
Abstract
Theoretical and empirical evidence suggest that socio-biological factors determine the expression of behavioural lateralization across species. One would expect the same association at the intraspecific level, that is, that the differences in social strategies of the two sexes entail the sex differences in the lateralized social processing. This study aimed to test whether this hypothesis applies to the lateralized behaviour of offspring towards a mother. The preferences in the use of the lateral visual field of the left and right eye were assessed in wild Asian elephant, Elephas maximus mothers and their young sons and daughters. The spatial positioning relative to a social partner during approach was used as a behavioural indicator of visual lateralization. At the population level, elephant mothers preferred to keep the young in their left visual field during slow travelling. In contrast, young did not display a one-sided bias for the whole sample. The lateralization, however, was pronounced in a sex-specific manner—sons preferentially kept their mothers in the right visual field, while daughters preferred to keep mothers in the left visual field. Intriguingly, both sons and daughters preferentially kept the familiar older young in the left visual field. Sons, thus, showed oppositely directed lateral preferences towards mother and non-mother companion. Presumably, sons aim to approach the mother from her left side (rather than to keep her in the right visual field) and benefit from optimized maternal perception, while daughters facilitate their own perception of the mother by keeping her in the left visual field. These sex-related differences in lateralized behaviour may result from strikingly different social strategies of two sexes.
Significance statement
Young mammals show robust lateralization in the form of one-sided behavioural preferences in the interactions with their mother. Previous studies suggest that the social lifestyle may serve as a driving force in the evolution of behavioural lateralization. To test this proposition, we investigated behavioural lateralization in young subjects of Asian elephants, a species in which females are more gregarious than males. The lateralized behaviour of offspring towards the mother was found to be strongly sex-specific. In contrast, interactions with older young were lateralized in a similar manner in sons and daughters. Our results suggest that the benefits of a left-sided or right-sided position relative to mother have different significance for sons and daughters because of the distinctive social strategies of two sexes.
Similar content being viewed by others
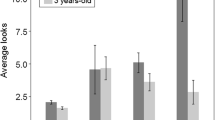


Avoid common mistakes on your manuscript.
Introduction
The relation between sociality and manifestation of lateralized responses is one of the main focuses of current research on behavioural lateralization (e.g. MacNeilage et al. 2009; Rogers et al. 2013; Chapelain et al. 2015; Boeving et al. 2017). It is argued that sociality is the driving force for the emergence of population-level biases in lateralized animal behaviour (Ghirlanda and Vallortigara 2004; Vallortigara and Rogers 2005; Ghirlanda et al. 2009). The uniformity in side preferences within a population is suggested to be relevant for social coordination, and mathematical modelling confirms that in a social context, behavioural lateralization at the population level can arise as an evolutionarily stable strategy (Ghirlanda and Vallortigara 2004; Ghirlanda et al. 2009). Side bias within the group seems to be advantageous, compared to an even distribution of left- and right-lateralized individuals, when asymmetrical subjects (i.e. subjects with a lateral preference for some behaviours) must coordinate their behaviour with that of conspecifics. This theoretical model was corroborated by experimental evidence demonstrating the benefits of social conformity in behavioural lateralization. Fish with a turning bias consistent with that of the majority of individuals in their group have improved escape performance (Chivers et al. 2016). Whether such benefits exist for other animals or other types of social behaviour remains unknown.
Lateralization of social behaviour has been shown in many vertebrate (e.g. Brancucci et al. 2009; Rosa Salva et al. 2012) and invertebrate species (e.g. Frasnelli 2013; Benelli et al. 2015). Yet, studies investigating coordination between behaviourally asymmetrical individuals, particularly conducted under natural circumstances in the wild, are scarce (e.g. Sakai et al. 2006). Interactions within mother–infant dyads, especially in species with long-lasting social bonds between the mother and her offspring, represent a good opportunity to fill this gap. Continuously maintaining physical proximity and engaging in synergistic activities, mothers and offspring may serve as a model to observe social coordination between asymmetrical individuals in nature. Mother-infant interactions have been recently found to be strongly lateralized in mammals. A consistent pattern of lateralization indicating left eye–right hemisphere advantage was revealed in 11 species of marine and terrestrial mammals (Karenina et al. 2017). This pattern demonstrated for members of the Cetartiodactyla, Perissodactyla (placentals) and Diprotodontia (marsupials) is in line with lateral biases in the behaviour of mothers and infants in human and non-human primates (e.g. Manning et al. 1994; Harris 2010; Forrester et al. 2014; Todd and Banerjee 2016) suggesting phylogenetic continuity of mother–offspring lateralization. However, a detailed study on a large-brained, highly social non-primate species with close, complex and long-lasting relations between female and her offspring comparable with those of primates is still lacking.
Socio-biological factors have been assumed to determine the expression of behavioural lateralization (MacNeilage et al. 2009). In line with this hypothesis, a consistent turning bias within a population was found in all studied gregarious species of fish that relied on schooling as a defence, while in more solitary species, population lateralization occurs significantly less likely (Bisazza et al. 2000). If sociality correlates with lateralized behaviour in various species, one would expect the same association at the intraspecific level when there are differences in sociality, e.g. different levels of sociality in males and females. Therefore, we aimed to test whether the differences in social strategies of the two sexes are reflected in sex differences in lateralized social processing, in particular, in the offspring’s perception of a mother. The natural history and socio-biology of elephants allow us to explore this supposition. When reaching adulthood, female elephants remain with their mother and become a member of the female social unit, while males disperse and have a more solitary lifestyle (Sukumar 2003; de Silva et al. 2011; Lee and Moss 2011). Although the differences in the degree of sociality between males and females become obvious in adult elephants, the behaviour of immature individuals differs significantly in two sexes. The social interactions of male and female young follow distinct behavioural patterns from an early age. For example, female young are involved both in more friendly and more aggressive interactions than are male young. The nearest non-mother companion of a male young is more likely to be a novel non-family male than is the case for female young with novel peer females. This may result in a greater risk of becoming separated from the family group for male young (Lee and Moss 2011). When interacting with the mother, sons attempt to suckle more often than daughters and are also more successful at their attempts. In addition, at older ages, mothers are more tolerant of their daughters’ demands to suckle (Lee and Moss 1986). Based on these findings, we hypothesized that lateralized interactions with the mother would differ between sons and daughters in elephants as opposed to previously studied species showing a uniform pattern of lateralization.
