
Abstract
As many plant secondary metabolites have antimicrobial activity, microorganisms of the anaerobic digestion process might be affected when plant material rich in these compounds is digested. Hitherto, the effects of plant secondary metabolites on the anaerobic digestion process are poorly investigated. In this study, the alkaloid gramine, a constituent of reed canary grass, was added daily to a continuous co-digestion of grass silage and cow manure. A transient decrease of the methane yield by 17 % and a subsequent recovery was observed, but no effect on other process parameters. When gramine was infrequently spiked in higher amounts, the observed inhibitory effect was even more pronounced including a 53 % decrease of the methane yield and an increase of acetic acid concentrations up to 96 mM. However, the process recovered and the process parameters were finally at initial values (methane yield around 255 LN CH4 per gram volatile solids of substrate and acetic acid concentration lower than 2 mM). The bacterial communities of the reactors remained stable upon gramine addition. In contrast, the methanogenic community changed from a well-balanced mixture of five phylotypes towards a strong dominance of Methanosarcina (more than two thirds of the methanogenic community) while Methanosaeta disappeared. Batch inhibition assays revealed that acetic acid was only converted to methane via acetoclastic methanogenesis which was more strongly affected by gramine than hydrogenotrophic methanogenesis and acetogenesis. Hence, when acetoclastic methanogenesis is the dominant pathway, a shift of the methanogenic community is necessary to digest gramine-rich plant material.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Many plants form secondary metabolites have ecological and physiological functions like regulation of growth, activity against herbivores, fungi, and prokaryotes as well as allelopathy. Plant secondary metabolites (PSM) are classified according to their chemical structures, e.g., phenols, alkaloids, or terpenoids (Harborne 1999). If plant biomass containing secondary metabolites is considered for energy generation like biogas production, the effects of these PSM on the anaerobic digestion (AD) process should be taken into account. Many PSM have antimicrobial activity (Wallace 2004) and may thus affect microorganisms of the biogas process.
The effects of PSM on AD processes are poorly studied. Recently, the phenolic PSM coumarin was proven to cause a reduction of methane formation and to induce changes of the bacterial community composition while the methanogenic community remained stable (Popp et al. 2015). Mono- and polyphenols were shown to have an inhibitory effect on methane production during anaerobic treatment of wastewater (Akassou et al. 2010; Borja et al. 1997). In contrast, two polyphenolic flavonoids, rutin and hesperidin, commonly found in brewery wastewater had no effect on AD (Herrmann and Janke 2001). Furthermore, polyphenolic tannins are known to inhibit anaerobic wastewater treatment processes by complexation of proteins (Mannucci et al. 2010). Effects on ruminal methane production were investigated by screening of hundreds of PSMs (Bodas et al. 2008; García-González et al. 2008). The motivation of this research, however, was the reduction of methane emissions from ruminal livestock. In contrast, maximized methane yields are desired for biogas processes.
Other inhibitors of the AD process like antibiotics, nanoparticles, long-chain fatty acids, and xenobiotics occurring in waste water were studied in more detail (Gonzalez-Estrella et al. 2013; Flores et al. 2015; Ma et al. 2015). Systematic studies with several antibiotics at different concentrations revealed that the volatile fatty acids (VFA) degrading pathways were affected (Cetecioglu et al. 2012; Aydin et al. 2015). Furthermore, recent investigations shed light on the interplay of antibiotics, the microbiome, and their biodegradation capabilities (Aydin et al. 2016). However, the effect of inhibitors on biogas production from agricultural and municipal sources might be different because of the different process conditions.
Perennial grasses as substrates for biogas production can provide high methane yields while entailing ecological advantages over established energy crops like maize. Cultivation of perennial grasses like reed canary grass (Phalaris arundinacaea) decreases soil erosion, sequesters carbon, and improves biological diversity by providing habitat for wildlife (Johnson et al. 2007). Reed canary grass is commonly found in temperate regions in Europe, North America, and Asia. It grows on moist and wet sites but also tolerates drought (Casler et al. 2009). Reed canary grass is not suitable for grazing or as fodder as it contains harmful alkaloids such as gramine, hordenine, and N,N-dimethyltryptamine, which reduce its palatability. Sheep grazing on alkaloid-rich reed canary grass pastures shows a reduced voluntary intake and weight gain and a higher rate of diarrhea (Marten et al. 1976). However, there is an increasing interest in using reed canary grass for other purposes than animal feed like energy production (Jasinskas et al. 2008; Oleszek et al. 2014).
Gramine (1-(1H-indol-3-yl)-N,N-dimethylmethanamine) is the simplest indole alkaloid commonly found in reed canary grass. Its concentration amounts up to 0.7 % of total solid (TS) content (Coulman et al. 1976). Conflicting results were reported for the effect of gramine in ruminal systems. Coulman et al. (1977) found only little effect of gramine on the in vitro digestibility of cellulose or plant material. In contrast, gramine inhibited the in vitro digestibility of orchard grass (G.W. Arnold, personal communication cited in Coulman et al. (1977)). Furthermore, gramine was proven to have antibacterial properties impairing the growth of Pseudomonas (Sepulveda and Corcuera 1990). To the best of our knowledge, the present study is the first addressing the effects of gramine on AD.
Gramine was added continuously or episodically to lab-scale continuous co-digestions of cow manure and grass silage. Effects on the process parameters were evaluated and bacterial as well as methanogenic communities were monitored by molecular fingerprinting on the DNA level. Furthermore, the inhibitory effects of gramine on acetogenesis and methanogenesis were investigated in more detail by batch inhibition assays. Acetic acid conversion pathways were distinguished by using 13C-methyl group-labeled acetic acid. In contrast to existing studies, no presumption about dominant pathways or the most sensitive step of the AD process was applied in the inhibition studies.
Methods
Operation of continuous stirred tank reactors
Three continuous stirred tank reactors (CSTRs) (named A, B, and C; total volume 15 L, working volume 10 L) were operated in parallel under mesophilic conditions (38 °C). The reactors were inoculated with a mixture of 7 L reactor content from a lab-scale biogas reactor digesting cow manure and grass silage and 23 L digestate from a pilot-scale biogas reactor digesting cow manure and maize silage. A mixture of 58 g grass silage (TS = 30 %fresh mass, volatile solids (VS) = 89 %TS) and 217–280 g cow manure (TS = 7–9 %fresh mass, VS = 77–80 %TS) was added daily to each reactor together with 0.04 g Ferrosorb DGμ (HeGo Biotec GmbH, Germany) per added grams of TS. Substrates were obtained from a farm-scale biogas plant near Neichen (Germany) and stored at 4 °C. Grass silage was milled in liquid nitrogen before adding to each reactor. Cow manure was diluted with tap water to obtain a TS content of 4.5 %fresh mass. The organic loading rate (OLR) was 3.2 gVS L−1 day−1, and the mean hydraulic retention time (HRT) was 21 days. A pre-experimental time of 80 days preceded the start of the gramine addition. The main experiment began with the start of the daily addition of 0.65 mmol of gramine (0.7 %TS of the grass silage) to reactor A. Reactor B was spiked with higher concentrations of gramine adding 13.8 mmol on day 63 (resulting in a theoretical concentration of 1.38 mM in the reactor), 27.5 mmol on day 86 (2.75 mM), and 27.5 mmol on day 121 (2.75 mM). Actual gramine concentrations after the second and third spike could be higher as the reactor might still contain gramine from the previous spike. The third reactor C served as a gramine-free control.
