vesicles shed by Legionella pneumophila inhibit fusion of phagosomes with lysosomes. Infect Immun 74(6):3285–3295. doi:10.1128/IAI.01382-05
Bauman SJ, Kuehn MJ (2009) Pseudomonas aeruginosa vesicles associate with and are internalized by human lung epithelial cells. BMC Microbiol 9:26. doi:10.1186/1471-2180-9-26
Bishop AL, Schild S, Patimalla B, Klein B, Camilli A (2010) Mucosal immunization with Vibrio cholerae outer membrane vesicles provides maternal protection mediated by antilipopolysaccharide antibodies that inhibit bacterial motility. Infect Immun 78(10):4402–4420. doi:10.1128/IAI.00398-10
Kim YR, Kim BU, Kim SY, Kim CM, Na HS, Koh JT, Choy HE, Rhee JH, Lee SE (2010) Outer membrane vesicles of Vibrio vulnificu deliver cytolysin-hemolysin VvhA into epithelial cells to induce cytotoxicity. Biochem Biophys Res Commun 399(4):607–612. doi:10.1016/j.bbrc.2010.07.122
Shoberg RJ, Thomas DD (1993) Specific adherence of Borrelia burgdorferi extracellular vesicles to human endothelial cells in culture. Infect Immun 61(9):3892–3900
Kaparakis M, Turnbull L, Carneiro L, Firth S, Coleman HA, Parkington HC, Le Bourhis L, Karrar A, Viala J, Mak J, Hutton ML, Davies JK, Crack PJ, Hertzog PJ, Philpott DJ, Girardin SE, Whitchurch CB, Ferrero RL (2010) Bacterial membrane vesicles deliver peptidoglycan to NOD1 in epithelial cells. Cell Microbiol 12(3):372–385. doi:10.1111/j.1462-5822.2009.01404.x
Kesty NC, Mason KM, Reedy M, Miller SE, Kuehn MJ (2004) Enterotoxigenic Escherichia coli vesicles target toxin delivery into mammalian cells. EMBO J 23(23):4538–4549. doi:10.1038/sj.emboj.7600471
Furuta N, Takeuchi H, Amano A (2009) Entry of Porphyromonas gingivalis outer membrane vesicles into epithelial cells causes cellular functional impairment. Infect Immun 77(11):4761–4770. doi:10.1128/IAI.00841-09
Hallstrom T, Riesbeck K (2010) Haemophilus influenzae and the complement system. Trends Microbiol 18(6):258–265. doi:S0966-842X(10)00045-4
Singh B, Su YC, Riesbeck K (2010) Vitronectin in bacterial pathogenesis: a host protein used in complement escape and cellular invasion. Mol Microbiol 78(3):545–560. doi:10.1111/j.1365-2958.2010.07373.x
Blom AM, Hallstrom T, Riesbeck K (2009) Complement evasion strategies of pathogens-acquisition of inhibitors and beyond. Mol Immunol 46(14):2808–2817. doi:10.1016/j.molimm.2009.04.025
Zipfel PF, Skerka C (2009) Complement regulators and inhibitory proteins. Nat Rev Immunol 9(10):729–740. doi:10.1038/nri2620
Nordstrom T, Blom AM, Forsgren A, Riesbeck K (2004) The emerging pathogen Moraxella catarrhalis interacts with complement inhibitor C4b binding protein through ubiquitous surface proteins A1 and A2. J Immunol 173(7):4598–4606
Nordstrom T, Blom AM, Tan TT, Forsgren A, Riesbeck K (2005) Ionic binding of C3 to the human pathogen Moraxella catarrhalis is a unique mechanism for combating innate immunity. J Immunol 175(6):3628–3636
Bergman MA, Cummings LA, Barrett SL, Smith KD, Lara JC, Aderem A, Cookson BT (2005) CD4+ T cells and toll-like receptors recognize Salmonella antigens expressed in bacterial surface organelles. Infect Immun 73(3):1350–1356. doi:10.1128/IAI.73.3.1350-1356.2005