In wild Asian elephants, Elephas maximus, we assessed lateral biases in the behaviour of female and male young, as well as of their mothers. Preferences in the use of the lateral visual field of the left and right eye, serving as a behavioural marker of brain lateralization (reviewed in Rogers 2017), were examined. The spatial positioning relative to a social partner was recorded to infer lateralization in social processing which has been suggested as an informative method in previous studies (e.g. Baraud et al. 2009; Rosa Salva et al. 2012; Karenina et al. 2013; Forrester et al. 2014; Quaresmini et al. 2014). Characteristics of elephants’ visual system make them suitable for investigations of visual lateralization. Elephants have laterally set eyes and relatively little binocular overlap comparable to that in the horse, Equus ferus caballus (Suedmeyer 2006; Fowler and Mikota 2008), which is a widely used model for the studies on lateralized eye use (e.g. Austin and Rogers 2012). In such species, the visual fields of the left and the right eye are mostly independent, the majority of optic fibres decussate, and, consequently, a stimulus seen with the left eye is primarily processed by the right hemisphere, and vice versa (Vallortigara et al. 1999). Thus, the preference to keep a social partner on a particular side reflects animals’ visual lateralization (i.e. the preferential use of one visual field) (e.g. Baraud et al. 2009; Nagy et al. 2010; Austin and Rogers 2012; Quaresmini et al. 2014). Lateralization in elephants was assessed in (a) young’ approaches to mother, (b) young’ approaches to non-mother companion (juvenile/subadult group member) and (c) mothers’ approaches to young.
Methods
Study site and subjects
Observations on the behaviour of wild Asian elephants, Elephas maximus, were carried out in Uda Walawe National Park (6°27′41.1″N 80°54′10.0″E), Sri Lanka during the dry season in August–September 2016. The park is spread over 308 sq. km and encloses a large area of natural and planted forest, bush vegetation and open grasslands. The total population of Uda Walawe elephants is estimated to be about a thousand individuals (de Silva et al. 2011).
During the study period, a total of 44 elephant female–young pairs were individually identified and monitored in the study area, although the sample size varied according to the type of behaviour studied (see “Results”). It was not possible to ascertain whether the female was the biological mother of the young observed. We termed ‘mother’ an adult female group member continuously present in close proximity to a certain young elephant, and which was observed both to directly approach the young and to be directly approached by the young at least once. In 63% of these pairs, suckling was observed. Each infant was assigned to only one ‘mother’. In one case, two female adults fit our definition of a ‘mother’ and this case was therefore excluded from the sample. In addition, in 32 individually identified youngsters, we observed approaches to older young (5–10-year-old juveniles/subadults). Such non-mother companions of the young were observed travelling with the groups and displaying the same type of activities as the other group members did. Therefore, in all probability, the non-mother companions were familiar to the young. For each infant, we recorded approach behaviour to only one non-mother companion.
Each individual was photographed for photo-identification, which was based primarily on the morphology of the ears, tail, back and other body characteristics (de Silva et al. 2011; Vidya et al. 2014). Young elephants were sexed visually and classified into three broad age classes: calves (0–1 year), young juveniles (1–2 years) and older juveniles (2–5 years) based on their relative height and morphological characters (Arivazhagan and Sukumar 2008; de Silva et al. 2011; Varma et al. 2012). The number of individuals in each age and sex class is presented in Tables 1 and 2.
Data collection
Data collection was conducted from a research vehicle. We drove along the park roads in the early morning and/or in the afternoon (depending on weather conditions), which are the best periods to observe active elephant behaviours in open areas of the park. Tourist vehicles are common on the park roads and elephants are used to the presence of cars. Observations with binoculars and video-recording of elephants’ behaviour were conducted for 5–8 h per day. Observations of elephants were conducted opportunistically and we followed groups for as long as they were visible (Giljov et al. 2017).
The procedure of data collection on lateral preferences in mothers and young is described in Karenina et al. (2017). Using a continuous focal animal sampling method (Altmann 1974), we recorded the spontaneous choice of lateral position by pair members in mother–young pairs and non-mother companion–young pairs during slow travelling. The single choice of lateral position by a young was defined as follows: after a pair members’ spatial separation, the young elephant approached the (non-) mother from behind at a distance of less than one adult animal length, then positioned itself on one side of the (non-) mother. Thus, the approach resulted in young travelling side-by-side relative to the companion (Fig. 1a, b). Only the approaches resulted in side-by-side travelling, which lasted more than 30 s, were included in the analysis. Approaches from the front were rare in travelling elephants and were discarded from the analysis. A mother’s lateral position choice was registered using the same inclusion criteria applied for the position choice of the young. Additionally, in mother–young pairs, we recorded the young’ approaches to the mother for suckling. The young approached the mother from behind, stopped on one side of her and then turned its head and body to the mother’s mammary gland for suckling (Fig. 1c).

Lateral positioning of elephant offspring: a travelling near mother; b travelling near subadult group member (to the left: adult female; to the right: a young–subadult pair); c suckling
Discrete responses were obtained as follows: after a single choice of lateral position was registered, a subsequent choice was taken into account only after the pair had been separated by a distance of more than one adult animal length and the young again approached the mother from behind (or when the mother approached the young from behind). If any feature of the landscape or other animals seemed to prevent the pair member from choosing one of the lateral positions near the other pair member, such an event was discarded from the analysis. If the mother turned her head or directed any other social response to the young (e.g. trunk touch) just before or during the young’s position choice (i.e. before it took a lateral position), the event was excluded from the further analysis. The number of approaches obtained from each young is presented in Table 1.
