
Abstract
While female preferences may vary depending on population-level differences in density or sex ratio, factors affecting mate choice may act at the individual level, i.e., females may encounter males with varying frequency or encounter multiple males simultaneously. The “socially cued anticipatory plasticity” hypothesis suggests that females may bias mate preferences based on prior experience. In the wolf spider, Schizocosa ocreata, males typically mature before females, allowing females to experience male courtship before maturation. Using video playback, we simulated differences in the encounter rate and the number of males simultaneously encountered to examine effects on female preference for a secondary sexual character (foreleg tufts). Penultimate females were exposed to video playback of zero, one, or three courting males either once every 2 days (low encounter rate) or twice per day (high encounter rate). At adulthood (week 2 post-maturity), females were presented video playback of courting males with small or large tufts to test for preferences in no-choice and two-choice designs. In two-choice (but not no-choice) presentations, female receptivity varied significantly with treatment. Females exposed to three males simultaneously at a higher encounter rate during their penultimate stage exhibited greater receptivity to large-tufted than small-tufted males as adults. Subsequent analyses revealed that females were more selective as adults if they encountered cumulatively more males during their penultimate stage, which was a repeatable trend when re-testing some individuals 3 weeks later. This study adds to the growing literature that demonstrates that invertebrates exhibit plasticity in mating preferences depending on social experience.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Across the animal kingdom, females often prefer males with brighter or larger secondary sexual characteristics (Andersson 1986; Kuijper et al. 2012). This selectivity is often argued to result in either indirect (Andersson 1994; Jennions and Petrie 2000) or direct (Kirkpatrick 1996; Iwasa and Pomiankowski 1999) fitness benefits for females choosing males that invested resources into increased trait expression. Female mating preferences may vary, however, based on intrinsic factors such as nutritional stress and condition (Hunt et al. 2005; Burley and Foster 2006; Byers et al. 2006; Hebets et al. 2008; Holveck and Riebel 2010), reproductive status (Lynch et al. 2005; Uetz and Norton 2007; Judge et al. 2010), and age (Gray 1999; Kodric-Brown and Nicoletto 2001; Coleman et al. 2004). Female mating preferences may also vary depending on extrinsic factors such as predation risks (Forsgren 1992; Hedrick and Dill 1993; Johnson and Basolo 2003), the presence of competitors (Fawcett and Johnstone 2003), social experience (Hebets 2003; Tudor and Morris 2009; Rutledge et al. 2010; Bailey 2011), and the physical environment (Gordon and Uetz 2011). Specifically, the effects of previous social experience on subsequent mate preferences, an example of “socially cued anticipatory plasticity” or SCAP (Kasumovic and Brooks 2011), have been increasingly studied over the last decade due to the impact of female preferences on the evolution of mating signals (Jennions and Petrie 1997; Kasumovic and Brooks 2011; Verzijden et al. 2012).
The density of potential mates and the operational sex ratio (OSR) are two demographic factors in an individual’s social environment that influence the overall availability of potential mates. Results from studies across a variety of taxa demonstrate variation in mating preferences based on the availability of potential mates. Females have been shown to be more selective (i.e., choose high-quality males vs low-quality mates) when male densities are high (e.g., Berglund 1995; Bonduriansky 2001; Kokko and Rankin 2006) or when OSRs are male-biased (e.g., Berglund 1994; Souroukis and Murray 1995; Baslhine-Earn 1996; Jirotkul 1999; Tinghitella et al. 2013). When male densities are low or when OSRs are female-biased, females are predicted to be less selective in order to avoid missing opportunities to mate. However, while OSR and male density are often quantified at the population level (e.g., Gwynne 1984; Palokangas et al. 1992), individual females presumably do not assess them at that level. Instead, individuals may have past experience with differences in the encounter rate (how frequently a female may encounter a male) and/or the number of males encountered simultaneously (e.g., encountering one or more males at a time), which is affected by the population density and OSR. SCAP in mating preferences based on the availability of potential mates would presumably be selected for over time, as long as social conditions of the past (e.g., high encounter rates or encounters with multiple males) accurately convey what the future social environment will be like. Consequently, SCAP would lead to behaviors (e.g., maintaining strict preferences for males with particular phenotypes or relaxing preferences) that are advantageous during mating decisions (Kasumovic and Brooks 2011). This study aims to determine whether SCAP in female selectivity is best predicted by previous experience with varying encounter rates and/or simultaneous encounters with multiple males.
Recent research across a variety of arthropods has demonstrated that invertebrates are not rigid in their mating preferences, making them useful study organisms to investigate SCAP (fruit flies: Dukas 2005; katydids: Fowler-Finn and Rodriguez 2012; crickets: Bailey and Zuk 2008; Bailey 2011; Kasumovic et al. 2012; Bailey and Macleod 2014; wolf spiders: Hebets 2003; Hebets and Vink 2007; Hebets 2007; Rutledge et al. 2010). In particular, Schizocosa ocreata is a useful organism to study the plasticity of female mate preferences for several reasons. Female S. ocreata have been shown to prefer males with large tufts (conspicuous dark tibial bristles on the forelegs) (McClintock and Uetz 1996; Scheffer et al. 1996) and can use this trait to choose high-quality males, because tuft size is a condition-dependent trait (Uetz et al. 2002). These previous studies, though, have been conducted on females without any prior experience with courting males, despite evidence from other studies in Schizocosa spp. demonstrating that previous experience affects mate preferences in some contexts (Hebets 2003; Hebets and Vink 2007; Rutledge et al. 2010). Studies have yet to investigate how variation in the availability of potential mates impact female mate preferences for tuft size. This is an interesting question to ask in S. ocreata, as variation in male densities across microhabitats exists and OSRs change as the mating season progresses (Roberts and Uetz, unpublished data).
