
Abstract
Signaling often involves complex suites of behaviors that incorporate different sensory modalities. Whatever modality is used to establish that a signal functions in communication researchers must demonstrate that receivers respond to it. The territory defense response of male swamp sparrows involves a variety of behaviors that includes both vocal and visual displays. One of these, the “wing wave” display, is a distinctive movement that predicts physical attack. Here, we use robotic taxidermic mounts paired with song to test the hypothesis that wing waving is a signal and, specifically, that male receivers respond to wing waving as a signal of aggressive intent. As predicted, subjects responded more aggressively to the mount during wing waving trials than during stationary trials. A second experiment demonstrated that this effect cannot be attributed simply to increased attention to movement. Less expectedly, subjects did not alter their own display behavior in response to wing waving as compared to a static mount. We conclude that the wing wave display in the context of singing is a signal that functions in male–male aggressive communication. Questions remain, including whether wing waving functions as a signal in the absence of singing and whether wing waving and song are redundant signals or communicate different information.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
To show that a behavior is a signal, it is necessary though not sufficient to demonstrate that receivers respond to the behavior. How practical such demonstrations are depends strongly on signal modality. Animals use a variety of modalities to signal the following: visual, auditory, chemical, seismic, electrical, and tactile (Marler 1967; Bradbury and Vehrencamp 2011). Most animal groups emphasize a subset of the available modalities; anurans, for example, emphasize auditory signals (Gerhardt and Huber 2002), while social insects emphasize chemical ones (Wilson 1965). In birds, two signaling modalities predominate, visual and auditory. Receiver response to auditory displays in birds has long been easy to investigate through sound playback, using a variety of playback designs (Weeden and Falls 1959; King and West 1977; Krebs et al. 1978). Receiver response to static visual displays has also been easily tested, using experimental manipulations such as dyeing (Rohwer 1977; Hill 1991) or the cutting or extending of feathers (Andersson 1982; Møller 1988; Petrie and Halliday 1994). What has proven much more difficult to study is another type of visual display—display movements. Here, we examine a display movement, “wing waving,” in swamp sparrows (Melospiza georgiana), to test for receiver response using a wing waving robotic swamp sparrow.
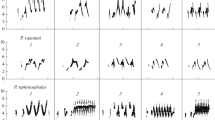
Wing waving is a distinctive display movement (Fig. 1; Supplementary Material video 1) in which sparrows raise and flutter one or both wings while remaining perched (Nice 1943; Nelson and Marler 1989). Swamp sparrows perform wing waves in the context of aggressive territory defense (Nelson and Marler 1989; Ballentine et al. 2008). In addition to wing waving, male swamp sparrows produce a variety of vocal signals during territorial aggression, including normal “broadcast” songs, rasp and wheeze calls, and low-amplitude “soft songs,” (Anderson et al. 2008; Ballentine et al. 2008). Wing waves and song are commonly produced together; in the Ballentine et al. (2008) study, for example, 93 % of the 308 bouts of wing waving which were observed were given together with soft song, broadcast song, or both (i.e., in the same 10-s block), and 7 % were given with neither category of song (B. Ballentine et al. unpublished data). Of the vocal and visual display behaviors given in response to territorial intrusion, only the numbers of soft songs and wing waves differ significantly between males that do and do not attack a conspecific model, with attackers giving more of each display than non-attackers (Ballentine et al. 2008). Soft songs and wing waves thus satisfy the “predictive” criterion for an aggressive signal (Searcy and Beecher 2009). Here, we test receiver response to wing waving in the context of singing during a territorial intrusion.

A male swamp sparrow, M. georgiana, performing the wing wave display in response to conspecific song playback
Ideally, receiver response to any display should be demonstrated experimentally, using designs in which the display of interest is manipulated independently of any other signal or cue provided by the animal, and response to the manipulated display is measured. For auditory signals, experiments of this sort have long been possible via playback of recorded sounds through loudspeakers (e.g., Martof and Thompson 1958; Weeden and Falls 1959; Wishart and Riordan 1959; Falls 1992). Playback experiments of this sort have already demonstrated response to soft song in song sparrows (Melospiza melodia) (Anderson et al. 2012), a close relative of swamp sparrows. Manipulation and playback of display movements such as wing waving have historically been more difficult. One solution has been video playback (Clark and Uetz 1992; Rowland 1995; Burford et al. 2000), but this technique is best suited for laboratory use. What seems more appropriate, especially for birds, is to use independently moving motorized models, popularly known as “robots.”