Statistical analysis
We investigated preferences in lateral position choice in youngsters (separately for slow travelling and suckling) and mothers. Only the individuals with at least 10 choice events were assessed for individual preferences (Versace et al. 2007; Karenina et al. 2013). The number of times an individual chose to keep the other pair member on the left side or on the right side was compared using binomial tests. As a result, each individual was classified as having no preference or a preference for keeping the other pair member on the left/right side. To explore whether the distribution of left-lateralized, right-lateralized and non-lateralized individuals differed significantly from chance, i.e. 25%L:25%R:50%N (Güven et al. 2003; Scheumann et al. 2011), Chi-square tests were performed.
The young and mothers, for whom individual preferences were assessed, were included in population-level analyses using a laterality index (LI). The index was calculated for each individual using the following formula: LI = (L − R)/(L + R), where L and R are the number of times the individual chose to keep the other pair member on the left or right side, respectively. LI scores range on a continuum from − 1.0 to + 1.0, with negative values indicating the right side bias and positive values indicating the left side bias. We also calculated the absolute value of each subject’s LI to assess the strength of individual preference regardless of right or left direction (Abs-LI). Distributions of LI and Abs-LI scores were tested for normality using a Shapiro-Wilk test. If normality was rejected, non-parametric tests were employed. Lateralization at the population level was examined using a one-sample Wilcoxon signed rank test (or one sample t test). The Mann–Whitney U test on LI scores was used to explore the influence of young’ sex on lateralization. The Kruskal–Wallis test of independent samples (with post hoc Dunn’ s tests for between-pair comparisons) was carried out to estimate the effect of young’ age class on the expression of lateralization. In addition, a logistic regression model was used to analyse possible associations between the distribution of lateralized and non-lateralized individuals (dependent variable) and age and sex (independent variable). This analysis was performed using R software (http://www.R-project.org).
In order to increase the number of individuals included in the sample, the variety of studied behaviours, and enhance comparability of data across studies (e.g. Karenina et al. 2013, 2017; Forrester et al. 2014), we applied population-level lateralization testing that incorporates single observations per individual. The analysis based on single observations per individual does not allow testing of individual preferences, but is used to assess the population-level biases in lateralized behaviour (e.g. Seligmann 2002; Bourne and Todd 2004; Siniscalchi et al. 2012; Karenina et al. 2013; Forrester et al. 2014). A study of lateralized mother–infant interactions in a variety of mammal species showed that the results of population-level analyses based on multiple and single observations are consistent and show the same lateral bias (Karenina et al. 2017). Thus, analysis based on single observations appears to be an adequate method to estimate population-level lateral biases (Coren 1993). This type of analysis allowed us to assess lateralization in the behaviours that were rarely observed and the analysis based on multiple observations was not possible. The first observation of lateral position choice from each individual was included in the analysis. The number of left side position choices and the number of right side position choices were compared using a binomial test. In the case of lateral position choice in young approaching non-mother companions, the population-level analysis was the only method used.
All tests were two-tailed and the level of significance was set at 0.05.
Data availability
All data generated or analysed during this study are included in this published article and its supplementary information files.
Results
Population-level biases in mothers
The testing based on single observations per individual showed that elephant mothers displayed a population-level preference to keep their young on the left side during slow travelling (23 out of 25 mothers; binomial test: z = 4.00, P < 0.001; Table 2), irrespective of offspring sex (z test for proportions: z = − 1.52, P = 0.127). When approaching both male and female offspring, most mothers kept their young on the left side (daughters: binomial test: z = 2.41, P = 0.013; sons: z = 3.02, P < 0.001). On a smaller sample of mothers (N = 7) for which ≥ 10 approaches to their young were observed, the analysis of individual LI scores was also conducted and a population-level preference to keep their young on the left side was confirmed (mean LI ± SEM = 0.77 ± 0.09; one-sample t test: t6 = 8.88, P < 0.001).
Distribution of individual preferences in young
Based on multiple observations, individual preferences were assessed in young elephants from 26 mother–young pairs during slow travelling (Table 1). Binomial z scores indicated the following distribution of individual preferences: ten young kept their mothers predominantly on the left side, 10 preferred to keep their mothers on the right side and six individuals showed no preference. This distribution differed significantly from chance (χ22 = 7.54, P = 0.023). The significant majority of individuals were lateralized (binomial test: z = 2.55, P = 0.009). Twenty out of 26 animals (77%) were lateralized. Nine out of 10 lateralized sons (90%) preferred to keep their mothers on the right side, whereas nine out of 10 of lateralized daughters (90%) preferentially kept their mothers on the left side. A logistic regression model failed to reveal any significant interaction of the distribution of lateralized and non-lateralized individuals with age (P > 0.995) and sex (P > 0.124). However, the likelihood ratio test of a given model versus null model revealed a marginally significant joint influence of the factors (P = 0.048; additional data are given in Online Resource). Given a small sample size within the sex/age subgroups, further research on a larger sample is needed before a conclusion can be drawn regarding the effect of sex and age on the distribution of lateralized and non-lateralized individuals.
Population-level biases in young
Population-level lateralization of position choice by a young was assessed based on individual LI scores of 26 slowly travelling mother–young pairs. The direction of lateralization was significantly related to the sex of the young elephants (Mann–Whitney U test: U = 6.0, N1 = 14, N2 = 12, P < 0.001; Fig. 2). Significant group-level preference to keep the mother on the left side was found for daughters (median LI [95% confidence intervals] = 0.57 [− 0.13, 0.76]; one-sample Wilcoxon signed–rank test: Z = 85, P = 0.005, N = 14). Conversely, in sons, we found a significant group-level preference to keep the mother on the right side (median LI [95% CI] = − 0.94 [− 1.00, − 0.76]; Z = − 74, P = 0.001, N = 12). The strength of preferences was also significantly influenced by the young elephants’ sex, with sons showing stronger lateralization than daughters (Abs-LI were analysed; Mann–Whitney U test: U = 24.5, N1 = 14, N2 = 12, P = 0.001). There was no significant effect of youngsters’ age class on neither the direction (Kruskal–Wallis test: χ22 = 0.58, P = 0.750) or strength (χ22 = 1.82, P = 0.403) of lateralization in spatial positioning near the mother.