During a period of asynchrony of maturation (males mature on average 2 weeks before females: Roberts and Uetz, unpublished data), female S. ocreata would be expected to encounter male courtship during their penultimate stage (i.e., the instar or growth stage before molting to maturity) for several reasons. Because males may court in response to chemical cues from penultimate females (though at a significantly lower level in comparison to male courtship levels in response to mature female silk), female S. ocreata may encounter male courtship prior to maturity (Roberts and Uetz 2005). There is also an overlap of juvenile and mature females, so males may court in response to silk from mature females while within the visual field of neighboring subadults. Additionally, males may court, though at a significantly reduced level, in response to no chemical cues at all (Roberts and Uetz 2005), providing additional opportunities for females to be exposed to male courtship. The ability to estimate the availability of potential mates during their penultimate stage (i.e., when they may be exposed to male courtship) would be particularly advantageous for species such as S. ocreata, in which females are monogamous and would not have the opportunity to re-mate (Norton and Uetz 2005).
In this study, we use video playback to give penultimate females experience with courting males at an individual level to examine the impact on female preferences of a wolf spider that exhibits asynchrony of maturation. We hypothesize that wolf spiders exhibit SCAP in mating preferences (Kasumovic and Brooks 2011) as a consequence of exposure to male courtship and that encounter frequency and/or simultaneous encounters with males experienced by penultimate females should influence female mate preferences as adults. Based on this hypothesis, we predict that when female S. ocreata either encounter males more frequently (twice per day) and/or encounter males simultaneously (three males per exposure) during their penultimate stage, they will increase their receptivity to large-tufted males relative to receptivity to small-tufted males. On the other hand, females that encounter males less frequently (every other day) or do not encounter males simultaneously (one male per exposure) will demonstrate relaxed preferences (i.e., equal receptivity to small-tufted and large-tufted males).
Using a subset of individuals, we also tested whether female preferences arising from social experience would be maintained at a later age (fifth week post-maturity). Previous studies have shown that in individuals without any previous social experience, age affects mate preference; i.e., at an age of 5 weeks post-maturity, female S. ocreata no longer maintain their earlier preferences for males with large tufts (Uetz and Norton 2007). However, if prior experience affects adult mate preferences as hypothesized above, we predict that female preferences will be repeatable and that relationships between experience during their penultimate stage and adult mate preferences will be maintained across multiple trials over time.
Methods
Animal care
Female S. ocreata were collected from the Cincinnati Nature Center, Clermont County, Ohio, USA, as virgin juveniles prior to the researchers encountering any mature males in the field. Each female was isolated in a plastic container with available water in a room with controlled humidity, temperature, and light cycle (L/D 11:13 h). All females were fed two to three crickets (10-day old) twice per week.
Overall design
In brief, we gave penultimate females repeated encounters (every other day or twice per day) with (zero, one, or three) visually courting males of average tuft size and tested female preferences for small-tufted and large-tufted males as adults during week 2 (see Fig. 1 for timeline). Week 2 trials were conducted in no-choice (days 1 and 2) and two-choice (day 3) experimental designs, and a subset of females was re-tested in two-choice trials during week 5 (days 4–7).

The experimental design, arenas, and sample sizes used in this study. The top row indicates the stage or age in which trials were conducted, while the bottom row depicts the arrangement of the iPod units in the clear plastic arenas (diameter 15.5 cm, height 6.5 cm) that were used in these trials. The first arena had three iPod units placed equilaterally around the edge and was used to provide females with experience during their penultimate stage. Once mature, females were presented a small-tufted or large-tufted male in separate no-choice trials on days 1–2 (one trial per day). On day 3 of week 2, females were simultaneously presented a small-tufted and large-tufted male in an arena with two iPods. The same two-choice arena was used again for a subset of individuals week 5 for three to four consecutive days to test the repeatability of female preferences
Video playback
Digital playback of courting male S. ocreata on LCD screens has been shown to elicit female receptivity behaviors (see review in Uetz and Roberts 2002). The use of video playback techniques allows precise control over interactions that an individual has with potential mates, while simultaneously providing a standardized stimulus that controls for individual variation in male phenotypes (in this study: body size, tuft size, and courtship vigor). These controlled variables are additionally important when measuring the repeatability of female preferences. Thus, we used video playback to vary the experience of penultimate females with males under lab-simulated conditions to determine whether adult females demonstrate plasticity in their receptivity according to male traits (foreleg tuft size). The courting male stimuli have been previously used in other studies (McClintock and Uetz 1996; Roberts et al. 2007; Moskalik and Uetz 2011). All courting male stimuli have a mean body size based on the population average from Roberts et al. (2007) and had identical courtship vigor (number of taps per minute). The courting male stimulus during the experience of penultimate females was a male with average tuft size (also based on population averages from Roberts et al. 2007). Digital stimuli of small-tufted and large-tufted males used to measure female selectivity during adulthood were previously manipulated to represent the lower and upper 95 % confidence intervals (see Roberts et al. 2007).
