
Abstract
Purpose
The purpose of this study was to determine the effect of exposure to pulsed electromagnetic fields (PEMF) on modulation of the cartilage apoptosis signalling pathway in ovariectomised rats by monitoring the expression of mRNA of X-linked inhibitor of apoptosis protein (XIAP) and Bax.
Methods
Forty-eight female Sprague-Dawley rats (250 ± 50 g) were randomly assigned to one of four groups: ovariectomy with PEMF treatment (PEMF group), ovariectomy with oestradiol (E2) treatment (oestrogen group), ovariectomy control (OVX group) and sham group. The ovariectomy model was prepared by surgical resection of the ovaries. After a three-month intermission, animals in the PEMF and oestrogen groups received treatment for 30 days; then serum 17β-oestradiol levels, chondrocyte morphology, and XIAP and Bax mRNA expression in knee joint cartilage were analysed.
Results
The results showed different chondrocyte formation in each group. Serum E2 content in the PEMF and oestrogen groups was significantly higher than in the OVX group (p < 0.05). The expression of XIAP mRNA in the PEMF and oestrogen groups was significantly up-regulated compared to the OVX group, while that of Bax mRNA was significantly down-regulated (p < 0.05). The correlation between E2 level and expression of Bax mRNA was positive (0.506) and statistically significant (p < 0.001).
Conclusion
These data demonstrate that PEMF can up-regulate XIAP mRNA expression and down-regulate Bax mRNA expression in ovariectomised rats. Changes in XIAP and Bax mRNA expression may be the mechanism by which PEMF therapy affects postmenopausal osteoarthritis.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Osteoarthritis (OA; see list of abbreviations in Table 1) is a common degenerative disease of human articular cartilage and contributes greatly to medical care costs and disability in the elderly. There is considerable epidemiological evidence suggesting that sex hormones, primarily oestrogen, play a part in the development of OA in women. Previous studies have reported that oestrogen may affect osteoarthritis through its effect on cytokines and growth factors that have a potential role in cartilage metabolism [1, 2]. In addition, there seems to be a relationship between the development of OA in women and sex steroid hormone levels, with the incidence of OA increasing at the time of the menopause, suggesting that female oestrogen loss after menopause might be a risk factor for OA development whereas oestrogen may be protective [3, 4]. Some studies have demonstrated that the adapted ovariectomised (OVX) model has potential as a useful in vivo model for postmenopausal OA [5, 6].
Conventional therapies for OA aim to reduce pain and progression of joint damage in order to minimise the disability and maximise the quality of life. Pulsed electromagnetic fields (PEMFs) have already been considered as a proven remedy for delayed union fractures [7]. The European League Against Rheumatism (EULAR) has now rated pulsed electromagnetic field treatment for OA as 1B evidence; it received a B rating for strength of recommendation [8].
In vitro studies have shown that chondrocyte proliferation and matrix synthesis are significantly enhanced by PEMF stimulation [9–11]. In a previous study, we showed that PEMFs have a systemic effect on oestrogen metabolism in ovariectomised rats [12]. Mechanisms related to this effect inhibit chondrocyte apoptosis and downregulate MMP13 expression in knee joint cartilage.
Two major apoptosis signalling pathways exist, the death receptor (extrinsic) pathway and the mitochondrial (intrinsic) pathway [13]. A central event in the process of apoptosis is the activation of caspases, a family of proteases that are ubiquitously expressed as weakly active zymogens [14, 15]. One important family of negative caspase regulators is represented by ‘inhibitor of apoptosis’ proteins (IAPs) [16]. The best studied mammalian IAP is the X-linked inhibitor of apoptosis (XIAP), which is considered the most potent caspase inhibitor in vitro [17]. XIAP can inhibit a broad range of caspases that act within both mitochondrial and death receptor signalling pathways [18]. Members of the Bcl-2 family are among the first groups of proteins recognised as having the ability to regulate apoptosis. Bax, a pro-apoptotic member of the Bcl-2 family, is a cytosolic protein that inserts into mitochondrial membranes upon induction of cell death [19]. Since XIAP and Bax are both crucial in the regulation of the mitochondrial pathway of apoptosis, expression profiling of XIAP and Bax genes in joint cartilage might reveal a specific expression pattern of these genes in OA.