Sex differences in the direction of lateralization in young approaching mothers (N = 26). Median LI scores (± interquartile range, boxes; and 95% CI, whiskers) are given with positive values indicating the leftward bias (keeping the mother on the left side), and negative values indicate the rightward bias (keeping the mother on the right side). *P < 0.01
Further analyses based on single observations per individual (N = 39, Table 2) were conducted for male and female young separately. The majority of female young kept the mother on the left side (17 out of 21; binomial test: z = 2.62, P = 0.007), while most male young kept the mother on the right side (16 out of 18; z = − 3.06, P = 0.001). In contrast, when youngsters (N = 25; Table 2) approached the mother for suckling, no preferred side was found either for females (z = − 0.27, P = 0.791) or for males (z = 0, P > 0.99). These findings demonstrate the consistency between the results of analyses based on multiple and single observations.
We further analysed the single observations of slow travelling and suckling obtained from the same individuals. No significant difference between the number of similarly (e.g. left–left) and oppositely (e.g. left–right) directed position choices in two types of behaviour were found (binomial test: z = 0.64, P = 0.523) suggesting that these two types of behaviour are not associated.
In addition, the lateralization in young approaching the non-mother companion (older young) was assessed based on single observations per individual during slow travelling (N = 39; Table 2). The majority of young elephants kept the non-mother companion on the left side in both females (binomial test: z = 2.58, P = 0.007) and males (z = 3.71, P < 0.001). No significant preference for a non-mother companion of a particular sex was found in female and male young (P > 0.999, Fisher’s exact test for 2 × 2 contingency table).
Discussion
One-sided biases in mother–infant spatial relations have been considered as a distinctive feature of humans (Harris 2010), even though it has been found also in great apes (Manning et al. 1994). In this paper, we show the lateralization in interactions of mother and young in a non-primate species which, like humans and great apes, is a large-brained long-lived mammal. Together with a recent study in a range of non-primate mammals (Karenina et al. 2017), our results on Asian elephants confirm that humans are only one of many species whose mother–infant relations are lateralized. However, there is a principal difference between the lateralized mother–offspring positioning investigated in humans and elephants. In humans, mothers determine the position of an infant when cradling. The one-sided bias, therefore, is supposed to reflect the maternal rather than infant’s preference, although the potential influence of infant’s behaviour on mother’s cradling-side cannot be excluded (Scola et al. 2013). Asian elephant young are significantly more spatially independent from their mothers, and it was possible to assess the preferences of both members of the dyad separately. Additionally, spatial independence of the elephant young facilitates the clear assessment of lateralized perception in mother–offspring interactions, whereas in primates, motor biases (e.g. forelimb preferences) may potentially impact on the mother–infant positioning (Hopkins 2004). In elephant mothers and young, we estimated the lateralized choice of position relative to each other. Within the context of the current literature (e.g. Rosa Salva et al. 2012; Quaresmini et al. 2014; Forrester et al. 2014), one-sided spatial preferences in the social environment arise from the lateralized perception of conspecifics and its underlying hemispheric specialization for the processing of social information. When choosing the relative position during the pair reunion, elephant mothers and young chose a lateral visual field in which to keep the other pair member. Thus, the revealed spatial biases may reflect the preferences in the use of visual fields and its corresponding brain lateralization.
Human maternal behaviour is thought to be predominantly mediated by the right hemisphere (e.g. Lorberbaum et al. 2002; Bourne and Todd 2004; Huggenberger et al. 2009; Minagawa-Kawai et al. 2009). Likely, the same is the case for gorillas and chimpanzees showing, like humans, the left-cradling bias (Manning et al. 1994). Asian elephant mothers preferred to keep their young on the left side, i.e. position the young in their left visual field. This result points to a dominance of the right hemisphere in females’ perception of young. Evidence for the left eye/right hemisphere advantage for maternal monitoring of the offspring, similar to that in elephants, has also been found in other non-primate mammals such as feral horses, E. f. caballus and wild eastern grey kangaroos, Macropus giganteus (Karenina et al. 2017). In these species, however, the preference for keeping offspring in the left visual field emerged in females only in potentially threatening situations, such as when fleeing away from the source of disturbance with signs of anxiety, when the control of youngster’s state is most critical. In slow travelling, horse and kangaroo mothers showed no lateral preferences choosing the position near their offspring. In elephants, in contrast, mothers showed lateralization in routine behaviour during slow travelling with no behavioural signs of high activity, arousal or anxiety. The differences with other non-primate mammals studied may be associated with the extensive maternal care in elephants (Lee and Moss 1986; Sukumar 2003). A convincing body of evidence indicates that the left eye–right hemisphere system has a relative advantage in the implementation of many social functions (Brancucci et al. 2009; Nagy et al. 2010; Rosa Salva et al. 2012; Lindell 2013). Being especially attentive mothers, elephant females may aim to gain optimal monitoring of their young by the use of the left eye/right hemisphere even under usual non-threatening circumstances. Yet, overall, lateralization in elephant mothers is consistent with previous findings on lateralized maternal behaviour. The consistency between phylogenetically diverse mammals (Proboscidea, Primates, Perissodactyla, Diprotodontia) suggests that lateralization in general, and right-hemispheric superiority in particular, is one of the fundamental characteristic of maternal behaviour in mammals.