Analyses of the process parameters
TS and VS contents were determined as described by Sträuber et al. (2012). TS content of grass silage was corrected for volatile components as described by Popp et al. (2015). Daily gas production was measured by drum-type gas meters TG05 (Ritter, Germany) and normalized to standard pressure (101.325 kPa) and standard temperature (273.15 K). Gas composition was determined by an AWIFLEX gas analyzer (AWITE Bioenergie, Germany). pH values of digestates were measured by a pH 3310 pH meter equipped with a SenTix 41 electrode (WTW, Germany). Ammonia nitrogen concentration, organic acid/alkalinity ratio, and concentrations of VFA (acetic, propionic, iso-butyric, n-butyric, iso-valeric, and n-valeric acid) were determined as described by Lv et al. (2014).
Microbial community analysis by terminal restriction fragment length polymorphism fingerprinting
Samples of digestates were sieved (mesh size 1 mm) and frozen at −20 °C until DNA extraction. DNA was isolated from 200 μL of each sample with NucleoSpin Soil Kit (Macherey-Nagel, Germany) using lysis buffer SL1 and Enhancer SX according to the manufacturer’s instructions. Bacterial 16S ribosomal RNA (rRNA) genes were amplified by polymerase chain reaction (PCR) using the MyTaq™ Mix (Bioline, Germany) and applying the primers 27f and 1492r (Lane 1991). The PCR reaction mixtures contained 6.25 μL of MyTaq™ Mix, 1 μL of each primer (5 pmol), and 1 μL genomic template DNA in a total volume of 12.5 μL. The cycling protocol comprised an initial denaturation at 95 °C for 1 min, followed by 30 cycles of denaturation at 95 °C for 15 s, primer annealing at 58 °C for 15 s, elongation at 72 °C for 10 s, and a final elongation step at 72 °C for 15 min. The methanogenic community was investigated by amplifying the mcrA (methyl coenzyme-M reductase) genes using the same polymerase and the primers mlas and mcrA-rev described by Steinberg and Regan (2008). The PCR reaction mixtures contained 6.25 μL of MyTaq™ Mix, 1 μL of each primer (5 pmol), and 1 μL of genomic template DNA in a total volume of 12.5 μL. The thermocycling protocol was as follows: 3 min at 95 °C, followed by 5 cycles for 30 s at 95 °C, 45 s at 48 °C, a temperature ramp from 48 °C to 72 °C at 0.1 K s−1 and 30 s at 72 °C, followed by 26 cycles at 95 °C for 30 s, 45 s at 52 °C, 120 s at 72 °C, and a final elongation step at 72 °C for 20 min. The primers 27f and mcrA-rev were labeled with phosphoramidite fluorochrome 5-carboxyfluorescein (FAM). For analyzing terminal restriction fragment length polymorphism (T-RFLP), restriction of 16S rRNA amplicons with HaeIII or MspI (New England Biolabs, Germany) and of mcrA amplicons with MwoI or BstNI (New England Biolabs, Germany) was performed as described earlier (Popp et al. 2015). For T-RFLP analysis of 16S rRNA amplicons, 0.2 μL of the size standard MapMarker 1000 labeled with carboxy-X-rodamine (ROX; Eurogentec, Germany) was added; for analyzing the mcrA amplicons, 0.2 μL of the MCLAB Red DNA Size Standard (Nimagen, Germany) was added. The terminal restriction fragments (T-RFs) were separated by an ABI PRISM 3130xl Genetic Analyzer (Applied Biosystems, USA). T-RFLP data were analyzed as described by Popp et al. (2015). Relative abundances of T-RFs were calculated by dividing the individual peak areas by the total peak area. Phylogenetic assignment of T-RFs based on mcrA amplicons was performed using the public mcrA T-RFLP database (Bühligen et al. 2016). Differences between the reactors regarding the 16S rRNA gene-based T-RF profiles from all sampling days (days 0, 49, 84, 119, and 148, respectively) were analyzed by analysis of similarities (ANOSIM) function of the “vegan” package (version 2.3-3) using R version 3.2.3 (Oksanen et al. 2016).
Batch inhibition experiments
Inhibition of acetogenesis and methanogenesis by gramine was further investigated in batch experiments. Therefore, gramine and specific substrates for acetogenesis (propionic or n-butyric acid) and methanogenesis (acetic acid or H2/CO2) were added to digestate from reactors B or C. Reactors B and C were further operated after the main experiment as described above. Reactor B was further spiked with gramine (27.5 mmol) on days 150, 170, 184, 196, 228, and 252. Digestates from reactors B and C were taken as inocula on day 214 for acetic acid, on day 263 for hydrogen, and on day 266 for propionic and n-butyric acid experiments. Digestate was stored under N2 atmosphere at 37 °C for at least 3 days to minimize background gas production. Afterwards, it was sieved (1 mm mesh size) and 50 g of it were filled in 120-mL cultivation bottles under N2 flow. Bottles were closed with rubber stoppers and sealed with aluminum caps, and headspaces were flushed with N2/CO2 (80:20 vol/vol) for at least 2 min (0.2 L min−1). Gramine (0.29 mmol) had been added beforehand to the bottles resulting in a final concentration of 5.7 mM. As substrates, 13C-labeled acetic acid (sodium acetate-2-13C, 10 mM, Sigma-Aldrich, Germany), propionic acid (10 mM), or n-butyric acid (2 mM) were added from anoxic stock solutions (sodium salts). Substrate concentrations were based on literature values and preliminary experiments performed in our laboratory to ensure complete substrate utilization and to avoid inhibition by high substrate concentrations (data not shown). H2/CO2 (80:20 vol/vol) gas was added to 1 bar overpressure. Duplicate cultivation bottles were incubated at 37 °C.