To our knowledge, the effects of PEMF on apoptosis signalling pathways in articular chondrocytes in OVX models of postmenopausal OA have not been clearly defined. The purpose of this study was to investigate the potential mechanisms responsible for chondrocyte apoptosis by studying the effects of PEMF on the expression of XIAP and Bax mRNA in an OVX rat model. With this model, we can further explore the mechanism of PEMF in the treatment of oestrogen deficiency-induced OA.
Materials and methods
Experimental model
Three-month-old, non-pregnant female Sprague-Dawley rats (purchased from the experimental animal center of West China Hospital, Sichuan University, certificate number 0003235) weighing 250 ± 50 g were randomly divided into the following four groups according to random digits tables: ovariectomised rats with PEMF treatment (PEMF group), ovariectomised rats with oestrogen treatment (oestrogen group), ovariectomised controls (OVX group) and sham-operated animals (sham group). All rats were subjected to bilateral OVX except those in the sham group; each group included 12 rats. All experimental procedures were approved by the animal care handling committee of Sichuan University and conformed to guidelines for the care and use of laboratory animals as adopted by the “Animal Protection Law of the People's Republic of China (2001)”.
Reagents and instruments
The following reagents and instruments were used in this study: E2 (conjugated E2, Favor-clan Hecto-Palace Pharmaceutical Factory, Soochow, China); Trizol reagent for RNA isolation (MRC, USA); RT-PCR Kit (Revert Aid™ First Strand cDNA Synthesis Kit, Lithuania); Union2000 electromagnetic field therapeutic instrument (Academy Institute of Medical Sciences Biomedical Engineering of China; Scientific Research East Lu No.7, Nakai region, Tianjin, China); FTC2000 fluorescence quantitative gene amplification instrument (FUNGLYN, Fengling, CA).
OA model and specimen preparation
Bilateral OVX was performed through a sagittal medial laparotomy under general anaesthesia with intraperitoneal injection of 5% chloraldurat, as previously described [20]. In the sham group, the surrounding fatty tissue of both ovaries was excised. Antibiotic prophylaxis with cefonicid (100 mg/kg) was administered before and for five days following surgery. Each group was housed in cages at room temperature (20–26°C) and humidity of 60–70% under a 12/12 hour light/dark cycle with free access to water and food. Each rat was intramuscularly injected with penicillin (400,000 units) once every day for four days to prevent infection.
Electromagnetic stimulation system
PEMF was generated by a commercial, clinically approved proprietary device, Union-2000 (Institute of Biomedical Engineering, Chinese Academy of Medical Sciences, China). Twelve weeks after surgery, the PEMF group rats (whole body) were exposed to pulsed electromagnetic fields with the following specifications: field frequency of 8 Hz, intensity of magnetism of 3.8 mT, 40 min per day for 30 days, with free running during exposure. Twelve weeks after surgery, the E group received premarin (0.0625 mg/kg/d; intragastric administration 1–2 ml/100 g). Drugs were diluted into suspension with stroke-physiological saline solution. As the same time, the E, OVX and sham group rats were exposed to placebo PEMF by placing them inside the instrument for the same periods of time as the PEMF group but without activating the on switch.
Detection of serum E2 levels
After thirty days of intervention, serum was collected from all experimental animals and supernatants (2 ml) were extracted and stored at −20°C. E2 levels were detected using an electrochemiluminescence immunoassay (ECLIA) following the manufacturer’s instructions (Roche, USA).