Manipulation of social experience
Exposure to male courtship began the day after females molted to their penultimate stage and ended as females matured. Each exposure lasted 5 min and took place in arenas with three iPod Classics placed equilaterally behind clear acetate sheets, facing the center of the arena. All iPods displayed identical leaf litter backgrounds during the exposures. However, we varied the number of iPods that displayed a video of a courting male with an average tuft size in the foreground (zero, one, or three) and the frequency of exposures (once every 2 days or twice per day). This resulted in six treatment groups: females (n = 12) exposed to zero males every other day, females (n = 8) exposed to zero males twice per day, females (n = 18) exposed to one male every other day, females (n = 18) exposed to one male twice per day, females (n = 22) exposed to three males every other day, and females (n = 18) exposed to three males twice per day. Originally, 130 penultimate females were randomly assigned to these six treatment groups. During the exposures, females escaped or developed parasites, resulting in reduced sample sizes. While there was natural variation in the length of a female’s penultimate instar (growth stage prior to maturity) during experience trials (i.e., females varied in how many days it took females to molt from their penultimate stage to their adult stage), there was not a significant difference in the penultimate instar length based on whether females viewed a digitally courting male or not (t 96 = 1.0853; p = 0.2805) or across the four treatment groups that viewed digitally courting males (F 3,73 = 1.2613, p = 0.2940). Additionally, there were no significant correlations between the length of a female’s penultimate instar stage and female selectivity in the no-choice or two-choice trials, and there were no significant correlations between the number of times a female was placed into the arena during the penultimate stage and female selectivity in no-choice or two-choice trials (p > 0.05 for all analyses). Therefore, any observed differences in female selectivity should be due to variation in whether or not female encountered males simultaneously (hereafter referred to as “simultaneous encounters”) and/or the encounter rate.
Measuring adult mate preferences
All females were fed two to three crickets the day before trials. A series of three 5-min trials initially took place when females were 10–15 days post-maturity (week 2, days 1–3). On days 1 and 2, females were placed in an arena facing a 5 min video of a courting male with small or large tufts in a no-choice trial (Fig. 1). The order of the males (i.e., small-tufted or large-tufted) was determined using systematic randomization to allow an equal number of females within each treatment group to view each male first. On day 3, females were placed in an arena and simultaneously presented two 5 min videos of courting males (Fig. 1), identical to those seen in days 1 and 2. Systematic randomization was used to allow an equal likelihood that the large-tufted male and the small-tufted male were randomly assigned to the iPod on the left side of the arena or the iPod on the right side. The center of the iPods was 12 cm from each other, and iPods were angled at 90° from each other, facing the center of the arena. Some females (n = 37) were tested again across a 4-day period (days 4–7) during their fifth week post-maturity to test for repeatability of mate preferences in two-choice trials using the same setup (Fig. 1). All trials were video recorded with a digital video camcorder (Canon ZR960). The observer was blind to which treatment group each female belonged to during video analysis. The number of female receptivity behaviors commonly observed in S. ocreata (tandem leg extensions, slow pivots, and settles) was recorded as a behavior directed to a particular male video only if the female was directly in front of the video. These receptivity behaviors have previously been shown to lead to copulation in S. ocreata (Uetz and Denterlein 1979; Scheffer et al. 1996; Montgomery 1903; Norton and Uetz 2005). Female receptivity displays are important for successful mating to take place for S. ocreata as they may signal to males that they will not be cannibalized. Additionally, mating does not typically proceed until a female signals receptivity.
Data analysis
All statistical analyses were conducted using the R statistical package (R Core Team 2013, version 2.15.3). Female “selectivity” refers to the cumulative number of receptivity displays to the courting male with large tufts relative to the cumulative number of receptivity displays to the courting male with small tufts. Statistically speaking, in week 2 trials, the number of receptivity displays to the small-tufted male and those to the large-tufted male were bound together using the “c-bind” function in R to define an individual’s selectivity in no-choice and (separately) two-choice trials. Consequently, when a generalized linear model (family = binomial) was conducted, the response variable included a matrix with two columns: the number of receptivity displays toward large-tufted males (i.e., successes) and the number of receptivity displays toward small-tufted males (i.e., failures). Initially, generalized linear models (family = binomial) were constructed using combinations of encounter rate, simultaneous encounters, and their interaction (encounter rate × simultaneous encounters). Each model was compared to an intercept-only model and compared to other models using likelihood ratio tests. This allowed us to test the initial hypothesis, for example, of whether the encounter rate (every other day or twice per day) or simultaneous encounters with males (zero, one, or three males per encounter) best predicted female selectivity. However, given the nature of the study (e.g., the instar length for penultimate females may vary among individuals), females were exposed to a wide range of males. Consequently, additional generalized linear models (family = binomial) were constructed using encounter rate, simultaneous encounters, encounter rate × simultaneous encounters, and total males (the cumulative number of courting males that a female saw during her entire penultimate stage) in all appropriate combinations for both no-choice and two-choice analyses. Each model was compared to an intercept-only model and compared to other models using likelihood ratio tests. Akaike information criterion (AIC) values were then compared to determine the combination of independent variables (encounter rate, simultaneous encounters, encounter rate × simultaneous encounters, and total males) that best fit variation in female selectivity in no-choice trials and two-choice trials during week 2.
Given the importance of total males (see “Results”) in two-choice trials from week 2, we statistically analyzed female receptivity in response to an increasing number of total males on a categorical level (receptive or not receptive) using logistic regression analyses. This allowed us to examine the effects of exposure to an increasing number of males (total males) on the probability of a female demonstrating receptivity to a male with small tufts or a male with large tufts.