VFA concentrations in the liquid phase were monitored by high-pressure liquid chromatography (HPLC) as described by Popp et al. (2015). Gas production was measured by using a liquid displacement technique as described by Porsch et al. (2015). Gas volumes were normalized to standard temperature (273.15 K) and pressure (101.325 kPa) and corrected for background gas production using substrate-free controls. Gas phase of cultivation bottles was sampled to determine the gas composition by taking 1 mL of gas with a syringe flushed with nitrogen. Gas samples were transferred into 20-mL vials filled with argon for gas chromatography (GC). Samples were pre-heated to 40 °C by a TurboMatrix HS-110 autosampler (PerkinElmer, Germany). Gas composition was measured by a gas chromatograph Clarus 580 (PerkinElmer, Germany) equipped with parallel columns. A Hayesep N column (7″ length, 1/8″ diameter) separated CO2 and a Mole sieve 13X column (9″ length, 1/8″ diameter) separated N2, H2 and CH4. Argon was used as carrier gas with a flow rate of 25 mL min−1 at a pressure of 230 kPa. Injector temperature was 150 °C and detector temperature was 200 °C. For analysis of 12CO2 and 13CO2 as well as 12CH4 and 13CH4 in the gas phase, gas chromatography-mass spectrometry (GC-MS) analysis was performed using a PerkinElmer GC Clarus 600 system with an Rtx®-1 capillary column (60 m × 320 μM). For GC-MS detection, an electron ionization system was operated with an ionization energy of 70 eV. Mass spectra were taken from 14 to 70 Da. Helium was used as carrier gas at a constant flow with 300 kPa, and an injection volume of 10 μL (split ratio 10:1) was employed manually using gas-tight syringes. Each sample was measured five times and average values were calculated. The total amounts of 12CO2 and 13CO2 as well as 12CH4 and 13CH4 were analyzed by extraction of the masses 44 and 45 as well as 16 and 17, respectively, followed by peak integration. Finally, the percentage of labeled CO2 and CH4 was calculated and corrected for background 13CO2 and 13CH4 contents by measuring the gas phase of a substrate-free control.
Results
Impact of gramine on a continuous biogas process
Continuous AD experiments were performed to investigate the effect of gramine. Therefore, three CSTRs A, B, and C digesting grass silage and cow manure were operated in parallel for 80 days at a constant OLR to obtain a stable biogas process. The biogas yield of each of the three reactors was around 445 mLN gVS −1 and the methane content about 60 % resulting in a methane yield of 267 mLN gVS −1. VFA concentrations in all reactors were lower than 3.2 mM acetic acid, 0.3 mM propionic acid, 68 μM iso-butyric acid, 28 μM n-butyric acid, 23 μM iso-valeric acid, and 6.1 μM n-valeric acid. Ammonia nitrogen concentrations were between 1.0 and 1.6 g L−1 and always similar in all reactors. pH values were around 7.5 in all reactors.
The impact of gramine on the biogas process was first tested by adding 0.65 mmol gramine (0.7 %TS of grass silage) daily to reactor A for 150 days until the end of the main experiment. Based on the HRT of 21 days, the gramine concentration would increase up to 1.38 mM. The process parameters were compared to those of reactor C which served as gramine-free control. However, none of the monitored process parameters (biogas yield, VFA concentrations, pH value, and ammonia nitrogen concentration) diverged from the parameters of the gramine-free control reactor except for the gas composition. The methane content of the biogas decreased from 60 % on day 20 to 54 % on day 30. Simultaneously, the carbon dioxide content increased from 39 to 46 %. Hence, a 17 % decrease of the methane yield was observed. However, the biogas composition returned to the initial ratio of about 60 % methane and 40 % carbon dioxide on day 38.
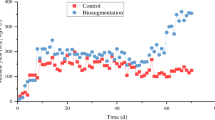
For further investigations, the impact of gramine on the biogas process was studied by adding higher gramine doses (13.8 mmol) to reactor B on day 63 resulting in a concentration of 1.38 mM gramine in the reactor content. Before, no gramine was added to reactor B. This spike of gramine had no effect on the process parameters (Fig. 1). A second spike of 27.5 mmol gramine was applied to reactor B on day 86 (2.75 mM). A decrease of the biogas yield was observed from day 89 on Fig. 1a. This decline continued until day 97 to a biogas yield of 262 mLN gVS −1. Simultaneously, the methane content decreased from 60 % to a minimum of 45 % while the carbon dioxide content increased to 55 %. This corresponded to a methane yield of 126 mLN gVS −1 which was about 53 % less than that of the gramine-free control reactor C. Furthermore, a drop of the pH value was observed from around 7.5 to below 7.2 around day 100 (Fig. 1b). In contrast, the pH values in the gramine-free reactor C remained stable at 7.5. Meanwhile, VFA concentrations increased in the gramine-spiked reactor B (Fig. 1c), whereas concentrations in the gramine-free control reactor C did not change. The concentration of acetic acid increased to a maximum of 96 mM on day 101. The concentration of other VFAs increased to a maximum of 3.9 mM propionic acid, 1.8 mM iso-butyric acid, 2.1 mM n-butyric acid, 1.8 mM iso-valeric acid and 0.1 mM n-valeric acid. Concentrations of these acids in the gramine-free control reactor C were lower than the detection limits of 4.2 μM for iso-butyric acid, 9.6 μM for n-butyric acid, 3.8 μM for iso-valeric acid, and 6.1 μM for n-valeric acid. Ammonia nitrogen concentrations remained between 1.0 and 1.6 g L−1 after gramine addition and were similar to those in the gramine-free control reactor C. The observed decreases of the biogas yield and pH and the increase of VFA concentrations indicated a transient process inhibition due to gramine before the biogas process recovered.

Process parameters of the co-digestion of grass silage and cow manure in the gramine-exposed reactors A and B and the gramine-free control reactor C between day 60 and 130. Gramine was added daily to reactor A and was spiked to reactor B on days 63, 86, and 121 as indicated by arrows. a Biogas yield as well as b pH value of all three reactors and c VFA concentrations of the gramine-spiked reactor B
The biogas yield increased to a maximum of 626 mLN gVS −1 on day 104, which was even more than the biogas yield of the gramine-free control reactor C (419 mLN gVS −1). Afterwards the biogas yield decreased to around 425 mLN gVS −1 which was similar to the biogas yield before the gramine addition and of the gramine-free reactor C. Likewise, the biogas composition returned to initial percentages of 60 % methane and 40 % carbon dioxide. Concentrations of VFAs also recovered to pre-inhibition values and those of control reactor C. Likewise, the pH value returned to the initial level of around 7.5. After this recovery, a third spike of gramine (27.5 mmol, resulting in a gramine concentration increase by 2.75 mM in the reactor) was applied on day 120 to reactor B. An 11 % decreased biogas yield compared to the gramine-free reactor was observed between day 123 and 129 (Fig. 1a) without any effect on the biogas composition. Thereafter, the biogas yield was as high as for the gramine-free reactor C. This third spike had no effect on the other monitored process parameters VFA concentrations, pH value, and ammonia nitrogen concentration.