Tissue preparation
After 30 days, all rats were sacrificed under ether anaesthesia. The articular capsule of the knee joint was excised and the medial condyle of the femur and the tibial plateau cartilage were separated. The tissue was prepared respectively as follows to observe chondrocyte morphology and reverse transcriptase-polymerase chain reaction (RT- PCR). Part of the tissue was further processed for analysis using neutral buffer formalin fixation (40 g/L), disodium ethylenediamine tetraacetic acid decalcification (100 g/L), and renoveatur decalcifying fluid on alternate days for two weeks, successive ethyl alcohol dehydration, dimethyl benzene transparence, and paraffin imbedding. Several microtome sections (5 μm) were collected from each tissue for morphological observation. The cartilaginous tissue was stained with haematoxylin and eosin (H&E) and mounted with neutral balsam for observation of chondrocyte morphology by light microscopy. The other freshly prepared cartilage tissue was used for preparation of total cellular RNA by RT-PCR (Molecular Research Center, Sichuan University, Chengdu, China) using the manufacturer's recommended method.
Histopathological study
The sections were stained with haematoxylin and eosin for histopathological studies. Two pathologists blindly evaluated all histological changes in the articular cartilages and enumerated histopathological findings obtained by light microscopy based on OA cartilage histopathology grade assessment [21]. The histopathological findings included flaking, superficial fibrillation, chondrocyte enlargement, hyalinisation, deep fibrillation, pitting, partial cartilage collapse, and total cartilage collapse.
Determination of gene expression by reverse-transcriptase polymerase chain reaction and real-time PCR
Total RNA (1 μg) from cartilaginous tissue was reversed-transcribed using the RevertAid™ (MBI) reverse transcribe kit (GeneAmp, Perkin-Elmer, Branchburg, NJ, USA) following the manufacturer’s protocol. Freshly reverse-transcribed cDNA was used for real-time PCR, using SYBR Green (Bio-Rad, Hercules, CA) to monitor DNA synthesis. The following rat primers were used (see Table 2; primer sequences and PCR product lengths): XIAP—forward: 5' CTCCAAGAAATCCAGCCAT 3', reverse: 5' CCTCCTCCACAGTGAAAG 3' (the transcribed sequence corresponds to 151 bases); Bax—forward: 5' GTTTCATCCAGGATCGAGCA 3', reverse: 5' CAATCATCCTCTGCAGCTCC 3' (the transcribed sequence corresponds to 156 bases); β-actin—forward: 5' GCCAACACAGTGCTGTCT 3', reverse: 5' AGGAGCAATGATCTTGATCTT 3' (sequence corresponding to 114 bases).
PCR was carried out using the FTC2000 amplification system (FUNGLYN, Fengling, Shanghai) with the following cycling protocol: 94°C denaturation step for two minutes, followed by 20 cycles of 94°C denaturation for 20 seconds, 48°C annealing for 20 seconds and 72°C extension for one minute. Detection of the fluorescent product was carried out at the end of the 72°C extension period. All the PCR products were preserved at −20°C until use.
The quantitative measurement of the expression of each gene was normalised to the amount of a housekeeping gene (β-actin) cDNA. To confirm amplification specificity, the PCR products were subjected to melting curve analysis, and the data were analysed and quantified using software for the calculation of relative expression in real-time PCR with pairwise fixed reallocation randomisation test (Relative Expression Software Tool). The ΔCt value for each sample was obtained by subtracting the Ct value of β-actin.
Statistical analysis
All values are expressed as mean ± standard deviation (SD). The effects of PEMF treatment on XIAP and Bax expression were first analysed by one-way ANOVA. When statistical differences were detected, Student’s t-test was used. The correlation of E2 levels and mRNA expression of XIAP and Bax was analysed by Bivariate correlations analysis, respectively. Statistics were performed by analysis of variance using the SPSS version 16.0 statistical software (SPSS, Inc., Chicago, IL, USA). Differences were considered significant when p < 0.05.
Results
Post-intervention serum E2 levels for all groups
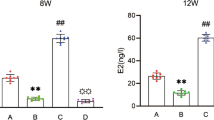
After interventions, levels of serum E2 in the oestrogen group were significantly higher than those in the OVX group (36.23 ± 9.35 pg/ml vs.