Asian elephant young were lateralized when choosing a lateral position near the mother after approaching her from behind during slow travelling. To the best of our knowledge, human infants’ active choice of spatial position near the mother has not been studied yet. However, unobtrusive observations within a playground showed that children preferentially chose a navigational path, which allowed them to position the adult (the relatedness was unknown) in their left hemispace/visual field implicating the right hemisphere (Forrester et al. 2014). A meta-analytical approach showed this bias to be consistent with lateralization in infants of 11 non-primate mammal species when approaching their mothers (Karenina et al. 2017). In a variety of behaviours, the infants of marine and terrestrial mammals displayed a uniform preference to keep the mother in the left visual field. Interestingly, a different picture was obtained in the present study. While there was no one-sided bias for the overall sample, male young preferentially kept their mothers in the right visual field, and female young preferred to keep mothers in the left visual field. This pattern was consistent across different age classes of young animals. In addition, males’ visual preferences were stronger than those of females. Previous studies found no effect of sex on youngsters’ visual lateralization in response to social stimuli (a mother), in domestic sheep, Ovis aries (Versace et al. 2007), feral horses, grey and red kangaroo, M. rufus (Karenina et al. 2017). In addition, chicks, Gallus gallus domesticus also did not display sex differences in lateralization of social responses (Deng and Rogers 2002). Sex differences, however, have been reported for lateralized processing of social information in adult vertebrates such as fish (e.g. Sovrano et al. 1999), humans (e.g. Reber and Tranel 2017) and non-human primates (e.g. Scheumann and Zimmermann 2008).
We suggest that the sex-specific lateralization in young elephants is not a result of differences in hemispheric lateralization of social functions in male and female young. Sex hormones exert significant influence on brain lateralization (Hausmann 2017; Reber and Tranel 2017) and presumably elicit differences in hemispheric dominance between males and females already at early developmental stages (Pfannkuche et al. 2009). If sex-specific lateralized responses toward the mother reflected general differences in hemispheric specialization between male and female young, these differences would likely be pronounced in other aspects of social behaviour. The observations of young’ approaches to non-mother companions demonstrate that this is not the case. Both male and female young preferentially kept the familiar older young in the left visual field. Thus, females display the same left eye preference irrespective of whether their social partner is the mother or not, while males show differential oppositely directed visual preferences toward mother and non-mother companion. More likely, sex-related differences in lateralized behaviour found here are associated with a different attitude to and relationship with the mother in elephant male and female young as a result of strikingly different social strategies of the two sexes (Lee and Moss 2011). In elephants, females stay with their natal family unit consisting of tightly associated females and their immature offspring, while males disperse on reaching adulthood and are characterized by relatively solitary lifestyle (Sukumar 2003; de Silva et al. 2011). Given these principal sex differences, male and female young may have distinctive emotional and/or motivational basis of relations with the mother and, consequently, display dissimilar lateralized behaviour towards her (discussed below).
Mother–offspring spatial positioning during travelling represents an example of social coordination between behaviourally asymmetrical individuals. The preferences found in the present study imply that mothers and daughters generally have a conflict from the point of view of spatial coordination since both pair members prefer to keep one another on the left when moving side-by-side. Consequently, in mother–daughter pairs, which of the two variants of relative lateral positioning of pair members will be taken during travelling is typically situation-specific and depends on who is choosing the lateral position, i.e. who is approaching whom. In mother–son pairs, in contrast, there is a low probability of a conflict in spatial positioning, since in both pair members, the preferential position choice leads to the same relative positioning (the young is on the left of the mother). Possibly, male young align their asymmetrical behaviour to the mother’s lateralization by taking the lateral position preferable for the mother. Previously the preference to approach a dominant group member by its left has been reported for grey-cheeked mangabeys, Lophocebus a. albigena (Baraud et al. 2009). Authors point out that such positioning may be beneficial for an approaching individual by improving the dominant individual’s perception and recognition. Similarly, in elephants, approaching the mother from her left the male young may benefit from mother’s better perception and/or faster recognition and, consequently, her more anticipated and appropriate reaction. Indeed, in humans, females with a left-cradling bias show a left visual field advantage for recognition of infant facial expressions (Huggenberger et al. 2009).
Thus, the potential benefits associated with optimized maternal perception are a plausible explanation of male young preference to position themselves so as to be in the mother’s left visual field. Why, then, female young prefer the opposite position relative to the mother? We suggest that for a female, it is more important to facilitate its own perception of the mother and social processing by keeping the mother in the left visual field. In elephants, female young have especially strong and prolonged affiliative relationships with the mother (Sukumar 2003; Lee and Moss 2011). The right brain hemisphere has been suggested to play a specialized role in social attachment (Lorberbaum et al. 2002; Nagasawa et al. 2013; Hecht 2014). In particular, when keeping a mother predominantly in the left visual field, feral horse foals direct more attachment behaviour to the mother than while perceiving her via the right eye–left hemisphere (Karenina et al. 2017). Moreover, the right hemisphere outperforms the left on social–emotional processing providing higher accuracy and speed for social responses (e.g. Calvo et al. 2015). In feral horses and Pacific walruses, Odobenus rosmarus divergens, the probability to be unintentionally left behind the mother is lower when the infant keep its mother in the left visual field. Thus, perceiving a mother via the left eye–right hemisphere system, infants maintain the spatial proximity to mother more successfully that is obviously beneficial for infant’s survival (Karenina et al. 2017). Given this hemispheric specialization and tight life-long bond with mother, in female young, the advantages gained from the use of the left eye–right hemisphere system for interactions with mother may outweigh the advantages of taking the opposite position which is preferable for the mother. To conclude, positioning to the left and to the right of the mother likely has its own benefits, but these benefits have differential significance for male and female elephant young because of their distinctive social strategies. In mosquitofish, Gambusia holbrooki, social females show a preferential use of the left eye during the inspection of their own mirror images, while solitary males did not exhibit any eye preferences during mirror-image inspection (Sovrano et al. 1999). Thus, the impact of different social strategies of the two sexes on the manifestation of lateralized social behaviour may be not restricted to the mammalian lineage.