We used the lme4 package (Bates et al. 2012) in R to build separate-slope, separate-intercept, and mixed (i.e., separate slope and intercept) models to examine aspects of the repeatability of female preferences. In these analyses that include week 5 results, female selectivity (response variable) was statistically defined as the difference in receptivity displays (i.e., the number of displays toward large-tufted males minus those toward small-tufted males). First, we determined if female selectivity (response variable) changed over time (fixed effect) in two-choice trials (days 3–7) after accounting for differences in individual variation (female ID: random effect). Additionally, a linear mixed-effects analysis was used to determine the relationship between the cumulative number of males that females encountered during their penultimate stage (fixed effect) and adult female selectivity (response variable), after accounting for differences in individual variation (female ID: random effect) and time (days 3–7: random effect). In all cases, a visual inspection of residual plots did not demonstrate any notable deviations from homoscedasticity or normality. Finally, a generalized linear model was run to examine the relationship between the cumulative number of males encountered during their penultimate stage and the differences in female receptivity scores averaged across all two-choice trials during week 5 (days 4–7).
We calculated the repeatability of female preferences to compare: (1) female selectivity from week 2 to week 5 and (2) female selectivity across all 4 days of week 5. In both cases, we used the “rptR” package (Nakagawa and Schielzeth 2010; Schielzeth and Nakagawa 2011) within the R statistics program. To calculate the repeatability of week 2 compared to week 5, we ran a correlation between the differences in female receptivity displays (those to large-tufted males minus those to small-tufted males) in week 2 and the mean differences in female receptivity displays of the four trials in week 5. This allowed us to calculate the correlation-based repeatability (RC; see Nakagawa and Schielzeth 2010 for further discussion). To compare female selectivity across multiple days within week 5, we calculated the ANOVA-based repeatability (RA; see Nakagawa and Schielzeth 2010 for further discussion).
Results
No-choice trials, week 2, days 1 and 2
Overall, there were no significant effects of any aspect of male availability on female selectivity in no-choice trials (Fig. 2). Specifically, likelihood ratio tests showed no significant effect of any combination of encounter rate, simultaneous encounters, and encounter rate × simultaneous encounters (Table 1). Including total males to any model still resulted in a non-significant improvement upon the intercept-only model (Table 1). Because female experience did not significantly explain variation in female selectivity during these no-choice trials, no further statistical analyses were conducted.

The mean (±SE) number of female receptivity displays to males with small tufts and males with large tufts across different treatment groups in no-choice trials
Two-choice trials, week 2, day 3
In contrast to the no-choice trials, when adult females were offered a choice between two males courting simultaneously, significant differences in female selectivity were seen across treatments (Fig. 3). A model containing encounter rate, simultaneous encounters, and their interaction was significantly better than an intercept-only model (likelihood ratio test χ 2 5 = 16.825, P = 0.0048; Table 2). Subsequent paired t tests within each treatment group revealed that females that saw three males twice per day (male-biased ratio, high frequency) had significantly more receptivity displays to males with large tufts than males with small tufts (paired t test t 17 = 4.1231, P = 0.0007). All other paired t tests within each treatment group were not significantly different (P > 0.05 in all cases). Further, the model including only simultaneous encounters was significantly better than the model including only encounter rate (likelihood ratio test: χ 2 1 = 6.061, P = 0.0138). The model with only simultaneous encounters was also significantly better than the intercept-only model (likelihood ratio test: χ 2 2 = 8.947, P = 0.0114), while the model with only encounter rate was not (likelihood ratio test: χ 2 1 = 2.886, P = 0.0894).

The mean (±SE) number of female receptivity displays to males with small tufts and males with large tufts across different treatment groups in two-choice trials. An asterisk indicates a significant difference in the number of receptivity displays to males with small tufts and males with large tufts within a particular treatment group
While a comparison of the effects of encounter rate and simultaneous encounters tests the original hypothesis, additional tests with inclusion of the cumulative number of males that females saw during their penultimate stage (total males) as a covariate significantly improved all statistical models (Table 1; all P < 0.05). Consequently, AIC values were then compared for each model (Table 2) to determine which factor of experience best predicts female receptivity. The best model has the lowest AIC value, which in this case includes simultaneous encounters and total males as factors predicting female selectivity. However, traditionally, any other model that is within two AIC units of the lowest AIC value should not be dismissed. More recently though, it has been suggested that models within 2–7 AIC units should have some support as well and analyses should often look at the AIC values for all statistical models to make a strong inference as to the importance of the factors in each model (Burnham et al. 2011). The five models with the lowest AIC values all include total males as a factor and removing this factor from any model increases the AIC value by >7 AIC units. Taken together, the cumulative number of males that penultimate females encountered best predicts female selectivity as adults in two-choice trials.
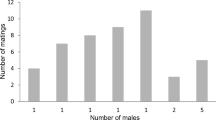
There was a significantly positive relationship between the total number of males that penultimate females saw and how selective (i.e., receptivity displays to large-tufted vs small-tufted males) they were as adults (Fig. 4; Pearson correlation: R 2 = 0.0976, N = 96, P = 0.0019). At a more categorical level (receptive or not receptive), there was a significantly increased likelihood for females to be receptive to males with large tufts if they saw more cumulative males during their penultimate stage (Fig. 5; logistic regression: Z = 2.126, P = 0.0335). While the relationship between total males and the likelihood for females to be receptive to males with small tufts is negative, it is not significant (Fig. 5; logistic regression: Z = 1.586, P = 0.1130).