25.65 ± 8.54 pg/ml) (p < 0.05). Interestingly, PEMF treatment has a similar effect to oestradiol replacement. Levels of serum E2 in the PEMF group were statistically higher than in the OVX group (35.35 ± 8.97 pg/ml vs. 25.65 ± 8.54 pg/ml; p < 0.05); there was no significant difference between the levels in the PEMF and oestrogen groups (35.35 ± 8.97 pg/ml vs. 36.23 ± 9.35 pg/ml; p > 0.05). Serum E2 levels in the PEMF, oestrogen, and OVX groups were statistically lower than in the sham group (p < 0.05) (see Table 3).
Comparison of chondrocyte morphology
In the sham group, chondrocyte morphology was free of histopathological changes (Fig. 1a). Healthy-appearing cartilage has a smooth surface and its chondrocytes are arranged in columns. According to the recommended OA grading method [21], Fig. 1a was assessed Grade 0. The matrix and associated chondrocytes were organised in three appropriately orientated, well ordered zones. No enlargement or distortion of chondrons and no proliferative changes of chondrocytes were observed. In the PEMF (Fig. 1b) and oestrogen groups (Fig. 1c), the cartilage surface was smooth, surface cartilage cells reduced, with cartilage cells arranged neatly and cartilage cell clusters present. Figure 1b and c were assessed as Grade 1 and showed retention of the articular surface layer, which were the characters of the threshold for OA. In the OVX group, coarse articular cartilage surfaces, partial chondrocyte abolition, cartilage cell clusters and fissuring were detected (Fig. 1d). The OVX group showed increased surface erosion and cartilage degeneration compared to the PEMF and oestrogen groups. Figure 1d was assessed Grade 2, which was characterised by focal discontinuity of the cartilage superficial zone.

a H&E stain of cartilage tissue for the sham group, ×200 magnification Grade 0 (arrows represented the smooth cartilage surface and organised chondrocytes). b H&E stain of cartilage tissue for the PEMF group, ×200 magnification Grade 1 (arrows represented retention of the articular surface layer). c H&E stain of cartilage tissue for the estrogen group, ×200 magnification Grade 1 (arrows represented retention of the articular surface layer). d H&E stain of cartilage tissue for the OVX group, ×200 magnification Grade 2 (arrows represented focal discontinuity of the cartilage superficial zone)
PEMF regulates the expression of XIAP and Bax mRNA
To investigate the possible basis for PEMF-regulated pathomorphological changes in chondrocytes, we measured the expression of XIAP and Bax mRNA by RT-PCR. The intersection of each amplification curve with the threshold yielded as a CT (threshold cycle) value that reflected the relative amount of the original mRNA and cDNA. The more the initial transcript, the lower the CT value is. The initial normalised value in each sample was represented by the ΔCT (target CT -β-actin CT [housekeeping gene]).
In PEMF-treated ovariectomised animals, the expression of XIAP mRNA was significantly increased compared to its expression in sham and OVX groups, while the level of Bax mRNA showed decreased expression compared to sham and OVX groups. There was no significant difference in the levels of XIAP or Bax mRNA in the PEMF and oestrogen groups. Thus, PEMF treatment may regulate the expression of anti-apoptotic and apoptotic proteins in chondrocytes (Fig. 2).

Effect of PEMF on expression of XIAP and Bax genes in cartilage tissue. The means of △CT for XIAP and Bax expression in the four groups are shown. The expression levels of mRNA were determined using real-time PCR. Real-time PCR analysis showed a significant decrease in XIAP mRNA levels in the PEMF and oestrogen groups, and a significant increase in Bax mRNA levels in the PEMF and oestrogen groups compared to the sham and OVX groups (p < 0.05) (n = 12 in each group)
The correlations of E2 levels and mRNA expression of XIAP and Bax
The correlation of E2 levels, mRNA expression of XIAP and Bax are reported in Table 4. The results showed the correlation between E2 level and mRNA expression of XIAP is negligible (−0.059) and not statistically significant (p = 0.690). However, the Pearson correlation coefficient measured the linear association between E2 levels and mRNA expression of Bax. The correlation for the E2 level and mRNA expression of Bax, meaningfully, was positive (0.506) and statistically significant (p = 0.0001). A scatterplot indicated that there was a linear relationship between the E2 level and mRNA expression of Bax (Fig. 3b), while the variables were not approximately normally distributed and have some outliers (Fig. 3a).