Dynamics of bacterial communities
Along with the process parameters, the microbial communities in all three reactors were monitored. The bacterial communities were studied by T-RFLP based on the 16S rRNA genes. In total, 35 and 36 T-RFs were obtained by using restriction endonucleases HaeIII and MspI, respectively. Visual comparison of the T-RFLP profiles showed only slight differences between gramine-exposed reactors (A and B) and the gramine-free reactor C (see Fig. 2 for results based on HaeIII restriction and Figure S1 for results based on MspI). Furthermore, analysis of similarity (ANOSIM) was performed for each sampling day (days 0, 49, 84, 119, and 148, respectively). Only small, insignificant differences (ANOSIM R = 0.2, p = 0.06) were observed between the three reactors indicating that the continuous or episodic addition of gramine had no significant effect on bacterial community composition.

T-RFLP profiles based on bacterial 16S rRNA gene amplicons and HaeIII restriction endonuclease. T-RFLP profiles of the gramine-exposed reactors A and B and gramine-free control reactor C were compared at five different time points. Gramine was added daily to reactor A starting on day 0. Gramine was spiked to reactor B on days 63, 86, and 121
Dynamics of methanogenic communities
The dynamics of the methanogenic communities were monitored by T-RFLP fingerprinting based on amplicons of the functional mcrA marker genes. T-RFLP analyses using the restriction endonucleases MwoI (Fig. 3) and BstNI (Figure S2) showed 20 T-RFs each. Assigned T-RFs accounted for 83 to 95 % or 48 to 93 % of the community with MwoI or BstNI, respectively. Numerical data presented here refer to T-RFLP results from restriction with MwoI, which were confirmed by BstNI. Before gramine addition, the communities in all three reactors were mainly composed of five phylotypes: Methanoculleus (6–37 % relative abundance), Methanosaeta (12–27 %), Methanosarcina (5–24 %), Methanobacterium (14–23 %), and Methanomassiliicoccus (15–19 %), as well as other unassigned T-RFs with abundances lower than 6 %.

T-RFLP profiles based on mcrA amplicons and MwoI restriction endonuclease describing the methanogenic community structure. T-RFLP profiles of the gramine-exposed reactors A and B and of the gramine-free control reactor C were compared at five different time points. Gramine was added daily to reactor A starting on day 0. Gramine was spiked to reactor B on days 63, 86, and 121
The composition in reactor A changed after the daily addition of gramine (Fig. 3). On day 49, Methanosaeta could not be detected anymore and relative abundance of Methanosarcina rose to 36 %. Furthermore, Methanoculleus (37 %), Methanobacterium (10 %), and Methanomassiliicoccus (4 %) were found. Communities at the following three time points (days 84, 119, and 148) were quite similar. Methanosarcina was dominating with an average relative abundance (±standard deviation) of 67 % (±1 %) and Methanoculleus, Methanobacterium, and Methanomassiliicoccus were less abundant with 16 (±1 %), 9 (±1 %), and 8 % (±1 %), respectively.
The gramine addition on day 63 to reactor B resulted in a dominance of Methanoculleus (38 %) and a lower abundance of Methanosaeta (13 %), whereas Methanosarcina could not be detected (Fig. 3). The rest of the community remained as before gramine addition (Methanomassiliicoccus 19 % and Methanobacterium 16 %). After the second spike on day 86, a pronounced change in favor of Methanosarcina (72 %) was seen, whereas Methanoculleus and Methanobacterium became less abundant and Methanosaeta as well as Methanomassiliicoccus were not detected anymore. After the third spike on day 121, the dominance of Methanosarcina increased further to 79 %.
In contrast to reactors A and B, the methanogenic community of the gramine-free control reactor C remained stable (Fig. 3). In summary, the addition of gramine resulted in a strong dominance of Methanosarcina and reduced abundances of Methanoculleus, Methanobacterium and Methanomassiliicoccus, whereas Methanosaeta was not detected anymore.
Effects of gramine on acetogenesis and methanogenesis
As the biogas yield of reactor B decreased and the concentrations of VFA increased after gramine addition, gramine most probably adversely affected the acetogenesis and/or methanogenesis of the AD process. To gain more insight into this inhibition, batch tests were performed in which the specific substrates for acetogenesis or methanogenesis and gramine were added to digestate of the gramine-free control reactor C (non-adapted inoculum) or the gramine-spiked reactor B (adapted inoculum). The gramine concentration in these experiments was as high as 5.7 mM to induce strong inhibitory effects. Substrate consumption and methane formation (corrected for background production by subtracting methane production of substrate-free controls) were monitored and compared to the respective gramine-free control.
The effect of gramine on butyric acid degradation was tested by adding gramine and n-butyric acid. When the non-adapted inoculum was used, n-butyric acid was consumed within the first 7 days (Fig. 4a). Concentrations of acetic acid increased to 10 mM and the methane production was lower than in the substrate-free control. The same n-butyric acid consumption was observed in the gramine-free control in which, however, the acetic concentration decreased to 0.4 mM. Furthermore, 0.4 mmol methane was produced (Fig. 4a). When adding n-butyric acid and gramine to the adapted digestate, n-butyric acid was consumed within the first 7 days as well (Fig. 4b). However, in contrast to the non-adapted inoculum, the acetic acid concentration slightly decreased. Methane production was lower than in the substrate-free control. In the absence of gramine, n-butyric acid was readily degraded, acetic acid concentrations slightly decreased, and 0.2 mmol methane was produced.

Substrate consumption, acetic acid concentrations, and methane formation during anaerobic batch digestion of n-butyric or propionic acid with or without gramine. Digestate from the gramine-free control reactor C (non-adapted) and from the gramine-exposed reactor B (adapted to gramine) was used as inoculum. Mean values are depicted and error bars represent the range
When propionic acid was added to the non-adapted inoculum in the presence of gramine, it was degraded within the first 7 days (Fig. 4c). Acetic acid concentrations increased to 16 mM and less methane than in the substrate-free control was produced. The same propionic acid degradation was observed in the gramine-free control (Fig. 4c). In contrast, acetic acid concentrations remained stable around 0.4 mM, and 0.7 mmol methane was produced. With the adapted inoculum in the presence of gramine, propionic acid was degraded within 7 days (Fig. 4d). Acetic acid concentration was around 1.2 mM with decreasing tendency, and less methane than in the gramine-free controls was formed from propionic acid. In the gramine-free controls, kinetics of propionic and acetic acid concentrations were similar as with gramine (Fig. 4d) but more methane was produced. Taken together, n-butyric or propionic acid was readily degraded regardless of gramine presence and inoculum type. However, acetic acid accumulated in case of the non-adapted inoculum and gramine caused a decrease of the methane production irrespective of the inoculum type.