Robots have been used previously to examine the function of display movements given both alone and as part of multimodal displays (Taylor et al. 2008). In the satin bowerbird (Ptilonorhynchus violaceus), females perform a crouching movement in response to male courtship. Patricelli et al. (2002) showed using a robotic female that the crouch elicits male courtship, with males courting more intensely the more often the robot crouched. Balsby and Dabelsteen (2002) studied a jump display given by female whitethroats (Sylvia communis). Using a mechanical model, they showed that male whitethroats responded with more courtship song when the model jumped than when it did not. Narins et al. (2003) studied the aggressive display of the dart-poison frog Epipedobates femoralis, which combines calling and vocal sac movements. Experiments with an electromechanical model frog demonstrated that neither calling nor vocal sac movement alone elicits attack, but that frogs will attack a model that produces both the auditory and visual display (Narins et al. 2003). A fourth example involves the Eastern gray squirrel (Sciurus carolinensis), which gives both alarm calls and tail movements in response to predators. Partan et al. (2009) used a mechanical model squirrel to show that Eastern gray squirrels respond with alarm behavior to either the vocal or the visual component of the display and show an enhanced response when the two modalities are presented together.
To test the hypothesis that wing waving is a signal and, specifically, that receivers respond to wing waving as an aggressive signal, we conducted two experiments. First, we compared the aggressive response of territorial male swamp sparrows to either a wing waving intruder or a motionless intruder, both coupled with broadcast song playback (henceforth, the “wing waving experiment”). The intruder was a robotic taxidermic mount of a male swamp sparrow designed to perform the wing wave display. We coupled song with the visual display because wing waves are typically given with either soft song or broadcast song. We used broadcast song for playback in order to draw the subject’s attention to the robotic mount, and because broadcast song rate does not correlate with wing wave rate, with approach to the simulated intruder, or with attack likelihood (Ballentine et al. 2008). Because only wing waving differs between the wing waving and motionless treatments, any difference in response indicates response to wing waving.
In a second experiment, we controlled for the possibility that movement alone, rather than the wing wave motion in particular, affects aggressive response. This could occur because a moving mount is more likely to draw and keep a subject’s attention than a motionless mount even if the movement has no signal value, or because movement makes the mount appear more realistic. For this experiment, we presented subjects with a robotic mount designed to perform an arbitrary nonaggressive movement, turning from side to side, coupled with broadcast song playback. Here again, we compared aggressive response between trials with the turning mount versus trials with the mount motionless (henceforth, “turn-motion experiment”).
Methods
We conducted trials in a population of swamp sparrows in Conneaut Marsh, Crawford County, Pennsylvania, USA, between May 22 and June 16, 2011 (wing wave experiment, N = 30) and May 19 and June 1, 2012 (turn-motion experiment, N = 30). Birds were not individually marked. We identified territorial males by location, as territories in this species remain stable during the height of the breeding season. The two experiments were performed in consecutive years, and because we visited some of the same territories in both years, some males may have been subjects in both experiments. We completed all trials between 0600 and 1000 hours.
During both experiments, males received both a motion treatment (either wing wave or turn motion) and a stationary treatment (mount not moving). Within each experiment, tests were performed 48 h apart (except for six subjects, which received treatments 72 h apart due to weather or other logistic constraints). Whether the motion or stationary treatment was presented first was counter-balanced across subjects.