a Simple scatter plot for the correlations of E2 and XIAP. The variables were not approximately normally distributed and have some outliers. b Simple scatter plot for the correlations of E2 and Bax. There was a linear relationship between the E2 level and mRNA expression of Bax
Discussion
In this study, the level of serum E2 was significantly increased in the PEMF group compared to that in the OVX group. The results of our investigation also showed that chondrocytes of the knee joint underwent pathological changes in OVX rats. After intervention with oestrogen, the chondrocyte lesions were significantly reduced. Thus, our data supports the view that low levels of E2 are associated with the development of OA. We presume that E2 inhibits pathological changes in chondrocytes, thus delaying the development of OA. We also found that exposure of OVX rats to PEMF for 30 days increased serum E2 levels compared to controls, and that such exposure resulted in a protective effect against histopathological changes similar to that seen in the control group that received E2 intervention. This demonstrated that PEMF enhanced serum E2 levels in OVX rats, suppressed the degeneration of cartilage, and had E2-like effects.
Our findings that bilateral OVX leads to OA through reduction in serum E2 levels and that PEMF affects the morphology of chondrocytes are consistent with those of a previous study [12]. Our preliminary study [12] suggested that PEMFs have systemic effects on oestrogen metabolism in ovariectomised rats. The study also highlights novel features important for understanding the mechanism of PEMF effects on the treatment of postmenopausal OA, since it shows for the first time that an effective stimulus for OVX rats of postmenopausal OA occurs through mediation of the intrinsic apoptosis signalling pathway.
Before age 50 years, the incidence of OA is much lower among women than among men. Around the age of 50, the prevalence of OA increases dramatically in women [22]. This suggests that reduced oestrogen levels present a risk for the development of OA. Series studies have found that decreasing E2 levels may result in OA and may be related to the oestrogen receptor (ER) and to gene polymorphisms of ER [23–27].
In this work, we examined the effect of PEMF exposure on ovariectomised rats by monitoring the expression of mRNA for the anti-apoptotic and apoptotic proteins XIAP and Bax. We also for the first time probed into the association between the E2 level and the mRNA expression of XIAP and Bax. In our previous study, we demonstrated that PEMF, as well as the administration of oestrogen, may inhibit the apoptosis of chondrocytes in OVX rats [12]. Our study supports previous work by Luo et al. indicating that PEMF has a systemic effect on oestrogen metabolism in ovariectomised rats. Since PEMF modulated apoptosis in our study, the data suggests that PEMF modulated the cartilage apoptosis signalling pathway by up-regulating XIAP mRNA and down-regulating Bax mRNA expression.
Inhibitor of apoptosis protein (IAP) belongs to a family of caspase inhibitors that prevent apoptosis at inappropriate times by inhibiting mature caspases [28]. The prototypical IAP, XIAP, inhibits caspase-9 and caspase-3 via specific binding regions that interact with mature caspases [28]. IAP proteins have been confirmed to protect diverse cells from programmed cell death in response to a variety of stimuli [29].
Bax is a member of the same family as the proapoptosis protein Bcl-2, a cytosolic protein that inserts into the mitochondrial membrane upon induction of apoptosis. Bax is found in many tissues of the body [30] and promotes apoptosis in a wide variety of cell types subjected to death stimuli; it plays an important role in diverse physiological events including spermatogenesis, anti-viral defense, and tumourigenesis [31].
Apoptosis signalling pathways are tightly regulated by both pro- and anti-apoptotic mechanisms to allow robustness of the system. Data presented in this paper clearly show up-regulation of XIAP and down-regulation of Bax in chondrocytes in response to PEMF stimulation, suggesting that PEMF effectively stimulates postmenopausal OA rats by regulating the relevant apoptotic proteins. The observed changes in the expression levels of XIAP and Bax mRNA in PEMF-treated animals demonstrate that changes in the expression levels of these proteins are potential targets of the electromagnetic fields. The role of these two proteins requires further investigation, however.