The relationship between the total number of males that penultimate female Schizocosa ocreata saw (total males) and female selectivity (number of receptivity displays to males with large tufts − number of receptivity displays to males with small tufts). There was a significantly positive relationship between the total number of males that females saw during their penultimate stage and how selective they are as adults (Pearson correlation R 2 = 0.0976, N = 96, P = 0.0019)

Logistic regressions of the total number of males that penultimate female Schizocosa ocreata saw and the probability of them demonstrating receptivity to small-tufted males (gray line) or large-tufted males (black line) as an adult. There is a significantly increased likelihood for adult females to be receptive to large-tufted males if they see more cumulative males during their penultimate stage (logistic regression Z = 2.126, P = 0.0335). While the relationship between total males and the likelihood for females to be receptive to males with small tufts is negative, it is not significant (logistic regression Z = 1.586, P = 0.1130). Vertical lines indicate the standard error of the predicted probability of the logistic regression fit
Repeatability of female preferences in two-choice trials
In analyses of female selectivity across all 5 days of two-choice trials (days 3–7), there was no effect of time on female selectivity (likelihood ratio test: Χ 1,141 = 1.0544, P = 0.3045), as shown when comparing a model with a separate intercept for each individual female with and without “day” as a factor. The model accounting for a separate intercept was used due to its lower AIC value (626.65) in comparison to the separate-slope (AIC 628.79) and mixed (AIC 632.79) models.
When analyzing female selectivity as a function of the cumulative number of males, there was a significant effect of total males seen as a penultimate on adult female selectivity when accounting for multiple measures on individuals over time and accounting for difference in slopes due to the test day (likelihood ratio test: Χ 1,136 = 11.571, P = 0.0007). The model accounting for a separate slope only was chosen due to its lower AIC value (613.04) in comparison to the intercept-only (AIC 621.20) and mixed (AIC 620.29) models. Despite this evidence that the slope varies depending on which day females were tested, there was, in general, a positive relationship between the cumulative number of males that females saw during their penultimate stage and adult female selectivity across each test day for two-choice trials (Pearson correlations: week 2, day 3 R 2 = 0.0976, n = 96, P = 0.0019; week 5, day 4 R 2 = 0.0065, n = 37, P = 0.6364; week 5, day 5 R 2 = 0.1118, n = 35, P = 0.0400; week 5, day 6 R 2 = 0.1067, n = 37, P = 0.0484; week 5, day 7 R 2 = 0.0641, n = 32, P = 0.1622). Mean female selectivity across all five two-choice trials (days 3–7) significantly increased as females were exposed to an increasing cumulative number of males during their penultimate stage (Pearson correlation F 1,35 = 9.5311, r 2 = 0.2140, P = 0.0039). This relationship remained significantly positive if only two-choice trials from week 5 (days 4–7) were included (Pearson correlation F 1,35 = 6.7794, r 2 = 0.1623, P = 0.0134).
There was a significantly positive correlation between female selectivity during week 2 and mean female selectivity during week 5 (Pearson correlation N = 36, r 2 = 0.1260, P = 0.0311). The repeatability of female selectivity from week 2 to week 5 was 0.2350. The repeatability of female selectivity across week 5 (days 4–7) was 0.1620.
Discussion
Female mate preferences in adult S. ocreata were influenced by past experience in their social environment, providing evidence for socially cued anticipatory plasticity (SCAP) in mating preferences. Specifically, females displayed more receptivity displays toward large-tufted males than small-tufted males based on some aspects of past experience during their penultimate stage. Initially, experience with simultaneous encounters with males, but not encounter rate, best predicted adult female selectivity. However, females became increasingly more selective as adults if they were exposed to cumulatively more courting males during their penultimate stage, performing more receptivity displays toward large-tufted males than small-tufted males. These results support other studies in which females are predicted to be less selective when males are rare, but more selective when male abundance is high (see Eshel 1979; Hubbell and Johnson 1987, and Crowley et al. 1991 for further discussion).
As mentioned, the density-dependent effect seen in two-choice trials was due to cumulative exposure, as other variables did not have an effect on female selectivity once the cumulative number of males was incorporated into analyses. While many studies examine the effects of sex ratio and population density separately, other studies demonstrate the importance of examining sex ratio and density simultaneously in the context of male courtship (Spence and Smith 2005), copulation duration (Alonso-Pimentel and Papaj 1996), and the opportunity for sexual selection (Wacker et al. 2013). Male zebrafish, Danio rerio, alter their courtship rates in response to changes in the adult sex ratio, but only when the overall population density is high (Spence and Smith 2005). In the walnut fly, Rhagoletis juglandis, male and female density both significantly impacted copulation duration (Alonso-Pimentel and Papaj 1996). However, overall density had no significant effect once OSR was taken into account (Alonso-Pimentel and Papaj 1996). These studies demonstrate the difficulty of teasing apart effects of sex ratio and mate density on mating behaviors and suggest, along with this study, that future studies should focus on investigating the effects of what occurs at the individual level (e.g., the cumulative number of males encountered by females in a high-density or male-biased population).
This paper highlights the importance of timing and context in SCAP. As with other studies in Schizocosa spp. (Hebets 2003, 2007; Hebets and Vink 2007; Rutledge et al. 2010; Rutledge and Uetz 2014), we focused on effects of experience during the penultimate stage. In nature, female S. ocreata would likely encounter males during their penultimate stage and their adult stage, but few studies on any invertebrates to date have compared the effects of juvenile versus adult experience on mate preferences. That such plasticity can occur in adulthood based on juvenile experience alone demonstrates the importance of the penultimate stage in S. ocreata. However, these results were only seen in the context of two-choice (simultaneous presentation) trials and not no-choice (single presentation) trials. In this study, such a result might imply that female S. ocreata need to be able to visually compare potential mates in order to demonstrate plasticity (but see Hebets 2003 and Rutledge et al. 2010 where female Schizocosa spp. demonstrated experience effects in no-choice designs using live males). Additionally, in a meta-analysis on 40 species, mating preferences were significantly stronger in two-choice experimental designs than no-choice experimental designs (Dougherty and Shuker 2014). This may be due to the costs of rejecting potential mates when presented a single male at a time (see Dougherty and Shuker 2014 for detailed discussion), possibly explaining the lack of a significant effect of experience observed in no-choice presentations in this particular context.