We constructed the wing waving sparrow by placing a robotic frame (designed and built by DP) actuated with a custom-built linear motor inside the body cavity of a taxidermic mount of a male swamp sparrow. The mount was affixed to a wood base in a standing position. The motor drove movement of the right wing, which waved at a speed and duration consistent with displays given by wild birds. Wing waving with one wing at a time is a common pattern in swamp sparrows (see Fig. 1). Wing waves occurred in bouts of 2.5 s at 1.2-s intervals. The turn-motion sparrow was a second taxidermic mount positioned the same as the wing waving sparrow. Its feet were affixed to a small dowel attached to a rotating motor hidden beneath the wood base. The motor turned the mount from side to side at roughly a 45° angle. Turns occurred in bouts of four (left, right, left, and right) lasting 2.0 s at 1.5-s intervals to roughly match the motion duty cycle of the wing waving sparrow (see Supplementary Material 2 for videos of both robotic mounts). For both mounts, motion was automated using electronics housed in a control box and held by an experimenter who stood approximately 10 m from the mount/loudspeaker setup. The motion routines were timed commands written in Basic and controlled with a PicAxe 28X2 microcontroller (Revolution Education, Ltd., Bath, UK). The motors were powered through two STMicroelectronics L298N H-Bridges (STMicroelectronics, Coppell, TX). The mounts were very similar visually in size and plumage characteristics. Both mounts were male swamp sparrows from the same population caught during the breeding season. The most obviously variable male plumage trait in swamp sparrows is the brightness and rustiness of the crown feathers (Greenberg 1988). The two robotic mounts did not differ noticeably in this regard. Both sparrow mounts were enclosed in a wire-mesh cage (15 cm W × 15 cm L × 17 cm H) to protect them from attacks by study subjects.
Using playback of broadcast song during trials was important for luring the subjects to the vicinity of the mount and ensuring they would notice it in a reasonable amount of time. Song playbacks consisted of a single exemplar of a swamp sparrow song presented at a rate of six songs per minute, a typical rate for this species. All song stimuli were recorded in the Conneaut Marsh population between 2003 and 2008. We played each subject a different song exemplar during each trial (i.e., 60 different song exemplars were used for the 30 pair-wise trials in each experiment). We calibrated playback amplitude to 80–83 dB at 1 m from the speaker using a BK Precision 732A sound level meter and broadcast playbacks using a Marantz 660 PMD digital recorder and an Advent Powered Partners AV570 speaker. During trials, we placed the robotic sparrow mount on a post at approximately 1 m height and approximately .5 m above the speaker.
With the exception of the motion performed by the two mounts, testing procedures were identical in the two experiments. We began trials by luring the subject to within 4 m of the speaker using playback. We played the lure song (one song exemplar) a maximum of six times (1 min total duration). If the subject did not approach within 4 m of the speaker during this time, we waited 2 min and then played the lure six more times; if the subject failed to approach within 4 m during this second set of lure songs, the trial was abandoned. Once the subject approached within 4 m of the speaker, we began the trial by restarting the playback and by starting the motion cycle if the trial was a motion treatment. A trial consisted of 2 min of song playback (12 total songs) accompanied by either a moving or stationary sparrow mount. During trials, one experimenter controlled the song playback and the mount’s motion cycle, while a second observer recorded the subject’s behaviors on a digital recorder for later transcription.
We measured the aggressive response to the mount and playback using the subject’s proximity to the mount/speaker averaged over the trial. This measure is strongly associated with attack likelihood in swamp sparrows (Ballentine et al. 2008) and related species (Searcy et al. 2006) and thus provides a good single-measure proxy for aggressiveness. In addition to proximity, we measured two display behaviors performed by subjects, wing waves and broadcast songs. With wing waves, we were especially interested in whether receivers adjust their own wing wave rate in response to an intruder that wing waves compared to one that does not, as this would be further evidence that wing waving is a signal. We did not consider soft song rate because it is strongly positively correlated with wing wave rate (Ballentine et al. 2008). Broadcast song rate does not correlate with wing wave rate or proximity and contains no information on attack likelihood (Ballentine et al. 2008). Nonetheless, it is a conspicuous auditory display performed in response to territorial intrusion, and features of broadcast song have been shown to influence male response (Nowicki et al. 2001, DuBois et al. 2009, 2011).
We sampled the subject’s proximity to the speaker at 5-s intervals and binned the samples into the categories 0–2, 2–4, 4–8, 8–16, and >16 m. Distance estimation was aided by placing markers at 2, 4, and 8 m from the speaker before the trial. We calculated each subject’s average proximity to the speaker following the method of Peters et al. (1980); we considered a bird in the 0–2-m range for a given 5-s interval to be at 1 m from the speaker during that interval, and a bird in the 2–4-m range at 3 m, in the 4–8-m range at 6 m, in the 8–16-m range at 12 m, and in the >16-m range to be at 24 m. We then averaged the proximity measures from the 5-s interval estimates across the 2-min observation period.
