
Abstract
Ethyl acetate is an environmentally friendly solvent with many industrial applications. The production of ethyl acetate currently proceeds by energy-intensive petrochemical processes which are based on natural gas and crude oil without exception. Microbial synthesis of ethyl acetate could become an interesting alternative. The formation of esters as aroma compounds in food has been repeatedly reviewed, but a survey which deals with microbial synthesis of ethyl acetate as a bulk product is missing. The ability of yeasts for producing larger amounts of this ester is known for a long time. In the past, this potential was mainly of scientific interest, but in the future, it could be applied to large-scale ester production from renewable raw materials. Pichia anomala, Candida utilis, and Kluyveromyces marxianus are yeasts which convert sugar into ethyl acetate with a high yield where the latter is the most promising one. Special attention was paid to the mechanism of ester synthesis including regulatory aspects and to the maximum and expectable yield. Synthesis of much ethyl acetate requires oxygen which is usually supplied by aeration. Ethyl acetate is highly volatile so that aeration results in its phase transfer and stripping. This stripping process cannot be avoided but requires adequate handling during experimentation and offers a chance for a cost-efficient process-integrated recovery of the synthesized ester.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The microbial formation of carboxylic-acid esters has been repeatedly reviewed. Plenty of various esters are formed where ethyl esters and acetate esters are clearly predominating (Park et al. 2009; Saerens et al. 2010; Sumby et al. 2010; Cordente et al. 2012). These reviews were especially focused on the spectrum of formed esters (Liu et al. 2004; Park et al. 2009; Saerens et al. 2010; Sumby et al. 2010), on ester-forming microbes (Liu et al. 2004; Jolly et al. 2006; Park et al. 2009; Sumby et al. 2010), on the mechanism of ester synthesis (Liu et al. 2004; Park et al. 2009; Saerens et al. 2010), on enzymes being involved in this process (Mason and Dufour 2000; Park et al. 2009; Saerens et al. 2010; Sumby et al. 2010; Cordente et al. 2012), on the related genes and regulation of gene expression (Mason and Dufour 2000), and on metabolic engineering in this field (Park et al. 2009; Cordente et al. 2012). Another extensively reviewed issue concerns ester formation in fermented dairy products (Liu et al. 2004; Longo and Sanromán 2006; Cordente et al. 2012). Esters give alcoholic beverages and fermented food a typical flavor and fragrance where ethyl acetate usually exhibits the highest concentration (Janssens et al. 1992; Verstrepen et al. 2003a; Willaert and Nedovic 2006; Saerens et al. 2008, 2010; Sumby et al. 2010). Ethyl acetate is not only an aroma compound, as one could deduce from the number of publications in this field, but it is much more namely an ester of great industrial relevance. Park et al. (2009) already mentioned the potential of ester-synthesizing microbes for production of esters as industrial solvents.
Ethyl acetate, also known as ethyl ethanoate, is one of the most important carboxylate esters with an estimated annual world production of 1.7 million tons (Posada et al. 2013). The price for ethyl acetate highly fluctuates due to varying feedstock costs and depends on the geographical region; from March 2009 to March 2014, the prices were in the following ranges: 770–1,440 US $/t in the USA, 1,030–1,560 US $/t in West Europe, and 620–1,100 US $/t in China (www.orbichem.com). Ethyl acetate is an organic solvent for countless substances due to its moderate polarity which offers versatile industrial applications. This ester is used as a solvent in chemical reactions, for cleaning surfaces, for processing coating formulations, for extraction and chromatographic recovery of pharmaceuticals, and for production of adhesives, print colors and other paints, herbicide formulations, and resins in the electronics industry. Another interesting application of ethyl acetate could become its use as an acyl acceptor, instead of methanol, at biodiesel production by lipase-catalyzed transesterification of vegetable oil (Kim et al. 2007; Modi et al. 2007; Uthoff et al. 2009; Röttig et al. 2010).