Appelmelk BJ, Simoons-Smit I, Negrini R, Moran AP, Aspinall GO, Forte JG, De Vries T, Quan H, Verboom T, Maaskant JJ, Ghiara P, Kuipers EJ, Bloemena E, Tadema TM, Townsend RR, Tyagarajan K, Crothers JM Jr, Monteiro MA, Savio A, De Graaff J (1996) Potential role of molecular mimicry between Helicobacter pylori lipopolysaccharide and host Lewis blood group antigens in autoimmunity. Infect Immun 64(6):2031–2040
Murphy TF, Parameswaran GI (2009) Moraxella catarrhalis, a human respiratory tract pathogen. Clin Infect Dis 49(1):124–131. doi:10.1086/599375
Palliyil S, Broadbent ID (2009) Novel immunotherapeutic approaches to the treatment of infections caused by Gram-negative bacteria. Curr Opin Pharmacol 9(5):566–570. doi:10.1016/j.coph.2009.07.007
Simon J, Kotloff K (2010) New and candidate vaccines for gastrointestinal infections. Curr Opin Gastroenterol 26(1):12–16. doi:10.1097/MOG.0b013e328333f8ee
Kaufmann SH (2007) The contribution of immunology to the rational design of novel antibacterial vaccines. Nat Rev Microbiol 5(7):491–504. doi:10.1038/nrmicro1688
Raghunath D (2008) Emerging antibiotic resistance in bacteria with special reference to India. J Biosci 33(4):593–603
Frasch CE, Peppler MS (1982) Protection against group B Neisseria meningitidis disease: preparation of soluble protein and protein-polysaccharide immunogens. Infect Immun 37(1):271–280
Karch H, Nixdorff K (1980) Comparison of quantitative and qualitative antibody-producing cell responses to lipopolysaccharide in cell walls of the bacterial form and in membranes of the protoplast L-form of Proteus mirabilis. Infect Immun 30(2):349–352
Karch H, Nixdorff K (1981) Antibody-producing cell responses to an isolated outer membrane protein and to complexes of this antigen with lipopolysaccharide or with vesicles of phospholipids from Proteus mirabilis. Infect Immun 31(3):862–867
Wang LY, Frasch CE (1984) Development of a Neisseria meningitidis group B serotype 2b protein vaccine and evaluation in a mouse model. Infect Immun 46(2):408–414
Zollinger WD, Mandrell RE, Altieri P, Berman S, Lowenthal J, Artenstein MS (1978) Safety and immunogenicity of a Neisseria meningitidis type 2 protein vaccine in animals and humans. J Infect Dis 137(6):728–739
Girard MP, Preziosi MP, Aguado MT, Kieny MP (2006) A review of vaccine research and development: meningococcal disease. Vaccine 24(22):4692–4700. doi:10.1016/j.vaccine.2006.03.034
Sadarangani M, Pollard AJ (2010) Serogroup B meningococcal vaccines-an unfinished story. Lancet Infect Dis 10(2):112–124. doi:10.1016/S1473-3099(09)70324-X
Miller E, Salisbury D, Ramsay M (2001) Planning, registration, and implementation of an immunisation campaign against meningococcal serogroup C disease in the UK: a success story. Vaccine 20(Suppl 1):S58–S67
Ramsay ME, Andrews N, Kaczmarski EB, Miller E (2001) Efficacy of meningococcal serogroup C conjugate vaccine in teenagers and toddlers in England. Lancet 357(9251):195–196. doi:10.1016/S0140-6736(00)03594-7
Pichichero ME (2005) Meningococcal conjugate vaccine in adolescents and children. Clin Pediatr (Phila) 44(6):479–489