Wing wave displays are rapid upward flips of one or the other wing typically given in bouts of varying length and are hard to count in the field as discrete behaviors. Accordingly, we calculated wing wave rate as the number of 5-s intervals (out of a possible 24 intervals) in which any wing waves occurred (Altmann 1974). In one analysis, we asked whether subjects differed in the strength of their aggressive response to the motion and stationary treatments by conducting a within-subjects comparison of our proximity measure using paired t tests (two tailed). In a second analysis, we asked whether subjects adjusted their signaling behaviors (wing waves and songs) between treatments, also using paired t tests (two tailed). We assessed the relationship between wing waves and aggressive response across subjects using Pearson product–moment correlation coefficients. All analyses were performed in JMP Pro v10 (SAS Institute, Inc.).
Results
The majority of subjects in both experiments approached the mount/speaker within 4 m during the first lure playback (77 % of trials in the wing wave experiment and 85 % in the turn-motion experiment). Subjects met the criterion during the second lure in the remainder of trials, with one exception; this subject was tested successfully the following day.
In the wing wave experiment, subjects approached significantly closer to the mount/speaker (more aggressive) during wing waving trials than during stationary trials (t = 3.27, 29 degrees of freedom (df), p = 0.003, Fig. 2a). The numbers of wing wave displays and songs performed by subjects did not differ significantly during trials with the wing waving mount compared to trials where the mount was stationary (wing waves t = 1.53, 29 df, p = 0.136; songs t = 1.170, 29 df, p = 0.252, Fig. 2b).

Behavioral responses during trials of conspecific playback coupled with a stationary or a moving taxidermic mount. a Mean (±SE) proximity to loudspeaker/mount during trials with the stationary mount versus the wing waving mount; b mean (±SE) broadcast songs (dark) and bouts of wing waves (light) during stationary and wing waving trials; c mean (±SE) proximity to loudspeaker/mount during trials with the stationary mount versus the turn-motion mount; d mean (±SE) broadcast songs (dark) and bouts of wing waves (light) during stationary and turn-motion trials; **p < 0.01
In the turn-motion experiment, subjects responded with similar aggressiveness during motion and stationary trials (comparing averaged distance to the mount/speaker, t = 0.81, 29 df, p = 0.428, Fig. 2c). Subjects also gave similar rates of wing wave displays (t = 0.585, 29 df, p = 0.576) and songs (t = 0.24, 29 df, p = 0.812, Fig. 2d) during motion and stationary trials.
As in Ballentine et al. (2008), we found a positive relationship between aggressiveness and wing waves during trials with the stationary mounts. Subjects that were more aggressive (lower proximity scores) produced more wing wave displays (stationary wing wave mount, Pearson r s = −0.46, p = 0.011, Fig. 3a; stationary turn-motion mount, r s = −0.42, p = 0.023, Fig. 3b). This relationship did not occur, however, during trials with the moving mounts (wing wave mount r s = −0.038, p = 0.84, Fig. 3c; turn-motion mount r s = −0.15, p = 0.443, Fig. 3d). In the case of the wing waving trials, this effect was largely due to subjects approaching the mount more closely and producing wing waves regardless of their proximity.

Relationships between bouts of wing wave displays given by subject birds and the subjects’ proximity to the loudspeaker/mount during trials with moving and stationary mounts. The wing waving mount with movement disabled was used as the stationary mount for the wing wave experiment, and the turn-motion mount with movement disabled was used as the stationary mount for the turn-motion experiment. The panels show subjects’ wing waves and proximity during trials with the stationary mount in the wing wave experiment (a), the stationary mount in the turn-motion experiment (b), the wing waving mount (c), and the turn-motion mount (d)
We found no effect of year when comparing responses during the stationary mount trials between the two experiments. Subjects responded similarly to the stationary mounts in the 2 years using both aggression and signaling measures (proximity to speaker, 2011 vs. 2012 mean ± standard error (SE) = 3.42 ± 0.47 m vs. 2.62 ± 0.30 m; t = 0.60, 58 df, p = 0.551) wing waves (5.3 ± 1.1 vs. 6.37 ± 1.4 bouts; t = 0.6, 58 df, p = 0.585), and songs (2.0 ± 0.62 vs. 0.77 ± 0.41 songs; t = 1.65, 58 df, p = 0.104).