An alternative explanation for sex differences in lateralized behaviour may be a bias to use the right or left hemisphere in daughters and sons, respectively. Human studies demonstrate that female subjects outperform males in a number of right hemisphere tasks (Herlitz et al. 1997; James and Kimura 1997; Rogers et al. 2013). If males use the abilities of the right hemisphere less effective than females, they may rely more on the use of the left hemisphere. In vertebrates, the left hemisphere predominantly controls a wide range of well-established patterns of behaviour performed in routine situations (Rogers 2010). The anterior regions of the left hemisphere control the approach system and reward-related responding (reviewed in Davidson 2004) which could play an important role in the male offsprings’ approaches to mother. This explanation, however, remains speculative, since the male infants of other mammal species besides elephants show left eye–right hemisphere advantage in their interactions with mother similarly to female infants (Karenina et al. 2017). In adult mammals also, males and females normally do not display oppositely directed visual preferences in social responses (e.g. Baraud et al. 2009; Austin and Rogers 2012; Boeving et al. 2017). It is more likely that sex-specific manifestation of social lateralization in elephant young which is more the exception than the rule is related to a distinctive feature of the species, i.e. different social strategies of males and females.
In contrast to slow travelling, no one-sided bias in elephant youngsters’ approaches to the mother for suckling was found neither in males nor in females. In feral horse and saiga antelope, Saiga t. tatarica, infants approaching mother for suckling displayed lateralized position choice similar to that in slow travelling and other behaviours (Karenina et al. 2017). The difference seen here with the mammals studied previously likely have a straightforward explanation. Horse and saiga offspring are able to reach the nipples on the left and right side of the centrally positioned udder without changing the lateral position relative to the mother. In contrast, Asian elephant young have to position themselves on the different sides of the mothers’ body to gain access to the left and to the right mammary glands located laterally in between the forelegs. Thus, to obtain milk from both glands, the young needs to alternate the sides of approaches for suckling.
In this study, we provide the first evidence for lateralized interactions between young and mother in an Afrotherian species. Previous studies suggested a pattern, consistent across diverse mammal species, with a preference to keep the other pair member on the left in both mother and young (Karenina et al. 2017). Although our results confirm this pattern for mothers, young’ behaviour toward the mother was found to be lateralized in a sex-specific manner. The findings suggest that in species with strikingly different social strategies of the two sexes, the lateralization of young-to-mother positioning may be determined by sex.
References
Altmann J (1974) Observational study of behavior: sampling methods. Behaviour 49(3):227–266. https://doi.org/10.1163/156853974X00534
Arivazhagan C, Sukumar R (2008) Constructing age structures of Asian elephant populations: a comparison of two field methods of age estimation. Gajah 29:11–16
Austin NP, Rogers LJ (2012) Limb preferences and lateralization of aggression, reactivity and vigilance in feral horses, Equus caballus. Anim Behav 83(1):239–247. https://doi.org/10.1016/j.anbehav.2011.10.033
Baraud I, Buytet B, Bec P, Blois-Heulin C (2009) Social laterality and ‘transversality’ in two species of mangabeys: influence of rank and implication for hemispheric specialization. Behav Brain Res 198(2):449–458. https://doi.org/10.1016/j.bbr.2008.11.032
Benelli G, Romano D, Messing RH, Canale A (2015) Population-level lateralized aggressive and courtship displays make better fighters not lovers: evidence from a fly. Behav Process 115:163–168. https://doi.org/10.1016/j.beproc.2015.04.005
Bisazza A, Cantalupo C, Capocchiano M, Vallortigara G (2000) Population lateralisation and social behaviour: a study with 16 species of fish. Laterality 5(3):269–284. https://doi.org/10.1080/713754381
Boeving ER, Belnap SC, Nelson EL (2017) Embraces are lateralized in spider monkeys (Ateles fusciceps rufiventris). Am J Primatol 79(6):e22654. https://doi.org/10.1002/ajp.22654
Bourne VJ, Todd BK (2004) When left means right: an explanation of the left cradling bias in terms of right hemisphere specializations. Dev Sci 7(1):19–24. https://doi.org/10.1111/j.1467-7687.2004.00318.x
Brancucci A, Lucci G, Mazzatenta A, Tommasi L (2009) Asymmetries of the human social brain in the visual, auditory and chemical modalities. Philos Trans R Soc Lond B Biol Sci 364(1519):895–914. https://doi.org/10.1098/rstb.2008.0279
Calvo MG, Rodríguez-Chinea S, Fernández-Martín A (2015) Lateralized discrimination of emotional scenes in peripheral vision. Exp Brain Res 233(3):997–1006. https://doi.org/10.1007/s00221-014-4174-8
Chapelain A, Pimbert P, Aube L, Perrocheau O, Debunne G, Bellido A, Blois-Heulin C (2015) Can population-level laterality stem from social pressures? Evidence from cheek kissing in humans. PLoS One 10(8):e0124477. https://doi.org/10.1371/journal.pone.0124477
Chivers DP, McCormick MI, Allan BJ, Mitchell MD, Gonçalves EJ, Bryshun R, Ferrari MC (2016) At odds with the group: changes in lateralization and escape performance reveal conformity and conflict in fish schools. Proc R Soc B 283(1841):20161127. https://doi.org/10.1098/rspb.2016.1127
Coren S (1993) Measurement of handedness via self-report: the relationship between brief and extended inventories. Percept Mot Skills 76(3):1035–1042. https://doi.org/10.2466/pms.1993.76.3.1035