To evaluate gramine effects on acetoclastic methanogenesis, 13C-methyl group-labeled acetic acid was used as substrate. It allows differentiating between acetoclastic methanogenesis and syntrophic acetate oxidation with subsequent hydrogenotrophic methanogenesis as the two possible conversion routes of acetic acid. If only acetoclastic methanogenesis takes place, methane will be 13C-labeled and carbon dioxide remains unlabeled. When syntrophic acetate oxidation is involved, also carbon dioxide will be 13C-labeled. In all experiments described in the following, 13C-labeled methane was detected in up to 60 % of the total methane (Fig. 5a, b). In contrast, less than 1 % of the total carbon dioxide was 13C-labeled. Hence, the conversion occurred exclusively via acetoclastic methanogenesis irrespective of the presence of gramine.

Substrate consumption, methane formation, and acetic acid concentrations during anaerobic batch digestion of acetic acid or H2/CO2 as substrate with and without gramine. Digestates from the gramine-free control reactor C (non-adapted) and from the gramine-exposed reactor B (adapted to gramine) were used as inoculum. In the experiments with 13C-labeled acetic acid as substrate additionally, the percentage of labeled methane was determined. Mean values are depicted and error bars represent the range
When acetic acid was added as substrate to the non-adapted inoculum with gramine, its concentration initially decreased to 8 mM and then increased to 12 mM accompanied by lower methane production than in the substrate-free control (Fig. 5a). Without gramine, acetic acid decreased to 0.3 mM while 0.4 mmol methane was produced (Fig. 5a). When added to the adapted inoculum in the presence of gramine, acetic acid decreased from 11 to 4.9 mM (Fig. 5b). Until day 3, 0.13 mmol methane was formed. Thereafter, methane production was lower than in the substrate-free control. In contrast, the acetic acid concentrations of the gramine-free control steadily decreased to 1.1 mM while 0.3 mmol methane was produced. In summary, acetic acid accumulated and less methane was formed in the presence of gramine. However, acetic acid accumulation was less pronounced and methane production was higher with the adapted inoculum compared to the non-adapted inoculum.
H2/CO2 was added to investigate the effects of gramine on the hydrogenotrophic methanogenesis. When supplied to the non-adapted inoculum together with gramine, hydrogen was completely consumed within 4 days and 1 mmol of methane was produced (Fig. 5c). Hydrogen was similarly consumed in the gramine-free controls. However, less methane was produced (0.5 mmol). As with the non-adapted inoculum, in the presence of gramine, hydrogen was depleted by the adapted inoculum within 4 days and up to 0.2 mmol methane was produced (Fig. 5d). Even though the hydrogen consumption was similar, twice the amount of methane was produced in the gramine-free controls. In contrast to the non-adapted inoculum, a lower methane production was observed in the presence of gramine than without. In all cases, the acetic acid concentration remained around or below 1 mM.
Discussion
Whereas reed canary grass is not suitable as fodder, it is a promising alternative energy crop. Cultivation of reed canary grass is possible without pesticide or herbicide application and improves soil quality, and it can be harvested with common agricultural equipment (Strasil 2012). Furthermore, high potential methane yields of up to 430 LN CH4 kgVS −1 (Butkutė et al. 2014; Lehtomäki et al. 2008) enhanced the interest in cultivating reed canary grass for biogas production and thereby reducing social and economic conflicts faced with common energy crops (Mathews 2009). However, reed canary grass contains the alkaloid PSM gramine which might affect the AD process. Here, we described the effect of gramine on a continuous co-digestion of grass silage and cow manure.
Impact on continuous biogas production
Before gramine addition, the reactors yielded slightly more methane than comparable digestion processes described in the literature (Wang et al. 2010) which might be due to differences in substrate quality. When gramine was added daily, mimicking a gramine-rich crop as substrate for biogas production, no changes of process parameters except for a transient decrease of the methane yield were observed. However, when higher gramine doses were spiked, the decrease of the methane yield was more pronounced. Furthermore, VFA concentrations increased, with especially acetic acid concentrations up to 96 mM indicating a severe process imbalance. Hill et al. (1987) took acetic acid concentrations above 13 mM as indicator for process disturbance. Concentrations of propionic, butyric, and valeric acids also increased after gramine addition, but to a much smaller extent. Proposed thresholds for iso-butyric and iso-valeric acid indicating process disturbances (Hill and Holmberg 1988) were exceeded more than tenfold. Decreased methane production in combination with increased VFA concentrations thus clearly indicated a gramine-induced inhibition of the acetogenesis and/or methanogenesis of the AD process. This is a typical inhibition pattern which was also observed for other inhibitors like ammonia nitrogen, long-chain fatty acids and the plant secondary metabolite coumarin (Angelidaki and Ahring 1993; Nielsen and Ahring 2006; Popp et al. 2015).
Response of the microbial communities
The addition of gramine had little influence on the bacterial community composition. In the literature, changes of the bacterial community had been reported as a result of the addition of the PSM coumarin (Popp et al. 2015), along with increasing phenol concentrations (Rosenkranz et al. 2013) and when substrates were varied (Wang et al. 2009). Thus, it was somewhat surprising that the bacterial community was not susceptible to gramine.
In contrast, the methanogenic community composition changed towards a strong dominance of Methanosarcina and disappearance of Methanosaeta after gramine was added. The shift could be directly attributed to gramine addition and not to high VFA concentrations during the inhibition because the shift also occurred in the daily gramine-exposed CSTR A which showed no increased VFA concentrations. Methanosarcina is known for its resistance to stressors like pH or temperature changes and high VFA or ammonia concentrations (De Vrieze et al. 2012). According to our results, Methanosarcina appears to be also more resistant to the alkaloid gramine than other methanogens, especially Methanosaeta. However, at this point, we do not know how the differential sensitivity to gramine relates to biochemical or physiological differences between these methanogens. Furthermore, the way of gramine addition, continuously or episodically, apparently influenced the methanogenic community shift. Methanosaeta was not detected in the CSTR A after 49 days of daily gramine addition resulting in a theoretical gramine concentration of 1.23 mM. In contrast, Methanosaeta was still present in the CSTR B after the first spike of gramine which resulted in a gramine concentration of 1.36 mM. Hence, applying similar concentrations in different ways resulted in different communities.