Discussion
Determining whether a putative signal conveys aggressive intent requires demonstrating that (1) the signal increases in aggressive contexts, (2) the signal predicts aggressive escalation by the signaler, and (3) receivers show differential responses to the signal versus a control stimulus (Searcy and Nowicki 2005; Searcy and Beecher 2009). The wing wave display in swamp sparrows satisfies the first two criteria as it occurs predominantly during aggressive interactions and has been shown to predict subsequent physical attack by the signaler (Ballentine et al. 2008). Here, we provide evidence for the third criterion, showing that male receivers respond more aggressively to a simulated singing and wing waving intruder compared to an intruder that sings only. Because only the presence and absence of wing waving differ between these two treatments, the difference in aggressive response to the treatments demonstrates that male swamp sparrows respond to wing waving. We also show that motion of the mount alone cannot account for this difference, as subjects did not respond more aggressively to a mount that turned side to side compared to the mount when stationary. Taken together, our results support the hypothesis that the wing wave display in swamp sparrows is perceived as an aggressive signal. As we did not test response to wing waving as an isolated display, we must be circumspect about whether wing waving by itself would affect receivers in a similar way.
Our measure of aggressive response was the subjects’ proximity to the mount/speaker setup, and we also measured wing waves and broadcast songs given by subjects (discussed below). We focused on proximity as the sole measure of aggressiveness because proximity is a strong predictor of attack in this species and thus directly measures aggressive intentions. Attacks are obviously the most direct measure of all, but only three of the 60 birds we tested attacked the caged mount in the course of 120 trials, so we did not include this measure. We also did not measure the number of soft songs, which, however, strongly correlates with two behaviors that we did measure, wing waves and proximity. Soft songs may prove useful in future studies that seek to test additional questions about how the wing wave display is perceived, for example, when paired with soft song versus broadcast song.
The within-subjects comparisons of wing wave rates between motion and stationary trials showed that on average, subjects are consistent in their wing waving behavior across treatments. However, in line with Ballentine et al. (2008), we found a positive relationship across subjects between wing waving and aggressiveness during stationary mount trials such that individuals that approached closely produced many wing waves, and those that did not approach produced few or no wing waves. This relationship did not arise during trials with the moving mounts.
In the case of the wing waving experiment, one explanation for this result is that the addition of the wing waving movement to the mount simulated a more realistic intruder and resulted in subjects responding more aggressively overall, including closer approach and more wing wave displays. In addition, wing waving may serve as part of a hierarchical signaling system in swamp sparrows. Hierarchical signaling systems are known for several bird species, in which signals are switched or added to escalate the encounter (e.g., Beecher et al. 1996; Burt et al. 2001). In song sparrows, a close relative of swamp sparrows, wing waving has been hypothesized to occur along with soft song at or near the top of the hierarchy of aggressive signals (Ç. Akçay et al. unpublished). The same may be true in swamp sparrows; wing waves reliably predict attack (Ballentine et al. 2008), so it is reasonable to conclude that wing waving is a highly escalated signal that evokes a highly escalated response from subjects.
In addition to the wing wave display, we also examined the rate of broadcast songs given by subjects. Song rate appears to convey no information on attack likelihood in swamp sparrows (Ballentine et al. 2008), yet an increase in broadcast singing is a typical response to a territorial challenge. This behavior could be explained by the “multiple messages” hypothesis, which states that different signals convey different kinds of information or information about different aspects of signaler quality (Møller and Pomiankowski 1993; Johnstone 1996; Hebets and Papaj 2005). The rate of broadcast songs, for example, might contain information on the singer’s condition (Davies and Lundberg 1984; Gottlander 1987) rather than on its aggressive intentions, information that would still be of interest to aggressive opponents. Alternatively, the intended receiver of broadcast songs could be neighboring males, the signaler’s social mate, or other nearby females that are potential extra-pair mates. Broadcast song rate has not been examined with regard to male qualities in swamp sparrows, but it has been shown that both male and female receivers respond to features of song such as vocal performance (Ballentine et al. 2004; DuBois et al. 2009, 2011), and vocal performance predicts male age and size (Ballentine 2009). Additional work is needed, then, to determine what information is being signaled by broadcast songs during agonistic interactions, and how the message may differ when broadcast songs are paired with wing waves.