In several taxa, females become less selective as they age (Gray 1999; Kodric-Brown and Nicoletto 2001; Uetz and Norton 2007), presumably in order to ensure that they do not miss out on mating opportunities. In S. ocreata, females at 2 weeks post-maturity prefer males with large tufts, but females at 5 weeks post-maturity no longer exhibit these preferences (Uetz and Norton 2007). This study demonstrates that the correlation between the cumulative number of males observed during the penultimate stage and adult female selectivity is retained during the fifth week post-maturity. This suggests that female preferences are repeatable despite female aging, presumably due to experience effects, which were not tested in Uetz and Norton (2007). However, females in other taxa do not always maintain their mating decisions at a later age, as interactions between age and social experience can lead to females with relaxed preferences (e.g., Tinghitella et al. 2013). While the repeatability values for female selectivity reported in this paper appear to be low, the repeatability of mating preferences is often lower in comparison to other behaviors (see review in Bell et al. 2009).
Increasing evidence that conditions in an individual’s social environment impact mating decisions (e.g., Rosenqvist and Houde 1997; Hebets 2003; Walling et al. 2008; Rutledge et al. 2010; Fowler-Finn and Rodriguez 2012) suggests that there may be selection for plasticity (i.e., instead of rigid preferences). That such plasticity is demonstrated in an invertebrate species adds to current research that demonstrates that invertebrate behaviors are not rigid in their mating preferences (butterflies: Westerman et al. 2012, 2014; crickets: Bailey and Zuk 2008; Bailey 2011; Kasumovic et al. 2012; Bailey and Macleod 2014; Atwell and Wagner 2014; fruit flies: Dukas 2005; katydids: Fowler-Finn and Rodriguez 2012; wolf spiders: Hebets 2003; Hebets and Vink 2007; Hebets 2007; Rutledge et al. 2010). Studies on the effects of the experience with the presence vs absence of experience with male courtship signals in some of these studies are biologically relevant (e.g., Bailey and Zuk 2008; Bailey 2011; Bailey and Macleod 2014). However, future studies on SCAP in all animal taxa should be sure to investigate the variation in the levels of experience (as opposed to presence vs absence), with consideration for the appropriate experimental design (e.g., experience with simultaneous vs subsequent male encounters and/or variation in encounter rate) to manipulate the perceived availability of potential mates.
Interestingly, the results from this study were based on social experience in the visual modality alone. However, male S. ocreata use both visual and vibratory courtship signals, leading to questions as to whether females demonstrate plasticity in mate preferences in response to variation in the availability of potential mates using vibratory signals as well. Vibratory signals are hypothesized to be plesiomorphic (Stratton 2005) and dominant (Hebets et al. 2013). However, within Schizocosa, courtship of male S. ocreata has a particularly high visual complexity score and has been shown to be equivalent to vibratory signals in eliciting female responses (see Uetz et al. 2009 and Hebets et al. 2013 for further discussion). The results of this study further emphasize the importance of visual signals in S. ocreata by demonstrating plasticity in female mate preferences in response to visual signals alone and add to the growing body of literature that female selectivity can be impacted by an individual’s social environment.
References
Alonso-Pimentel H, Papaj DR (1996) Operational sex ratio versus gender density as determinants of copulation duration in the walnut fly, Rhagoletis juglandis (Diptera: Tephritidae). Behav Ecol Sociobiol 39:171–180
Andersson M (1986) Evolution of condition-dependent sex ornaments and mating preferences: sexual selection based on viability differences. Evolution 40:804–816
Andersson M (1994) Sexual selection. Princeton University Press, Princeton
Atwell A, Wagner WE (2014) Female mate choice plasticity is affected by the interaction between male density and female age in a field cricket. Anim Behav 98:177–183
Bailey NW (2011) Mate choice plasticity in the field cricket Teleogryllus oceanicus: effects of social experience in multiple modalities. Behav Ecol Sociobiol 65:2269–2278
Bailey NW, Macleod E (2014) Socially flexible female choice and premating isolation in field crickets (Teleogryllus spp.). J Evol Biol 27:170–180
Bailey NW, Zuk M (2008) Acoustic experience shapes female mate choice in field crickets. Proc R Soc B 275:2645–2650
Balshine-Earn S (1996) Reproductive rates, operational sex ratios and mate choice in St. Peter’s fish. Behav Ecol Sociobiol 39:107–116
Bates D, Maechler M, Bolker B (2012). lme4: linear mixed-effects models using S4 classes. R package version 0.999999-0. http://CRAN.R-project.org/package=lme4
Bell AM, Hankison SJ, Laskowski KL (2009) The repeatability of behaviour: a meta-analysis. Anim Behav 77:771–783
Berglund A (1994) The operational sex ratio influences choosiness in a pipefish. Behav Ecol 5:254–258
Berglund A (1995) Many mates make male pipefish choosy. Behaviour 132:213–218
Bonduriansky R (2001) The evolution of male mate choice in insects: a synthesis of ideas and evidence. Biol Rev 76:305–339