Pichichero M, Casey J, Blatter M, Rothstein E, Ryall R, Bybel M, Gilmet G, Papa T (2005) Comparative trial of the safety and immunogenicity of quadrivalent (A, C, Y, W-135) meningococcal polysaccharide-diphtheria conjugate vaccine versus quadrivalent polysaccharide vaccine in two- to ten-year-old children. Pediatr Infect Dis J 24(1):57–62
Shepard CW, Ortega-Sanchez IR, Scott RD 2nd, Rosenstein NE (2005) Cost-effectiveness of conjugate meningococcal vaccination strategies in the United States. Pediatrics 115(5):1220–1232. doi:10.1542/peds.2004-2514
Wyle FA, Artenstein MS, Brandt BL, Tramont EC, Kasper DL, Altieri PL, Berman SL, Lowenthal JP (1972) Immunologic response of man to group B meningococcal polysaccharide vaccines. J Infect Dis 126(5):514–521
Jennings HJ, Lugowski C (1981) Immunochemistry of groups A, B, and C meningococcal polysaccharide-tetanus toxoid conjugates. J Immunol 127(3):1011–1018
Finne J, Leinonen M, Makela PH (1983) Antigenic similarities between brain components and bacteria causing meningitis. Implications for vaccine development and pathogenesis. Lancet 2(8346):355–357
Bovre K, Holten E, Vik-Mo H, Brondbo A, Bratlid D, Bjark P, Moe PJ (1977) Neisseria meningitidis infections in northern Norway: an epidemic in 1974–1975 due mainly to group b organisms. J Infect Dis 135(4):669–672
Kelly C, Arnold R, Galloway Y, O’Hallahan J (2007) A prospective study of the effectiveness of the New Zealand meningococcal B vaccine. Am J Epidemiol 166(7):817–823. doi:10.1093/aje/kwm147
Sierra GV, Campa HC, Varcacel NM, Garcia IL, Izquierdo PL, Sotolongo PF, Casanueva GV, Rico CO, Rodriguez CR, Terry MH (1991) Vaccine against group B Neisseria meningitidis: protection trial and mass vaccination results in cuba. NIPH Ann 14(2):195–207, discussion 208–110
Martin DR, Walker SJ, Baker MG, Lennon DR (1998) New Zealand epidemic of meningococcal disease identified by a strain with phenotype B:4:P1.4. J Infect Dis 177(2):497–500
Holst J, Martin D, Arnold R, Huergo CC, Oster P, O’Hallahan J, Rosenqvist E (2009) Properties and clinical performance of vaccines containing outer membrane vesicles from Neisseria meningitidis. Vaccine 27(Suppl 2):B3–B12. doi:10.1016/j.vaccine.2009.04.071
van de Waterbeemd B, Streefland M, van der Ley P, Zomer B, van Dijken H, Martens D, Wijffels R, van der Pol L (2010) Improved OMV vaccine against Neisseria meningitidis using genetically engineered strains and a detergent-free purification process. Vaccine 28(30):4810–4816. doi:10.1016/j.vaccine.2010.04.082
Saunders NB, Brandt BL, Warren RL, Hansen BD, Zollinger WD (1998) Immunological and molecular characterization of three variant subtype P1.14 strains of Neisseria meningitidis. Infect Immun 66(7):3218–3222
Scholten RJ, Kuipers B, Valkenburg HA, Dankert J, Zollinger WD, Poolman JT (1994) Lipo-oligosaccharide immunotyping of Neisseria meningitidis by a whole-cell ELISA with monoclonal antibodies. J Med Microbiol 41(4):236–243
Urwin R, Russell JE, Thompson EA, Holmes EC, Feavers IM, Maiden MC (2004) Distribution of surface protein variants among hyperinvasive meningococci: implications for vaccine design. Infect Immun 72(10):5955–5962. doi:10.1128/IAI.72.10.5955-5962.2004