The genera Methanosarcina and Methanosaeta are the only known acetoclastic methanogens. As Methanosaeta became undetectable after gramine addition and acetic acid was not converted via syntrophic acetate oxidation (as shown by the inhibition experiments with both inocula using 13C-labeled acetic acid), acetoclastic methanogenesis appeared to be performed only by Methanosarcina. Hence, acetoclastic methanogenesis became the bottleneck for acetic acid conversion to methane in the presence of gramine. Gramine addition with its effect on the functional redundancy of the methanogenic community is thus likely to cause a higher vulnerability of the AD process to process disturbances. Presumably, any further process imbalances compromising the activity of Methanosarcina would have put the AD process at risk and acetic acid might accumulate. In general, functional redundancy results in greater process stability (Briones and Raskin 2003) and a high (bacterial) diversity denotes the proper functioning of a biogas reactor (Carballa et al. 2011). Plants like reed canary grass contain usually a mixture of alkaloids including gramine (Duynisveld et al. 1990). It can be expected that other alkaloids also affect the AD, and even synergistic effects are possible as described for antibiotics and solvents in AD processes (Akyol et al. 2015; Aydin et al. 2015).
That the third spike of gramine to CSTR B had only a minor impact on the AD process might be explained by the shift to a methanogenic community that is better adapted to gramine. Alternatively, anaerobic degradation of gramine might have occurred decreasing its toxic effect. The anaerobic degradation of gramine was only observed in ruminal systems hitherto (Aguiar and Wink 2005) and is therefore also likely to happen in the biogas process.
Batch inhibition assays
From the results of the batch inhibition assays we concluded that mainly the acetoclastic methanogenesis was inhibited by gramine. This is in agreement with results from the CSTR experiments in which acetic acid was the most prominent VFA during the inhibition. The batch inhibition assays revealed that the conversion of n-butyric or propionic to acetic acid was not affected by gramine. However, methane production from n-butyric or propionic acid decreased as acetic acid accumulated. Unexpectedly, hydrogenotrophic methanogenesis of the non-adapted inoculum was actually positively influenced by gramine, a finding that could be confirmed by repeating the experiment with a similar setup (data not shown). This beneficial effect of gramine might be explained by a mechanism comparable to hormesis. Hormesis describes an overcompensating response to low levels of a toxin resulting in a higher fitness of the physiological system (Calabrese and Baldwin 2002). Hernandez and Edyvean (2008) observed a hormetic response of an AD process after addition of phenolic compounds. Experiments using different gramine concentrations and the resulting dose response curve would help to clarify if gramine has a hormetic effect on hydrogenotrophic methanogenesis.
Even though acetoclastic methanogenesis was inhibited by gramine, acetic acid was not converted via syntrophic acetate oxidation and subsequent hydrogenotrophic methanogenesis. However, the absence of this pathway was not due to gramine as acetate oxidation was observed neither with the non-adapted nor with the adapted inoculum. Hence, the syntrophic acetate oxidation was not possible due to unfavorable conditions or the absence of bacteria capable of acetate oxidation. This latter explanation is however implausible because reactors were supplied daily with cow manure likely containing syntrophic acetate oxidizing bacteria as they are commonly found in methanogenic environments (Hattori 2008).
Many studies investigating the effects of specific inhibitors on AD in sewage treatment focused on acetoclastic methanogenesis (Akyol et al. 2015; Cetecioglu et al. 2012; Hwu and Lettinga 1997). It has been argued that methanogenesis is the most sensitive step of AD and that 70 % of the methane comes from acetic acid via acetoclastic methanogenesis (Jeris and McCarty 1965). However, other steps of the AD process were shown to be more sensitive to specific inhibitors than acetoclastic methanogenesis (Hernandez and Edyvean 2008; Karri et al. 2006). Furthermore, hydrogenotrophic methanogenesis can also be the dominant methanogenic pathway in AD processes for biogas production (Nikolausz et al. 2013; Town et al. 2014). Few studies so far addressed acetoclastic and hydrogenotrophic methanogenesis by using acetic acid and hydrogen as substrate (Gonzalez-Estrella et al. 2013; Karri et al. 2006; Rodriguez-Freire et al. 2015). Furthermore, an even broader perspective was applied by investigating inhibitory effects on methanogenesis and on syntrophic butyric and propionic acid degradation using these compounds as substrates for acetogenesis (Aydin et al. 2015). Nonetheless, the alternative acetic acid conversion via syntrophic acetate oxidation with subsequent hydrogenotrophic methanogenesis is usually neglected. Here, syntrophic acetate oxidation was not studied directly but indirectly by using labeled acetic acid and measuring labeled methane and carbon dioxide.
Literature about effects of specific inhibitors on AD for biogas production is rare. Ma et al. (2015) described an inhibition of AD of algal biomass by long-chain fatty acids comprising analysis of process parameter as well as microbial communities and kinetic modeling. Similarly, the effects of phenol on anaerobic cellulose degradation and the bacterial as well as archaeal community were studied recently (Chapleur et al. 2015). Here, we described detailed inhibition experiments using CSTR processes (continuous addition and spiking of gramine) including monitoring of microbial communities and batch assays embracing the acetogenic as well as the methanogenic steps of the AD process. We suggest that inhibition studies in biogas systems should be performed without presumptions about preferred metabolic pathways or the most sensitive step. Only this unbiased approach will result in a comprehensive insight into the effects of specific inhibitors on the AD process.