Of the various behaviors, male swamp sparrows perform in response to a rival male, two displays, wing waves and soft songs, predict attack, and thus appear to be signals of aggressive intentions. These displays are sometimes given on their own but are most frequently given together. We suggest that the combined display constitutes a case of multimodal signaling (Rowe 1999; Candolin 2003; Hebets and Papaj 2005; Partan and Marler 2005). In some cases of multimodal signaling, signals in two modalities must be given together because of linked production mechanisms; an example is the coupling together of the auditory signal of male calling and the visual display of vocal sac inflation in male anurans (Narins et al. 2003; Pauly et al. 2006). In other cases, signals in two modalities are not coupled together by production mechanisms and so can be given alone as well as together; an example here is the combination of tail flagging and alarm calling used by gray squirrels in response to predators (Partan et al. 2009). Wing waving and soft song in swamp sparrows fall in the latter category; though they are strongly associated with each other, there is no link between their production mechanisms.
Various hypotheses have been proposed to explain the evolution and maintenance of multimodal signals (Hebets and Papaj 2005; Partan and Marler 2005). Ballentine et al. (2008) suggested that wing waves and soft song fit with the “redundant signals” hypothesis, which states that multiple displays broadcast the same information, either to intensify the signal (Hasson 1989; Rowe 1999) or to counteract error-prone signals (Møller and Pomiankowski 1993; van Doorn and Weissing 2006). In either case, receivers gain more information by attending to both signals rather than to either alone. Further, it may be advantageous for an animal to use different signaling modalities for redundant signals, so that one signal can be perceived if the other cannot, as suggested by the efficacy backup hypothesis (Hebets and Papaj 2005). We speculate that soft songs and wing waves may work as redundant signals in this way. Soft songs are defined by their low amplitude, and because of this trait, they are easily masked by other sounds or noise. Thus, wing waves may serve to draw and keep the receiver’s attention so that soft songs may be heard, as well as to amplify the aggressive message. Wing waving may often be visually occluded by reeds and other vegetation in the swamp sparrow’s habitat, making an auditory backup important. This hypothesis could be tested by measuring the latency to respond to either signal in isolation compared to the signals combined or by examining the strength of response over a longer period to test whether response wanes differentially to single versus combined signals. As wing waves are also given with broadcast songs, future studies should test whether receiver response differs when wing waves are paired with soft songs versus broadcast songs.
In their review of complex signal function, Hebets and Papaj (2005) suggest that four questions are a key to understanding how a multiple signaling system functions: (1) Do the signals covary and is the relationship positive or negative? (2) Do the signals covary with some attribute of the signaler? (3) Are the signals necessary or sufficient to elicit a receiver response? (4) Does the presence of one signal influence the receiver’s response to a second? We are making progress toward answering these questions for the swamp sparrow, allowing us to narrow in on possible hypotheses for the function of complex display in this species. With regard to the first two questions, we know that wing waves and soft songs covary positively with each other and with aggressiveness (Ballentine et al. 2008), while broadcast songs do not predict aggression. With regard to the third question, we show in this study that wing waves, in the absence of soft songs, are sufficient to elicit a strong aggressive response from receivers. What remains to be tested is whether wing waves alone in the absence of any vocalization are sufficient to elicit aggression, whether soft song is sufficient to elicit aggression in swamp sparrows as it is in song sparrows (Anderson et al. 2012), and whether the combination of soft songs and wing waves is more effective than either signal alone.