Tondella ML, Popovic T, Rosenstein NE, Lake DB, Carlone GM, Mayer LW, Perkins BA (2000) Distribution of Neisseria meningitidis serogroup B serosubtypes and serotypes circulating in the United States. The active bacterial core surveillance team. J Clin Microbiol 38(9):3323–3328
Findlow J, Borrow R, Snape MD, Dawson T, Holland A, John TM, Evans A, Telford KL, Ypma E, Toneatto D, Oster P, Miller E, Pollard AJ (2010) Multicenter, open-label, randomized phase II controlled trial of an investigational recombinant meningococcal serogroup B vaccine with and without outer membrane vesicles, administered in infancy. Clin Infect Dis 51(10):1127–1137. doi:10.1086/656741
Giuliani MM, Adu-Bobie J, Comanducci M, Arico B, Savino S, Santini L, Brunelli B, Bambini S, Biolchi A, Capecchi B, Cartocci E, Ciucchi L, Di Marcello F, Ferlicca F, Galli B, Luzzi E, Masignani V, Serruto D, Veggi D, Contorni M, Morandi M, Bartalesi A, Cinotti V, Mannucci D, Titta F, Ovidi E, Welsch JA, Granoff D, Rappuoli R, Pizza M (2006) A universal vaccine for serogroup B meningococcus. Proc Natl Acad Sci USA 103(29):10834–10839. doi:10.1073/pnas.0603940103
Keiser PB, Gibbs BT, Coster TS, Moran EE, Stoddard MB, Labrie JE 3rd, Schmiel DH, Pinto V, Chen P, Zollinger WD (2010) A phase 1 study of a group B meningococcal native outer membrane vesicle vaccine made from a strain with deleted lpxL2 and synX and stable expression of opcA. Vaccine 28(43):6970–6976. doi:10.1016/j.vaccine.2010.08.048
Zollinger WD, Donets MA, Schmiel DH, Pinto VB, Labrie JE 3rd, Moran EE, Brandt BL, Ionin B, Marques R, Wu M, Chen P, Stoddard MB, Keiser PB (2010) Design and evaluation in mice of a broadly protective meningococcal group B native outer membrane vesicle vaccine. Vaccine 28(31):5057–5067
Klugman KP, Gotschlich EC, Blake MS (1989) Sequence of the structural gene (rmpM) for the class 4 outer membrane protein of Neisseria meningitidis, homology of the protein to gonococcal protein III and Escherichia coli OmpA, and construction of meningococcal strains that lack class 4 protein. Infect Immun 57(7):2066–2071
Lee SR, Kim SH, Jeong KJ, Kim KS, Kim YH, Kim SJ, Kim E, Kim JW, Chang KT (2009) Multi-immunogenic outer membrane vesicles derived from an msbB-deficient Salmonella enterica serovar Typhimurium mutant. J Microbiol Biotechnol 19(10):1271–1279
Roy N, Barman S, Ghosh A, Pal A, Chakraborty K, Das SS, Saha DR, Yamasaki S, Koley H (2010) Immunogenicity and protective efficacy of Vibrio cholerae outer membrane vesicles in rabbit model. FEMS Immunol Med Microbiol. doi:10.1111/j.1574-695X.2010.00692.x
Schild S, Nelson EJ, Bishop AL, Camilli A (2009) Characterization of Vibrio cholerae outer membrane vesicles as a candidate vaccine for cholera. Infect Immun 77(1):472–484. doi:10.1128/IAI.01139-08
Schild S, Nelson EJ, Camilli A (2008) Immunization with Vibrio cholerae outer membrane vesicles induces protective immunity in mice. Infect Immun 76(10):4554–4563. doi:10.1128/IAI.00532-08
Shang ES, Champion CI, Wu XY, Skare JT, Blanco DR, Miller JN, Lovett MA (2000) Comparison of protection in rabbits against host-adapted and cultivated Borrelia burgdorferi following infection-derived immunity or immunization with outer membrane vesicles or outer surface protein A. Infect Immun 68(7):4189–4199