References
Aguiar R, Wink M (2005) Do naive ruminants degrade alkaloids in the rumen? J Chem Ecol 31:761–787. doi:10.1007/s10886-005-3543-y
Akassou M, Kaanane A, Crolla A, Kinsley C (2010) Statistical modelling of the impact of some polyphenols on the efficiency of anaerobic digestion and the co-digestion of the wine distillery wastewater with dairy cattle manure and cheese whey. Water Sci Technol 62:475–483. doi:10.2166/wst.2010.235
Akyol Ç, Ince O, Coban H, Koksel G, Cetecioglu Z, Ayman Oz N, Ince B (2015) Individual and combined inhibitory effects of methanol and toluene on acetyl-CoA synthetase expression level of acetoclastic methanogen, Methanosaeta concilii. Int Biodeter Biodegr 105:233–238. doi:10.1016/j.ibiod.2015.09.013
Angelidaki I, Ahring BK (1993) Thermophilic anaerobic digestion of livestock waste: the effect of ammonia. Appl Microbiol Biotechnol 38:560–564. doi:10.1007/BF00242955
Aydin S, Cetecioglu Z, Arikan O, Ince B, Ozbayram EG, Ince O (2015) Inhibitory effects of antibiotic combinations on syntrophic bacteria, homoacetogens and methanogens. Chemosphere 120:515–520. doi:10.1016/j.chemosphere.2014.09.045
Aydin S, Ince B, Ince O (2016) Assessment of anaerobic bacterial diversity and its effects on anaerobic system stability and the occurrence of antibiotic resistance genes. Bioresour Technol 207:332–338. doi:10.1016/j.biortech.2016.01.080
Bodas R, López S, Fernández M, García-González R, Rodríguez AB, Wallace RJ, González JS (2008) In vitro screening of the potential of numerous plant species as antimethanogenic feed additives for ruminants. Anim Feed Sci Tech 145:245–258. doi:10.1016/j.anifeedsci.2007.04.015
Borja R, Alba J, Banks CJ (1997) Impact of the main phenolic compounds of olive mill wastewater (OMW) on the kinetics of acetoclastic methanogenesis. Process Biochem 32:121–133. doi:10.1016/s0032-9592(96)00055-6
Briones A, Raskin L (2003) Diversity and dynamics of microbial communities in engineered environments and their implications for process stability. Curr Opin Biotechnol 14:270–276. doi:10.1016/s0958-1669(03)00065-x
Bühligen F, Lucas R, Nikolausz M, Kleinsteuber S (2016) A T-RFLP database for the rapid profiling of methanogenic communities in anaerobic digesters. Anaerobe 39:114–116. doi:10.1016/j.anaerobe.2016.03.013
Butkutė B, Lemežienė N, Kanapeckas J, Navickas K, Dabkevičius Z, Venslauskas K (2014) Cocksfoot, tall fescue and reed canary grass: dry matter yield, chemical composition and biomass convertibility to methane. Biomass Bioenerg 66:1–11. doi:10.1016/j.biombioe.2014.03.014
Calabrese EJ, Baldwin LA (2002) Defining hormesis. Hum Exp Toxicol 21:91–97. doi:10.1191/0960327102ht217oa
Carballa M, Smits M, Etchebehere C, Boon N, Verstraete W (2011) Correlations between molecular and operational parameters in continuous lab-scale anaerobic reactors. Appl Microbiol Biotechnol 89:303–314. doi:10.1007/s00253-010-2858-y
Casler MD, Cherney JH, Brummer EC (2009) Biomass yield of naturalized populations and cultivars of reed canary grass. Bioenergy Res 2:165–173. doi:10.1007/s12155-009-9043-0
Cetecioglu Z, Ince B, Orhon D, Ince O (2012) Acute inhibitory impact of antimicrobials on acetoclastic methanogenic activity. Bioresour Technol 114:109–116. doi:10.1016/j.biortech.2012.03.020
Chapleur O, Madigou C, Civade R, Rodolphe Y, Mazéas L, Bouchez T (2015) Increasing concentrations of phenol progressively affect anaerobic digestion of cellulose and associated microbial communities. Biodegradation 27:15–27. doi:10.1007/s10532-015-9751-4
Coulman BE, Clark KW, Woods DL (1977) Effects of selected reed canary grass alkaloids on in vitro digestibility. Can J Plant Sci 57:779–785
Coulman BE, Woods DL, Clark KW (1976) Identification of low alkaloid genotypes of reed canary grass. Can J Plant Sci 56:837–845
De Vrieze J, Hennebel T, Boon N, Verstraete W (2012) Methanosarcina: the rediscovered methanogen for heavy duty biomethanation. Bioresour Technol 112:1–9. doi:10.1016/j.biortech.2012.02.079
Duynisveld GW, Slominski BA, Wittenberg KM, Campbell LD (1990) Alkaloid content of reed canarygrass (Phalaris arundinaceae L.) as determined by gas-liquid chromatography. Can J Plant Sci 70:1097–1103
Flores GAE, Fotidis IA, Karakashev DB, Kjellberg K, Angelidaki I (2015) Effects of benzalkonium chloride, Proxel LV, P3 hypochloran, triton X-100 and DOWFAX 63 N10 on anaerobic digestion processes. Bioresour Technol 193:393–400. doi:10.1016/j.biortech.2015.06.125
García-González R, López S, Fernández M, Bodas R, González JS (2008) Screening the activity of plants and spices for decreasing ruminal methane production in vitro. Anim Feed Sci Tech 147:36–52. doi:10.1016/j.anifeedsci.2007.09.008
Gonzalez-Estrella J, Sierra-Alvarez R, Field JA (2013) Toxicity assessment of inorganic nanoparticles to acetoclastic and hydrogenotrophic methanogenic activity in anaerobic granular sludge. J Hazard Mater 260:278–285. doi:10.1016/j.jhazmat.2013.05.029
Harborne JB (1999) Classes and functions of secondary products from plants. In: Walton NJ, Brown, D.E. (ed) Chemicals from plants: perspectives on plant secondary products. World Scientific & Imperial College Press, London, pp 1–26
Hattori S (2008) Syntrophic acetate-oxidizing microbes in methanogenic environments. Microbes Environ 23:118–127. doi:10.1264/jsme2.23.118
Hernandez JE, Edyvean RGJ (2008) Inhibition of biogas production and biodegradability by substituted phenolic compounds in anaerobic sludge. J Hazard Mater 160:20–28. doi:10.1016/j.jhazmat.2008.02.075
Herrmann AP, Janke HD (2001) Cofermentation of rutin and hesperidin during two-stage anaerobic pre-treatment of high-loaded brewery wastewater. Water Res 35:2583–2588. doi:10.1016/s0043-1354(00)00575-3
Hill DT, Cobb SA, Bolte JP (1987) Using volatile fatty-acid relationships to predict anaerobic digester failure. Transactions of the ASAE 30:496–501
Hill DT, Holmberg RD (1988) Long-chain volatile fatty acid relationships in anaerobic digestion of swine waste. Biol Waste 23:195–214. doi:10.1016/0269-7483(88)90034-1
Hwu CS, Lettinga G (1997) Acute toxicity of oleate to acetate-utilizing methanogens in mesophilic and thermophilic anaerobic sludges. Enzyme Microb Tech 21:297–301. doi:10.1016/s0141-0229(97)00050-1
Jasinskas A, Zaltauskas A, Kryzeviciene A (2008) The investigation of growing and using of tall perennial grasses as energy crops. Biomass Bioenerg 32:981–987. doi:10.1016/j.biombioe.2008.01.025
Jeris JS, McCarty PL (1965) The biochemistry of methane fermentation using C14 tracers. Water Pollut Con F 37:178–192