References
Altmann J (1974) Observational study of behavior: sampling methods. Behaviour 49:227–266
Anderson RC, Searcy WA, Nowicki S (2008) Soft song in song sparrows: acoustic structure and implications for signal function. Ethology 114:662–676
Anderson RC, Searcy WA, Hughes M, Nowicki S (2012) The receiver-dependent cost of soft song: a signal of aggressive intent in songbirds. Anim Behav 83:1443–1448
Andersson M (1982) Female choice selects for extreme tail length in a widowbird. Nature 99(818):820
Ballentine B (2009) The ability to perform physically challenging songs predicts age and size in male swamp sparrows, Melospiza georgiana. Anim Behav 77:973–978
Ballentine B, Hyman J, Nowicki S (2004) Vocal performance influences female response to male bird song: an experimental test. Behav Ecol 15:163–168
Ballentine B, Searcy WA, Nowicki S (2008) Reliable aggressive signalling in swamp sparrows. Anim Behav 75:693–703
Balsby TJS, Dabelsteen T (2002) Female behaviour affects male courtship in whitethroats, Sylvia communis: an interactive experiment using visual and acoustic cues. Anim Behav 63:251–257
Beecher MD, Stoddard PK, Campbell SE, Horning CL (1996) Repertoire matching between neighbouring song sparrows. Anim Behav 51:917–923
Bradbury J, Vehrencamp S (2011) Principles of animal communication. Sinauer, Sunderland
Burford FRL, McGregor PK, Oliveira RF (2000) Response of fiddler crabs (Uca tangeri) to video playback in the field. Acta Ethol 3:55–59
Burt JM, Campbell SE, Beecher MD (2001) Song type matching as threat: a test using interactive playback. Anim Behav 62:1163–1170
Candolin U (2003) The use of multiple cues in mate choice. Biol Rev 78:575–595
Clark DL, Uetz GW (1992) Morph-independent mate selection in a dimorphic jumping spider: demonstration of movement bias in female choice using video-controlled courtship behaviour. Anim Behav 43:247–254
Davies NB, Lundberg A (1984) Food distribution and a variable mating system in the dunnock, Prunella modularis. J Anim Ecol 53:895–912
DuBois AL, Nowicki S, Searcy WA (2009) Swamp sparrows modulate vocal performance in an aggressive context. Biol Lett 5:163–165
DuBois AL, Nowicki S, Searcy WA (2011) Discrimination of vocal performance by male swamp sparrows. Behav Ecol Sociobiol 65:717–726
Falls J (1992) Playback: A historical perspective. In: McGregor P (ed) Playback and studies of animal communication. Plenum, New York, pp 11–33
Gerhardt H, Huber F (2002) Acoustic communication in insects and anurans. University of Chicago Press, Chicago
Gottlander K (1987) Variation in the song rate of the male pied flycatcher Ficedula hypoleuca: causes and consequences. Anim Behav 35:1037–1043
Greenberg R (1988) Seasonal plumage dimorphism in the swamp sparrow (Melospiza georgiana). J Field Ornithol 59:149–154
Hasson O (1989) Amplifiers and the handicap principle in sexual selection: a different emphasis. Proc R Soc B 235:383–406
Hebets EA, Papaj DR (2005) Complex signal function: developing a framework for testable hypotheses. Behav Ecol Sociobiol 57:197–214
Hill GE (1991) Plumage coloration is a sexually selected indicator of male quality. Nature 350:337–339
Johnstone RA (1996) Multiple displays in animal communication: ‘backup signals’ and ‘multiple messages’. Phil Trans R Soc B 351:329–338
King AP, West MJ (1977) Species identification in the North American cowbird: appropriate responses to abnormal song. Science 195:1002–1004
Krebs JR, Ashcroft R, Webber M (1978) Song repertoires and territory defence in the great tit. Nature 271:539–542
Marler P (1967) Animal communication signals. Science 157:769–774
Martof BS, Thompson EF (1958) Reproductive behaviour of the chorus frog, Pseudacris nigrita. Behaviour 13:243–258
Møller AP (1988) Female choice selects for male sexual tail ornaments in the monogamous swallow. Nature 332:640–642
Møller AP, Pomiankowski A (1993) Why have birds got multiple sexual ornaments? Behav Ecol Sociobiol 32:167–176
Narins PM, Hödl W, Grabul DS (2003) Bimodal signal requisite for agonistic behavior in a dart-poison frog, Epipedobates femoralis. Proc Natl Acad Sci USA 100:577–580
Nelson DA, Marler P (1989) Categorical perception of a natural stimulus continuum: birdsong. Science 244:976–978