Roberts R, Moreno G, Bottero D, Gaillard ME, Fingermann M, Graieb A, Rumbo M, Hozbor D (2008) Outer membrane vesicles as acellular vaccine against pertussis. Vaccine 26(36):4639–4646. doi:10.1016/j.vaccine.2008.07.004
Blanco DR, Champion CI, Lewinski MA, Shang ES, Simkins SG, Miller JN, Lovett MA (1999) Immunization with Treponema pallidum outer membrane vesicles induces high-titer complement-dependent treponemicidal activity and aggregation of T. pallidum rare outer membrane proteins (TROMPS). J Immunol 163(5):2741–2746
Demuth DR, James D, Kowashi Y, Kato S (2003) Interaction of Actinobacillus actinomycetemcomitans outer membrane vesicles with HL60 cells does not require leukotoxin. Cell Microbiol 5(2):111–121
Kato S, Kowashi Y, Demuth DR (2002) Outer membrane-like vesicles secreted by Actinobacillus actinomycetemcomitans are enriched in leukotoxin. Microb Pathog 32(1):1–13. doi:10.1006/mpat.2001.0474
Hanson MS, Cassatt DR, Guo BP, Patel NK, McCarthy MP, Dorward DW, Hook M (1998) Active and passive immunity against Borrelia burgdorferi decorin binding protein A (DbpA) protects against infection. Infect Immun 66(5):2143–2153
Lindmark B, Rompikuntal PK, Vaitkevicius K, Song T, Mizunoe Y, Uhlin BE, Guerry P, Wai SN (2009) Outer membrane vesicle-mediated release of cytolethal distending toxin (CDT) from Campylobacter jejuni. BMC Microbiol 9:220. doi:10.1186/1471-2180-9-220
Wai SN, Lindmark B, Soderblom T, Takade A, Westermark M, Oscarsson J, Jass J, Richter-Dahlfors A, Mizunoe Y, Uhlin BE (2003) Vesicle-mediated export and assembly of pore-forming oligomers of the enterobacterial ClyA cytotoxin. Cell 115(1):25–35
Fiocca R, Necchi V, Sommi P, Ricci V, Telford J, Cover TL, Solcia E (1999) Release of Helicobacter pylori vacuolating cytotoxin by both a specific secretion pathway and budding of outer membrane vesicles. Uptake of released toxin and vesicles by gastric epithelium. J Pathol 188(2):220–226. doi:10.1002/(SICI)1096-9896(199906)188:2<220::AID-PATH307>3.0.CO;2-C
Kahnt J, Aguiluz K, Koch J, Treuner-Lange A, Konovalova A, Huntley S, Hoppert M, Sogaard-Andersen L, Hedderich R (2010) Profiling the outer membrane proteome during growth and development of the social bacterium Myxococcus xanthus by selective biotinylation and analyses of outer membrane vesicles. J Proteome Res 9(10):5197–5208. doi:10.1021/pr1004983
Ram S, Cullinane M, Blom AM, Gulati S, McQuillen DP, Boden R, Monks BG, O’Connell C, Elkins C, Pangburn MK, Dahlback B, Rice PA (2001) C4bp binding to porin mediates stable serum resistance of Neisseria gonorrhoeae. Int Immunopharmacol 1(3):423–432
Massari P, Gunawardana J, Liu X, Wetzler LM (2010) Meningococcal porin PorB prevents cellular apoptosis in a toll-like receptor 2- and NF-κB-independent manner. Infect Immun 78(3):994–1003. doi:10.1128/IAI.00156-09
Massari P, Ho Y, Wetzler LM (2000) Neisseria meningitidis porin PorB interacts with mitochondria and protects cells from apoptosis. Proc Natl Acad Sci USA 97(16):9070–9075
Orihuela CJ, Mahdavi J, Thornton J, Mann B, Wooldridge KG, Abouseada N, Oldfield NJ, Self T, Ala’Aldeen DA, Tuomanen EI (2009) Laminin receptor initiates bacterial contact with the blood brain barrier in experimental meningitis models. J Clin Invest 119(6):1638–1646. doi:10.1172/JCI36759