Burley NT, Foster VS (2006) Variation in female choice of mates: condition influences selectivity. Anim Behav 72:713–719
Burnham KP, Anderson DR, Huyvaert KP (2011) AIC model selection and multimodel inference in behavioral ecology: some background, observations, and comparisons. Behav Ecol Sociobiol 65:23–35
Byers JA, Byers AA, Dunn SJ (2006) A dry summer diminishes mate search effort by pronghorn females: evidence for a significant cost of mate search. Ethology 112:74–80
Coleman SW, Patricelli GL, Borgia G (2004) Variable female preferences drive complex male displays. Nature 24:742–745
Crowley PH, Travers SE, Linton MC, Cohn SL, Sih A, Sargent RC (1991) Mate density, predation risk, and the seasonal sequence of mate choices: a dynamic game. Am Nat 137:567–596
Dougherty LR, Shuker DM (2014) The effect of experimental design on the measurement of mate choice: a meta-analysis. Behav Ecol. doi:10.1093/beheco/aru125
Dukas R (2005) Learning affects mate choice in female fruit flies. Behav Ecol 16:800–804
Eshel I (1979) Sexual selection, population density, and availability of mates. Theor Popul Biol 16:301–314
Fawcett TW, Johnstone RA (2003) Mate choice in the face of costly competition. Behav Ecol 14:771–779
Forsgren E (1992) Predation risk affects mate choice in a gobiid fish. Am Nat 140:1041–1049
Fowler-Finn KD, Rodriguez RL (2012) Experience-mediated plasticity in mate preferences: mating assurance in a variable environment. Evolution 66:459–468
Gordon SD, Uetz GW (2011) Multimodal communication of wolf spiders on different substrates: evidence for behavioural plasticity. Anim Behav 81:367–375
Gray D (1999) Intrinsic factors affecting female choice in house crickets: time of first reproduction, female age, nutritional condition, body size, and size-relative reproductive investment. J Insect Behav 12:691–700
Gwynne DT (1984) Sexual selection and sexual differences in Mormon crickets (Orthoptera: Tettigoniidae, Anabrus simplex). Evolution 38:1011–1022
Hebets EA (2003) Subadult experience influences adult mate choice in an arthropod: exposed female wolf spiders prefer males of a familiar phenotype. Proc Natl Acad Sci U S A 100:13390–13395
Hebets EA (2007) Subadult female experience does not influence species recognition in the wolf spider Schizocosa uetzi Stratton 1997. J Arachnol 35:1–10
Hebets EA, Vink CJ (2007) Experience leads to preference: experienced females prefer brush-legged males in a population of syntopic wolf spiders. Behav Ecol 18:1010–1020
Hebets EA, Wesson J, Shamble PS (2008) Diet influences mate choice selectivity in adult female wolf spiders. Anim Behav 76:355–363
Hebets EA, Vink CJ, Sullivan-Beckers L, Rosenthal MF (2013) The dominance of seismic signaling and selection for signal complexity in Schizocosa multimodal courtship displays. Behav Ecol Sociobiol 67:1483–1498
Hedrick AV, Dill LM (1993) Mate choice by female crickets is influenced by predation risk. Anim Behav 46:193–196
Holveck MJ, Riebel K (2010) Low-quality females prefer low-quality males when choosing a mate. Proc R Soc B Biol Sci 277:153–160
Hubbell SP, Johnson LK (1987) Environmental variance in lifetime mating success, mate choice and sexual selection. Am Nat 130:91–112
Hunt J, Brooks R, Jennions MD (2005) Female mate choice as a condition-dependent life-history trait. Am Nat 166:79–92
Iwasa Y, Pomiankowski A (1999) Good parent and good genes models of handicap evolution. J Theor Biol 200:97–109
Jennions MD, Petrie M (1997) Variation in mate choice and mating preferences: a review of causes and consequences. Biol Rev 72:283–327
Jennions MD, Petrie M (2000) Why do females mate multiply? A review of the genetic benefits. Biol Rev 75:21–64
Jirotkul M (1999) Operational sex ratio influences female preference and male-male competition in guppies. Anim Behav 58:287–294
Johnson JB, Basolo AL (2003) Predator exposure alters female mate choice in the green swordtail. Behav Ecol 14:619–625
Judge KA, Tran K, Gwynne DT (2010) The relative effects of mating status and age on the mating behaviour of female field crickets. Can J Zool 88:219–223
Kasumovic MM, Brooks RC (2011) It’s all who you know: the evolution of socially cued anticipatory plasticity as a mating strategy. Q Rev Biol 86:181–197
Kasumovic MM, Hall MD, Brooks RC (2012) The juvenile social environment introduces variation in the choice and expression of sexually selected traits. Ecol Evol 2:1036–1047
Kirkpatrick M (1996) Good genes and direct selection in evolution of mating preferences. Evolution 50:2125–2140
Kodric-Brown A, Nicoletto PF (2001) Age and experience affect female choice in the guppy (Poecilia reticulata). Am Nat 157:316–323
Kokko H, Rankin DJ (2006) Lonely hearts or sex in the city? Density-dependent effects in mating systems. Philos Trans R Soc B 361:319–334
Kuijper B, Pen I, Weissing FJ (2012) A guide to sexual selection theory. Annu Rev Ecol Evol Syst 43:287–311
Lynch KS, Rand AS, Ryan MJ, Wilczynski W (2005) Plasticity in female mate choice associated with changing reproductive states. Anim Behav 69:689–699
McClintock WJ, Uetz GW (1996) Female choice and pre-existing bias: visual cues during courtship in two Schizocosa wolf spiders (Araneae: Lycosidae). Anim Behav 52:167–181
Montgomery TH (1903) Studies on the habits of spiders, particularly those of the mating period. Proc Acad Natl Sci Phila 55:59–149
Moskalik B, Uetz GW (2011) Female hunger state affects mate choice of a sexually selected trait in a wolf spider. Anim Behav 81:715–722