Johnson JM-F, Coleman MD, Gesch R, Jaradat A, Mitchell R, Reicosky D, Wilhelm WW (2007) Biomass-bioenergy crops in the United States: a changing paradigm. Am J Plant Sci Bio 1:1–28
Karri S, Sierra-Alvarez R, Field JA (2006) Toxicity of copper to acetoclastic and hydrogenotrophic activities of methanogens and sulfate reducers in anaerobic sludge. Chemosphere 62:121–127. doi:10.1016/j.chemosphere.2005.04.016
Lane DJ (1991) 16S/23S rRNA sequencing. In: Stackebrandt E, M G (eds) nucleic acid techniques in bacterial systematics. John Wiley & Sons, Chichester, pp. 177–203
Lehtomäki A, Viinikainen TA, Rintala JA (2008) Screening boreal energy crops and crop residues for methane biofuel production. Biomass Bioenergy 32:541–550. doi:10.1016/j.biombioe.2007.11.013
Lv Z, Hu M, Harms H, Richnow HH, Liebetrau J, Nikolausz M (2014) Stable isotope composition of biogas allows early warning of complete process failure as a result of ammonia inhibition in anaerobic digesters. Bioresour Technol 167:251–259. doi:10.1016/j.biortech.2014.06.029
Ma J, Zhao Q-B, Laurens LLM, Jarvis EE, Nagle NJ, Chen S, Frear CS (2015) Mechanism, kinetics and microbiology of inhibition caused by long-chain fatty acids in anaerobic digestion of algal biomass. Biotechnol Biofuels 8:1–12. doi:10.1186/s13068-015-0322-z
Mannucci A, Munz G, Mori G, Lubello C (2010) Anaerobic treatment of vegetable tannery wastewaters: a review. Desalination 264:1–8. doi:10.1016/j.desal.2010.07.021
Marten GC, Jordan RM, Hovin AW (1976) Biological significance of reed canarygrass alkaloids and associated palatability variation to grazing sheep and cattle. Agron J 68:909–914. doi:10.2134/agronj1976.00021962006800060017x
Mathews JA (2009) From the petroeconomy to the bioeconomy: integrating bioenergy production with agricultural demands. Biofuel Bioprod Bior 3:613–632. doi:10.1002/bbb.181
Nielsen HB, Ahring BK (2006) Responses of the biogas process to pulses of oleate in reactors treating mixtures of cattle and pig manure. Biotechnol Bioeng 95:96–105. doi:10.1002/bit.20963
Nikolausz M, Walter RFH, Sträuber H, Liebetrau J, Schmidt T, Kleinsteuber S, Bratfisch F, Günther U, Richnow HH (2013) Evaluation of stable isotope fingerprinting techniques for the assessment of the predominant methanogenic pathways in anaerobic digesters. Appl Microbiol Biot 97:2251–2262. doi:10.1007/s00253-012-4657-0
Oksanen J, Blanchet FG, Kindt R, Legendre P, Minchin PR, O'Hara RB, Simpson GL, Solymos P, Henry M, Stevens H, Wagner H (2016) Multivariate analysis of ecological communities in R: vegan tutorial. http://cc.oulu.fi/~jarioksa/opetus/metodi/vegantutor.pdf Accessed 12.01.2016 2016
Oleszek M, Król A, Tys J, Matyka M, Kulik M (2014) Comparison of biogas production from wild and cultivated varieties of reed canary grass. Bioresour Technol 156:303–306. doi:10.1016/j.biortech.2014.01.055
Popp D, Schrader S, Kleinsteuber S, Harms H, Sträuber H (2015) Biogas production from coumarin-rich plants—inhibition by coumarin and recovery by adaptation of the bacterial community. FEMS Microbiol Ecol 91. doi:10.1093/femsec/fiv103
Porsch K, Wirth B, Tóth EM, Schattenberg F, Nikolausz M (2015) Characterization of wheat straw-degrading anaerobic alkali-tolerant mixed cultures from soda lake sediments by molecular and cultivation techniques. Microbial Biotechnol 8:801–814. doi:10.1111/1751-7915.12272
Rodriguez-Freire L, Moore SE, Sierra-Alvarez R, Field JA (2015) Adaptation of a methanogenic consortium to arsenite inhibition. Water Air Soil Poll 226:1–7. doi:10.1007/s11270-015-2672-3
Rosenkranz F, Cabrol L, Carballa M, Donoso-Bravo A, Cruz L, Ruiz-Filippi G, Chamy R, Lema JM (2013) Relationship between phenol degradation efficiency and microbial community structure in an anaerobic SBR. Water Res 47:6739–6749. doi:10.1016/j.watres.2013.09.004
Sepulveda BA, Corcuera LJ (1990) Effect of gramine on the susceptibility of barley leaves to Pseudomonas syringae. Phytochemistry 29:465–467. doi:10.1016/0031-9422(90)85098-z
Steinberg LM, Regan JM (2008) Phylogenetic comparison of the methanogenic communities from an acidic, oligotrophic fen and an anaerobic digester treating municipal wastewater sludge. Appl Environ Microbiol 74:6663–6671. doi:10.1128/AEM.00553-08
Strasil Z (2012) Evaluation of reed canary grass (Phalaris arundinacea L.) grown for energy use. Res Agr Eng 58:119–130
Sträuber H, Schröder M, Kleinsteuber S (2012) Metabolic and microbial community dynamics during the hydrolytic and acidogenic fermentation in a leach-bed process. Energ Sustain Soc 2:13. doi:10.1186/2192-0567-2-13
Town JR, Links MG, Fonstad TA, Dumonceaux TJ (2014) Molecular characterization of anaerobic digester microbial communities identifies microorganisms that correlate to reactor performance. Bioresour Technol 151:249–257. doi:10.1016/j.biortech.2013.10.070
Wallace RJ (2004) Antimicrobial properties of plant secondary metabolites. P Nutr Soc 63:621–629. doi:10.1079/pns2004393
Wang H, Lehtomäki A, Tolvanen K, Puhakka J, Rintala J (2009) Impact of crop species on bacterial community structure during anaerobic co-digestion of crops and cow manure. Bioresour Technol 100:2311–2315. doi:10.1016/j.biortech.2008.10.040
Wang H, Tolvanen K, Lehtomäki A, Puhakka J, Rintala J (2010) Microbial community structure in anaerobic co-digestion of grass silage and cow manure in a laboratory continuously stirred tank reactor. Biodegradation 21:135–146. doi:10.1007/s10532-009-9288-5
Acknowledgments
We would like to acknowledge our collaboration partners from Deutsches Biomasseforschungszentrum (DBFZ) for support on chemical analyses. We like to thank Ute Lohse for technical support (T-RFLP), Franziska Bühligen, and Sebastian Röther for operating the reactors and Dirk K. Wissenbach for instructions and providing the GC-MS instrument to measure ratios of 13C-labeled CH4 and CO2.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Funding
This work was supported by the portfolio theme Sustainable Bioeconomy of the Helmholtz Association.
Conflicts of interest
The authors declare that they have no conflict of interest.
Ethical approval
This article does not contain any studies with human participants or animals performed by any of the authors.
Electronic supplementary material
ESM 1
(PDF 116 kb)
Rights and permissions
About this article

Cite this article
Popp, D., Harms, H. & Sträuber, H. The alkaloid gramine in the anaerobic digestion process—inhibition and adaptation of the methanogenic community. Appl Microbiol Biotechnol 100, 7311–7322 (2016). https://doi.org/10.1007/s00253-016-7571-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00253-016-7571-z