Davidson RJ (2004) Well-being and affective style: neural substrates and biobehavioural correlates. Philos Trans R Soc Lond B Biol Sci 359(1449):1395–1411. https://doi.org/10.1098/rstb.2004.1510
de Silva S, Ranjeewa AD, Kryazhimskiy S (2011) The dynamics of social networks among female Asian elephants. BMC Ecol 11(1):17. https://doi.org/10.1186/1472-6785-11-17
de Silva S, Ranjeewa AD, Weerakoon D (2011) Demography of Asian elephants (Elephas maximus) at Uda Walawe National Park, Sri Lanka based on identified individuals. Biol Conserv 144(5):1742–1752. https://doi.org/10.1016/j.biocon.2011.03.011
Deng C, Rogers LJ (2002) Social recognition and approach in the chick: lateralization and effect of visual experience. Anim Behav 63(4):697–706. https://doi.org/10.1006/anbe.2001.1942
Forrester GS, Crawley M, Palmer C (2014) Social environment elicits lateralized navigational paths in two populations of typically developing children. Brain Cogn 91:21–27. https://doi.org/10.1016/j.bandc.2014.07.005
Fowler ME, Mikota SK (2008) Biology, medicine, and surgery of elephants. Blackwell, Oxford
Frasnelli E (2013) Brain and behavioral lateralization in invertebrates. Front Psychol 4:939
Ghirlanda S, Frasnelli E, Vallortigara G (2009) Intraspecific competition and coordination in the evolution of lateralization. Philos Trans R Soc Lond B Biol Sci 364(1519):861–866. https://doi.org/10.1098/rstb.2008.0227
Ghirlanda S, Vallortigara G (2004) The evolution of brain lateralization: a game-theoretical analysis of population structure. Proc R Soc Lond B 271(1541):853–858. https://doi.org/10.1098/rspb.2003.2669
Giljov A, Karenina K, de Silva S (2017) Context-dependent lateralization of trunk movements in wild Asian elephants. Biol Commun 62(2):82–92. https://doi.org/10.21638/11701/spbu03.2017.204
Güven M, Elalmis DD, Binokay S, Tan U (2003) Population-level right-paw preference in rats assessed by a new computerized food-reaching test. Int J Neurosci 113(12):1675–1689. https://doi.org/10.1080/00207450390249258
Harris LJ (2010) Side biases for holding and carrying infants: reports from the past and possible lessons for today. Laterality 15(1-2):56–135. https://doi.org/10.1080/13576500802584371
Hausmann M (2017) Why sex hormones matter for neuroscience: a very short review on sex, sex hormones, and functional brain asymmetries. J Neurosci Res 95(1-2):40–49. https://doi.org/10.1002/jnr.23857
Hecht D (2014) Cerebral lateralization of pro-and anti-social tendencies. Exp Neurobiol 23(1):1–27. https://doi.org/10.5607/en.2014.23.1.1
Herlitz A, Nilsson L-G, Backman L (1997) Gender differences in episodic memory. Mem Cognition 25(6):801–811. https://doi.org/10.3758/BF03211324
Hopkins WD (2004) Laterality in maternal cradling and infant positional biases: implications for the development and evolution of hand preferences in nonhuman primates. Int J Primatol 25(6):1243–1265. https://doi.org/10.1023/B:IJOP.0000043961.89133.3d
Huggenberger HJ, Suter SE, Reijnen E, Schachinger H (2009) Cradling side preference is associated with lateralized processing of baby facial expressions in females. Brain Cogn 70(1):67–72. https://doi.org/10.1016/j.bandc.2008.12.010
James TW, Kimura D (1997) Sex differences in remembering the locations of objects in an array: location-shift versus location-exchanges. Evol Hum Behav 18(3):155–163. https://doi.org/10.1016/S1090-5138(97)00004-4
Karenina K, Giljov A, Glazov D, Malashichev Y (2013) Social laterality in wild beluga whale infants: comparisons between locations, escort conditions, and ages. Behav Ecol Sociobiol 67(7):1195–1204. https://doi.org/10.1007/s00265-013-1545-2
Karenina K, Giljov A, Ingram J, Rowntree VJ, Malashichev Y (2017) Lateralization of mother–infant interactions in a diverse range of mammal species. Nat Ecol Evol 1(2):0030. https://doi.org/10.1038/s41559-016-0030
Karenina K, Giljov A, Ivkovich T, Burdin A, Malashichev Y (2013) Lateralization of spatial relationships between wild mother and infant orcas, Orcinus orca. Anim Behav 86(6):1225–1231. https://doi.org/10.1016/j.anbehav.2013.09.025
Lee PC, Moss CJ (1986) Early maternal investment in male and female African elephant calves. Behav Ecol Sociobiol 18(5):353–361. https://doi.org/10.1007/BF00299666
Lee PC, Moss CJ (2011) Calf development and maternal rearing strategies. In: Moss CJ, Croze H, Lee PC (eds) The Amboseli elephants: a long-term perspective on a long-lived mammal. University of Chicago Press, Chicago, pp 224–237. https://doi.org/10.7208/chicago/9780226542263.003.0014
Lindell AK (2013) Continuities in emotion lateralization in human and nonhuman primates. Front Hum Neurosci 7:464
Lorberbaum JP, Newman JD, Horwitz AR, Dubno JR, Lydiard RB, Hamner MB, Bohning DE, George MS (2002) A potential role for thalamocingulate circuitry in human maternal behavior. Biol Psychiatry 51(6):431–445. https://doi.org/10.1016/S0006-3223(01)01284-7
MacNeilage PF, Rogers LJ, Vallortigara G (2009) Origins of the left and right brain. Sci Am 301(1):60–67. https://doi.org/10.1038/scientificamerican0709-60
Manning JT, Heaton R, Chamberlain AT (1994) Left-side cradling: similarities and differences between apes and humans. J Hum Evol 26(1):77–83. https://doi.org/10.1006/jhev.1994.1005
Minagawa-Kawai Y, Matsuoka S, Dan I, Naoi N, Nakamura K, Kojima S (2009) Prefrontal activation associated with social attachment: facial-emotion recognition in mothers and infants. Cereb Cortex 19(2):284–292. https://doi.org/10.1093/cercor/bhn081
Nagasawa M, Kawai E, Mogi K, Kikusui T (2013) Dogs show left facial lateralization upon reunion with their owners. Behav Process 98:112–116. https://doi.org/10.1016/j.beproc.2013.05.012
Nagy M, Akos Z, Biro D, Vicsek T (2010) Hierarchical group dynamics in pigeon flocks. Nature 464(7290):890–893. https://doi.org/10.1038/nature08891
Pfannkuche KA, Bouma A, Groothuis TG (2009) Does testosterone affect lateralization of brain and behaviour? A meta-analysis in humans and other animal species. Philos Trans R Soc Lond B Biol Sci 364(1519):929–942. https://doi.org/10.1098/rstb.2008.0282