Nakagawa S, Schielzeth H (2010) Repeatability for Gaussian and non-Gaussian data: a practical guide for biologists. Biol Rev 85:935–956
Norton S, Uetz GW (2005) Mating frequency in Schizocosa ocreata (Hentz) wolf spiders: evidence for a mating system with female monogamy and male polygamy. J Arachnol 33:16–24
Palokangas P, Alatalo RV, Korpimäki E (1992) Female choice in the kestrel under different availability of mating options. Anim Behav 43:659–665
R Core Team (2013) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. ISBN 3-900051-07-0, URL http://www.R-project.org/
Roberts JA, Uetz GW (2005) Information content of female chemical signals in the wolf spider, Schizocosa ocreata: male discrimination of reproductive state and receptivity. Anim Behav 70:217–223
Roberts JA, Taylor PW, Uetz GW (2007) Consequences of complex signaling: predator detection of multimodal cues. Behav Ecol 18:236–240
Rosenqvist G, Houde A (1997) Prior exposure to male phenotypes influences mate choice in the guppy, Poecilia reticulata. Behav Ecol 8:194–198
Rutledge JM, Miller A, Uetz GW (2010) Exposure to multiple sensory cues as a juvenile affects adult female mate preferences in wolf spiders. Anim Behav 80:419–426
Rutledge JM, Uetz GW (2014) Juvenile experience and adult female mating preferences in two closely related species of Schizocosa species. J Arachnol 42:170–177
Scheffer SJ, Uetz GW, Stratton GE (1996) Sexual selection, male morphology, and the efficacy of courtship signalling in two wolf spiders (Araneae: Lycosidae). Behav Ecol Sociobiol 38:17–23
Schielzeth H, Nakagawa S (2011) rptR: repeatability for Gaussian and non-Gaussian data. R package version 0.6.404/r44. http://R-Forge.R-project.org/projects/rptr/
Souroukis K, Murray A (1995) Female mating behavior in the field cricket, Gryllus pennsylvanicus (Orthoptera: Gryllidae) at different operational sex ratios. J Insect Behav 8:269–279
Spence R, Smith C (2005) Male territoriality mediates density and sex ratio effects on oviposition in the zebrafish, Danio rerio. Anim Behav 69:1317–1323
Stratton GE (2005) Evolution of ornamentation and courtship behavior in Schizocosa: insights from a phylogeny based on morphology (Araneae, Lycosidae). J Arachnol 33:347–376
Tinghitella RM, Weigel EG, Head M, Boughman JW (2013) Flexible mate choice when mates are rare and time is short. Ecol Evol 3:2820–2831
Tudor MS, Morris MR (2009) Experience plays a role in female preference for symmetry in the swordtail fish Xiphophorus malinche. Ethology 115:812–822
Uetz GW, Denterlein G (1979) Courtship behavior, habitat and reproductive isolation in Schizocosa rovneri Uetz & Dondale (Araneae: Lycosidae). J Arachnol 7:121–128
Uetz GW, Norton S (2007) Preference for male traits in female wolf spiders varies with the choice of available mates, female age and reproductive state. Behav Ecol Sociobiol 61:631–641
Uetz GW, Roberts JA (2002) Multisensory cues and multimodal communication in spiders: insights from video/audio playback studies. Brain Behav Evol 59:222–230
Uetz GW, Papke R, Kilinc B (2002) Influence of feeding regime on body size, body condition and a male secondary sexual character in Schizocosa ocreata wolf spiders (Araneae, Lycosidae): condition-dependence in a visual signaling trait. J Arachnol 30:461–469
Uetz GW, Roberts JA, Taylor PW (2009) Multimodal communication and mate choice in wolf spiders: female responses to multimodal vs. unimodal male signals in two sibling wolf spider species. Anim Behav 78:299–305
Verzijden MN, ten Cate C, Servedio MR, Kozak GM, Boughman JW, Svensson EI (2012) The impact of learning on sexual selection and speciation. Trends Ecol Evol 27:511–519
Wacker S, Mobley K, Forsgren E, Myhre LC, de Jong K, Amundsen T (2013) Operational sex ratio but not density affects sexual selection in a fish. Evolution 67:1937–1949
Walling CA, Royle NJ, Lindström J, Metcalfe NB (2008) Experience-induced preference for short-sworded males in the green swordtail, Xiphophorus helleri. Anim Behav 76:271–276
Westerman EL, Hodgins-Davis A, Dinwiddie A, Monteiro A (2012) Biased learning affects mate choice in a butterfly. Proc Natl Acad Sci U S A 109:10948–10953
Westerman EL, Chirathivat N, Schyling E, Monteiro A (2014) Mate preferences for a phenotypically plastic trait is learned, and may facilitate preference-phenotype matching. Evolution 68:1661–1670
Acknowledgments
Funding was provided by NSF Award IOS-1026995 (to GWU) and a University of Cincinnati, Biological Sciences Department Wieman-Wendell-Benedict grant (to BS). This research was conducted as part of the requirements for the Ph.D. in Biological Sciences from the University of Cincinnati. Special thanks to the undergraduate (Danielle Bailey, Rebecca Wilson) and graduate (Rachel Gilbert, Elizabeth Kozak and Alex Sweger) students in the Uetz lab for assistance with animal care, lab maintenance, and feedback. Thanks as well to Dr. Ann L. Rypstra and anonymous referees for comments on the earlier revisions of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by M. Elgar
Rights and permissions
About this article

Cite this article
Stoffer, B., Uetz, G.W. The effects of social experience with varying male availability on female mate preferences in a wolf spider. Behav Ecol Sociobiol 69, 927–937 (2015). https://doi.org/10.1007/s00265-015-1904-2
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00265-015-1904-2