Apoptosis of chondrocytes is a major contributing factor to the development of arthritis. We propose that in older females, the exposure of chondrocytes to low oestrogen levels may lead to chondrocyte apoptosis and cartilage degradation. Our study has important implications for the use of PEMF in the prevention of diseases, such as arthritis, that damage cartilage. Finally, we found there was a linear relationship between E2 level and the expression level of Bax mRNA. This highlighted the association between oestrogen and chondrocyte apoptosis related to the aetiology OA development. Nevertheless, the results showed the negative correlation of E2 level and the expression level of XIAP mRNA. Whether different intensity and frequency of PEMF will have different effects on E2 levels and chondrocyte apoptosis needs to be further investigated.
In conclusion, low E2 levels result in chondrocyte pathology and in down-regulated XIAP expression and up-regulated Bax expression. PEMF can reduce cartilage degeneration induced by decreasing E2 levels through regulation of relevant apoptotic proteins, can significantly enhance the level of serum E2 in OVX rats, and can up-regulate the expression of XIAP and down-regulate the expression of Bax. The results of our study suggest that regulation of the expression of apoptotic proteins may be one of the mechanisms by which PEMF is effective in treatment of OA disease and that apoptosis may play a significant role in the physiopathology of articular cartilage in the course of OA. The signal pathways involved in this mechanism should be addressed. If the relevant mechanisms can be identified, PEMF may be a new and attractive target for OA treatment. Further studies are necessary to confirm the ER as a protective factor for OA cartilage and to address the potential correlation between ER and IAP proteins.
References
Nevitt MC, Felson DT (1996) The epidemiology of osteoarthritis in the peripheral joints: sex hormones and the risk of osteoarthritis in women: epidemiological evidence. Ann Rheum Dis 55:673–676
Cecil RL, Archer BH (1992) Classification and treatment of chronic arthritis. JAMA 87:741–746
Nevitt MC, Cummings SR, Lane NE et al (1996) Association of estrogen replacement therapy with the risk of osteoarthritis of the hip in elderly white women. Study of osteoporotic fractures research group. Arch Intern Med 156:2073–2080
Lawrence JS, Bremner JM, Bier F (1996) Osteo-arthrosis. Prevalence in the population and relationship between symptoms and X-ray changes. Ann Rheum Dis 25:1–24
Hoegh-Andersen P, Tanko LB, Andersen TL et al (2004) Ovariectomized rats as a model of postmenopausal osteoarthritis: validation and application. Arthritis Res Ther 6:R169–R180
Dai G, Wang S, Li J et al (2006) The validity of osteoarthritis model induced by bilateral ovariectomy in guinea pig. J Huazhong Univ Sci Technolog Med Sci 26:716–719
Bassett CAL, Payluk RJ, Pilla AA (1974) Augmentation of bone repair by inductively coupled electromagnetic fields. Science 184:575–577
Markov MS (2007) Pulsed electromagnetic field therapy history, state of the art and future. Environmentalist 27:465–475
Fioravanti A, Nerucci F, Collodel G et al (2002) Biochemical and morphological study of human articular chondrocytes cultivated in the presence of pulsed signal therapy. Ann Rheum Dis 61:1032–1033
De Mattei M, Pasello M, Pellati A et al (2003) Effects of electromagnetic fields on proteoglycan metabolism of bovine articular cartilage explants. Connect Tissue Res 44:54–59
De Mattei M, Pellati A, Pasello M et al (2004) Effects of physical stimulation with electromagnetic field and insulin growth factor-I treatment on proteoglycan sysnthesis of bovine articular cartilage. Osteoarthritis Cartilage 12:793–800
Luo Q, Li S, He C et al (2009) Pulse electromagnetic fields effects on serum E2 levels, chondrocyte apoptosis, and matrix metalloproteinase-13 expression in ovariectomized rats. Rheumatol Int 29:927–935