Sharma A, Novak EK, Sojar HT, Swank RT, Kuramitsu HK, Genco RJ (2000) Porphyromonas gingivalis platelet aggregation activity: outer membrane vesicles are potent activators of murine platelets. Oral Microbiol Immunol 15(6):393–396
Mashburn-Warren L, Howe J, Garidel P, Richter W, Steiniger F, Roessle M, Brandenburg K, Whiteley M (2008) Interaction of quorum signals with outer membrane lipids: insights into prokaryotic membrane vesicle formation. Mol Microbiol 69(2):491–502
Ciofu O, Beveridge TJ, Kadurugamuwa J, Walther-Rasmussen J, Hoiby N (2000) Chromosomal beta-lactamase is packaged into membrane vesicles and secreted from Pseudomonas aeruginosa. J Antimicrob Chemother 45(1):9–13
Ellis TN, Leiman SA, Kuehn MJ (2010) Naturally produced outer membrane vesicles from Pseudomonas aeruginosa elicit a potent innate immune response via combined sensing of both lipopolysaccharide and protein components. Infect Immun 78(9):3822–3831. doi:10.1128/IAI.00433-10
Mashburn LM, Whiteley M (2005) Membrane vesicles traffic signals and facilitate group activities in a prokaryote. Nature 437(7057):422–425. doi:10.1038/nature03925
Kitagawa R, Takaya A, Ohya M, Mizunoe Y, Takade A, Yoshida S, Isogai E, Yamamoto T (2010) Biogenesis of Salmonella enterica serovar Typhimurium membrane vesicles provoked by induction of pagc. J Bacteriol 192(21):5645–5656. doi:10.1128/JB.00590-10
Dutta S, Iida K, Takade A, Meno Y, Nair GB, Yoshida S (2004) Release of shiga toxin by membrane vesicles in Shigella dysenteriae serotype 1 strains and in vitro effects of antimicrobials on toxin production and release. Microbiol Immunol 48(12):965–969
Gorringe A, Halliwell D, Matheson M, Reddin K, Finney M, Hudson M (2005) The development of a meningococcal disease vaccine based on Neisseria lactamica outer membrane vesicles. Vaccine 23(17–18):2210–2213. doi:10.1016/j.vaccine.2005.01.055
Gorringe AR (2005) Can Neisseria lactamica antigens provide an effective vaccine to prevent meningococcal disease? Expert Rev Vaccines 4(3):373–379. doi:10.1586/14760584.4.3.373
Sardinas G, Yero D, Climent Y, Caballero E, Cobas K, Niebla O (2009) Neisseria meningitidis antigen NMB0088: sequence variability, protein topology and vaccine potential. J Med Microbiol 58(Pt 2):196–208. doi:10.1099/jmm.0.004820-0
Acknowledgments
This work was supported by grants from Alfred Österlund, the Anna and Edwin Berger, the Janne Elgqvist, the Marianne and Marcus Wallenberg, Krapperup, and the Greta and Johan Kock Foundations, the Swedish Medical Research Council, the Cancer Foundation at the University Hospital in Malmö, and Skane county council’s research and development foundation.
Author information
Authors and Affiliations
Corresponding author
Additional information
This article is published as part of the Special Issue on “Small vesicles as immune modulators.”
Rights and permissions
About this article
Cite this article
Ünal, C.M., Schaar, V. & Riesbeck, K. Bacterial outer membrane vesicles in disease and preventive medicine. Semin Immunopathol 33, 395–408 (2011). https://doi.org/10.1007/s00281-010-0231-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00281-010-0231-y