Nice MM (1943) Studies in the life history of the song sparrow II: the behavior of the song sparrow and other passerines. Trans Linn Soc NY 6:1–328
Nowicki S, Searcy WA, Hughes M, Podos J (2001) Motor performance and the evolution of birds song: male and female response to song innovation in swamp sparrows. Anim Behav 62:1189–1195
Partan SR, Marler P (2005) Issues in the classification of multimodal communication signals. Am Nat 166:231–245
Partan SR, Larco CP, Owens MJ (2009) Wild tree squirrels respond with multisensory enhancement to conspecific robot. Anim Behav 77:1127–1135
Patricelli GL, Uy JAC, Walsh G, Borgia G (2002) Sexual selection: male displays adjusted to female's response. Nature 415:279–280
Pauly GB, Bernal XE, Rand AS, Ryan MJ (2006) The vocal sac increases call rate in the Tungara frog Physalaemus pustulosus. Physiol Biochem Zool 79:708–719
Peters S, Searcy WA, Marler P (1980) Species song discrimination in choice experiments with territorial male swamp and song sparrows. Anim Behav 28:393–404
Petrie M, Halliday T (1994) Experimental and natural changes in the peacock's (Pavo cristatus) train can affect mating success. Behav Ecol Sociobiol 35:213–217
Rohwer S (1977) Status signaling in Harris sparrows: some experiments in deception. Behaviour 61:107–129
Rowe C (1999) Receiver psychology and the evolution of multicomponent signals. Anim Behav 58:921–931
Rowland WJ (1995) Do female stickleback care about male courtship vigor? Manipulation of display tempo using video playback. Behaviour 132:951–961
Searcy WA, Beecher MD (2009) Song as an aggressive signal in songbirds. Anim Behav 78:1281–1292
Searcy W, Nowicki S (2005) The evolution of animal communication: reliability and deception in signaling systems. Princeton University Press, Princeton
Searcy WA, Anderson RC, Nowicki S (2006) Bird song as a signal of aggressive intent. Behav Ecol Sociobiol 60:234–241
Taylor RC, Klein BA, Stein J, Ryan MJ (2008) Faux frogs: multimodal signalling and the value of robotics in animal behaviour. Anim Behav 76:1089–1097
van Doorn GS, Weissing FJ (2006) Sexual conflict and the evolution of female preferences for indicators of male quality. Am Nat 168:742–757
Weeden JS, Falls JB (1959) Differential responses of male ovenbirds to recorded songs of neighboring and more distant individuals. Auk 76:343–351
Wilson EO (1965) Chemical communication in the social insects. Science 149:1064–1071
Wishart G, Riordan DF (1959) Flight responses to various sound by adult males of Aedes aegypti L. (Diptera, Culicidae). Can Entomol 91:181–191
Acknowledgments
We thank Jason Dudley at Whistlin’ Wings Taxidermy, Raleigh, North Carolina, USA, for preparing the taxidermic mount used in the wing waving experiment, and Meng Kang and Martin Steren for their assistance with the design and creation of the actuator mechanism and controller used to animate this mount. We thank Susan Peters for her support throughout the study and for her help with preparing playback stimuli. Robert Lachlan kindly provided his photo of a wing waving swamp sparrow. Thanks to three anonymous reviewers whose suggestions improved the manuscript. Funding was provided by the Duke University Office of the Provost; access to field sites was granted by the Pennsylvania State Game Commission, and logistical support in the field was provided by the Pymatuning Laboratory of Ecology of the University of Pittsburgh.
Ethical standards
All work conforms to the ABS/ASAB guidelines for the treatment of animals in behavioural research and teaching and was approved by the Duke Institutional Animal Care and Use Committee (protocol no. A061-11-03) and the University of Pittsburgh Institutional Animal Care and Use Committee (protocol no. 12030268).
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by D. Rubenstein
Electronic supplementary material
Below is the link to the electronic supplementary material.
(M4V 7801 kb)
(M4V 17651 kb)
Rights and permissions
About this article
Cite this article
Anderson, R.C., DuBois, A.L., Piech, D.K. et al. Male response to an aggressive visual signal, the wing wave display, in swamp sparrows. Behav Ecol Sociobiol 67, 593–600 (2013). https://doi.org/10.1007/s00265-013-1478-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00265-013-1478-9