Reed JC (2000) Mechanisms of apoptosis. Am J Pathol 157:1415–1430
Thornberry NA, Lazebnik Y (1998) Caspases: enemies within. Science 281:1312–1316
Salvesen GS, Abrams JM et al (2004) Caspase activation stepping on the gas or releasing the brakes? Lessons from humans and flies. Oncogene 23:2774–2784
Miller LK (1999) An exegesis of IAPs: salvation and surprises from BIR motifs. Trends Cell Biol 9:323–328
Eckelman BP, Salvesen GS (2006) The human anti-apoptotic proteins cIAP1 and cIAP2 bind but do not inhibit caspases. J Biol Chem 281:3254–3260
Deveraux QL, Roy N, Stennicke HR et al (1998) IAPs block apoptotic events induced by caspase-8 and cytochrome c by direct inhibition of distinct caspases. EMBO J 17:2215–2223
Smaili SS, Hsu YT, Sanders KM et al (2001) Bax translocation to mitochondria subsequent to a rapid loss of mitochondrial membrane potential. Cell Death Differ 8:909–920
Hoegh-Andersen P, Tanko LB, Andersen TL et al (2004) Ovariectomized rats as a model of postmenopausal osteoarthritis: validation and application. Arthritis Res Ther 6:R169–R180
Pritzker KPH, Gay S, Jimenez SA et al (1995) Osteoarthritis cartilage histopathology: grading and staging. Osteoarthritis Cartilage 14:13–29
Oliveria SA, Felson DT, Reed JI et al (1995) Incidence of symptomatic hand, hip and knee osteoarthritis among patients in a health maintenance organisation. Arthritis Rheum 38:1134–1141
Oshima Y, Matsuda K, Yoshida A et al (2007) Localization of estrogen receptors alpha and beta in the articular surface of the rat femur. Acta Histochem Cytochem 40:27–34
Kinney RC, Schwartz Z, Week K et al (2005) Human articular chondrocytes exhibit sexual dimorphism in their responses to 17 beta-estradiol. Osteoarthr Cartil 13:330–337
Bergink AP, van Meurs JB, Loughlin J et al (2003) Estrogen receptor alpha gene haplotype is associated with radiographic osteoarthritis of the knee in elderly men and women. Arthritis Rheum 48:1913–1922
Lian K, Lui L, Zmuda JM et al (2007) Estrogen receptor alpha genotype is associated with a reduced prevalence of radiographic hip osteoarthritis in elderly Caucasian women. Osteoarthr Cartil 15:972–978
Fytili P, Giannatou E, Papanikolaou V et al (2005) Association of repeat polymorphisms in the estrogen receptors alpha, beta, and androgen receptor genes with knee osteoarthritis. Clin Genet 68:268–277
Deveraux QL, Roy N, Stennicke HR et al (1998) IAPs block apoptotic events induced by caspase-8 and cytochrome c by direct inhibition of distinct caspases. EMBO J 17:2215–2223
Nicholson DW (2001) Apoptosis. Baiting death inhibitors. Nature 410:33–34
Krajewski S, Krajewski M, Shabaik A et al (1994) Immunohistochemical determination of in vivo distribution of Bax, a dominant inhibitor of Bcl-2. Am J Path 145:1323–1336
Yang E, Korsmeyer SJ (1996) Molecular apoptosis: a discourse on the Bcl2 family and cell death. Blood 188:386–401
Acknowledgements
This research was supported by the National Natural Science Foundation of China (Grant No. 30672215). The authors acknowledge support from the Key Lab of Molecular Research, West China Hospital/West China Medical School of Sichuan University. The authors are also most grateful to Dr. Deying Kang for his statistical analysis assistance.
Declaration of interest
The authors report no conflicts of interest. The authors alone are responsible for the content and writing of this article.
Author information
Authors and Affiliations
Corresponding author
Additional information
Shasha Li and Qinglu Luo contributed equally to this work.
Rights and permissions
About this article
Cite this article
Li, S., Luo, Q., Huang, L. et al. Effects of pulsed electromagnetic fields on cartilage apoptosis signalling pathways in ovariectomised rats. International Orthopaedics (SICOT) 35, 1875–1882 (2011). https://doi.org/10.1007/s00264-011-1245-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00264-011-1245-3