Quaresmini C, Forrester GS, Spiezio C, Vallortigara G (2014) Social environment elicits lateralized behaviors in gorillas and chimpanzees. J Comp Psychol 128(3):276–284. https://doi.org/10.1037/a0036355
Reber J, Tranel D (2017) Sex differences in the functional lateralization of emotion and decision making in the human brain. J Neurosci Res 95(1-2):270–278. https://doi.org/10.1002/jnr.23829
Rogers LJ (2010) Relevance of brain and behavioural lateralization to animal welfare. Appl Anim Behav Sci 127 (1–2):1–11. https://doi.org/10.1016/j.applanim.2010.06.008
Rogers LJ (2017) Eye and ear preferences. In: Rogers LJ, Vallortigara G (eds) Lateralized brain functions: methods in human and non-human species. Springer, New York, pp 79–102. https://doi.org/10.1007/978-1-4939-6725-4_3
Rogers LJ, Vallortigara G, Andrew RJ (2013) Divided brains: the biology and behavior of brain asymmetries. Cambridge University Press, New York. https://doi.org/10.1017/CBO9780511793899
Rosa Salva O, Regolin L, Mascalzoni E, Vallortigara G (2012) Cerebral and behavioural asymmetries in animal social recognition. Comp Cogn Behav Rev 7:110–138. https://doi.org/10.3819/ccbr.2012.70006
Sakai M, Hishii T, Takeda S, Kohshima S (2006) Laterality of flipper rubbing behaviour in wild bottlenose dolphins (Tursiops aduncus): caused by asymmetry of eye use? Behav Brain Res 170(2):204–210. https://doi.org/10.1016/j.bbr.2006.02.018
Scheumann M, Joly-Radko M, Leliveld L, Zimmermann E (2011) Does body posture influence hand preference in an ancestral primate model? BMC Evol Biol 11(1):52. https://doi.org/10.1186/1471-2148-11-52
Scheumann M, Zimmermann E (2008) Sex-specific asymmetries in communication sound perception are not related to hand preference in an early primate. BMC Biol 6(1):3. https://doi.org/10.1186/1741-7007-6-3
Scola C, Arciszewski T, Measelle J, Vauclair J (2013) Infant-holding bias variations in mother–child relationships: a longitudinal study. Eur J Dev Psychol 10(6):707–722. https://doi.org/10.1080/17405629.2013.791230
Seligmann H (2002) Behavioural and morphological asymmetries in hindlimbs of Hoplodactylus duvaucelii (Lacertilia: Gekkonomorpha: Gekkota: Diplodactylinae). Laterality 7(3):277–283. https://doi.org/10.1080/13576500244000067
Siniscalchi M, Dimatteo S, Pepe AM, Sasso R, Quaranta A (2012) Visual lateralization in wild striped dolphins (Stenella coeruleoalba) in response to stimuli with different degrees of familiarity. PLoS One 7(1):e30001. https://doi.org/10.1371/journal.pone.0030001
Sovrano VA, Rainoldi C, Bisazza A, Vallortigara G (1999) Roots of brain specializations: preferential left-eye use during mirror-image inspection in six species of teleost fish. Behav Brain Res 106(1-2):175–180. https://doi.org/10.1016/S0166-4328(99)00105-9
Suedmeyer WK (2006) Special senses. In: Fowler ME, Mikota SK (eds) Elephant biology, medicine, and surgery. Blackwell Publishing, Ames. https://doi.org/10.1002/9780470344484.ch31
Sukumar R (2003) The living elephants: evolutionary ecology, behaviour, and conservation. Oxford University Press, Oxford
Todd BK, Banerjee R (2016) Lateralization of infant holding by mothers: a longitudinal evaluation of variations over the first 12 weeks. Laterality 21(1):12–33. https://doi.org/10.1080/1357650X.2015.1059434
Vallortigara G, Rogers LJ (2005) Survival with an asymmetrical brain: advantages and disadvantages of cerebral lateralization. Behav Brain Sci 28:575–589
Vallortigara G, Rogers LJ, Bisazza A (1999) Possible evolutionary origins of cognitive brain lateralization. Brain Res Rev 30(2):164–175. https://doi.org/10.1016/S0165-0173(99)00012-0
Varma S, Baskaran N, Sukumar R (2012) Field key for elephant population estimation and age and sex classification. Asian Nature Conservation Foundation, Innovation Centre, Indian Institute of Science, Bangalore
Versace E, Morgante M, Pulina G, Vallortigara G (2007) Behavioural lateralization in sheep (Ovis aries). Behav Brain Res 184(1):72–80. https://doi.org/10.1016/j.bbr.2007.06.016
Vidya TNC, Prasad D, Ghosh A (2014) Individual identification in Asian elephants. Gajah 40:3–17
Acknowledgments
We are grateful to the field staff of the Uda Walawe Elephant Research Project and Uda Walawe National Park personnel. The authors thank the editor (Bernhard Voelkl) and anonymous reviewers for their helpful suggestions and comments on the manuscript.
Funding
The complex zoological expedition to Sri Lanka was funded by Saint Petersburg University (grant no. 1.42.1095.2016). The analytical work was conducted within a broader framework of the project funded by the Russian Science Foundation (no. 14-14-00284).
Author information
Authors and Affiliations
Contributions
KK and AG designed and organized the study, performed data collection and analyses and wrote the paper. SdS organized the field work and edited the paper. YM supervised the project and contributed to and edited the paper.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical approval
This study followed the ASAB/ABS (2012) guidelines for the use of animals in research and was approved by St. Petersburg State University ethical committee (permit no. 131-03-5).
Additional information
Communicated by B. Voelkl
Electronic supplementary material
ESM 1
Output of logistic regression model used to analyze possible associations of the distribution of lateralized and non-lateralized individuals (dependent variable) with age and sex (independent variable). (PDF 16 kb)
Rights and permissions
About this article

Cite this article
Karenina, K., Giljov, A., de Silva, S. et al. Social lateralization in wild Asian elephants: visual preferences of mothers and offspring. Behav Ecol Sociobiol 72, 21 (2018). https://doi.org/10.1007/s00265-018-2440-7
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00265-018-2440-7