Ethyl acetate is an environmentally friendly solvent since this ester is easily degraded by bacteria; biofilters are preferably used for removing ethyl acetate from exhaust gases of manufacturing plants which are polluted by ethyl acetate and further volatile organics (Hwang et al. 2003; Kam et al. 2005; Koutinas et al. 2005; Chan and Su 2008). Ethyl acetate discharged to the atmosphere is attacked by OH radicals (Atkinson 2000) like its hydrolysis products ethanol and acetic acid (Atkinson 2007). Such oxidative attack of carboxylate esters results in water-soluble organic acids and acid anhydrides (Orlando and Tyndall 2010) which are quickly removed from the atmosphere by rainfall. Ethyl acetate is thus regarded as a non-persistent atmospheric pollutant. Although acting as an irritant and intoxicant in higher concentrations, ethyl acetate is relatively non-toxic to humans compared to many other organic solvents and exhibits an agreeable odor. The global demand of ethyl acetate is thus expected to grow for replacement of other, more harmful solvents.
No review is so far available which is focused on microbial synthesis of ethyl acetate as a bulk product (in the following also referred to as “bulky synthesis”). The presented mini-review wants to close this gap and summarizes the knowledge in this field with special attention on formation of this ester by yeasts.
Current production of ethyl acetate
Synthesis of ethyl acetate currently proceeds by petrochemical processes. The following methods play a larger role in industrial-scale production (Lin et al. 1998; Colley et al. 2004; Arpe 2007; Fig. 1):

Schemes of chemical synthesis of ethyl acetate in industrially relevant production processes
-
a)
Fischer esterification: reaction of ethanol with acetic acid in the presence of strong acids as a catalyst, in part combined with removal of water for shifting the equilibrium toward the ester
-
b)
Tischtschenko reaction: addition of two equivalents acetaldehyde by disproportionation under the action of alkoxides, especially aluminum triethoxide, as a catalyst
-
c)
Dehydrogenation of ethanol: ethanol is dehydrogenated to acetaldehyde, the aldehyde is then added to ethanol to form a hemiacetal which in turn is dehydrogenated to generate ethyl acetate (Inui et al. 2002)
-
d)
Avada process developed by BP Chemicals: Avada abbreviates “Advanced acetates by direct addition” and means synthesis of ethyl acetate by catalytic addition of ethylene and acetic acid in the gas phase
-
e)
Partial oxidation of ethanol plus esterification: this process combines catalytic oxidation of ethanol and esterification of the formed acetic acid with excess ethanol (Lin et al. 1998)
The economy of a given process depends on regional specifics such as availability of raw materials, expenses for all resources, and on fiscal conditions too. All the chemical reactions occur at elevated temperature and often at high pressure. Additional sub-processes are required for recovery of ethyl acetate and for recycling of residual precursors. The conversion process is frequently incomplete due to reverse reactions, and catalysts are commonly required.
Current production of ethyl acetate is exclusively based on fossil hydrocarbons. This becomes clearly visible for the ethylene-based process as ethylene is produced by steam cracking of natural-gas and crude-oil constituents, but this also applies to all other processes since their precursors originate from ethylene; ethanol is formed by its hydration, acetaldehyde is obtained by its oxidation, and acetic acid arises by oxidation of acetaldehyde or ethylene (Fig. 2). Alternative acetic acid production by addition of carbon monoxide and methanol does not change the situation since both are synthesized from natural gas (steam-methane reforming). All these processes require catalysts and a high input of energy (heat and pressure).

Current production of ethyl acetate from natural gas and crude-oil constituents by chemical reactions (reactions labeled by lowercase letters are detailed in Fig. 1) or, alternatively, from sugar as a renewable recourse by microbial processes (labeled by dotted lines)
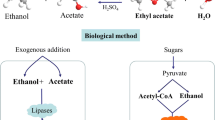
Fossil hydrocarbons are limited resources with steadily growing expenses for their recovery. Ethanol as a precursor for chemical synthesis of ethyl acetate could be produced from sugar by fermentation (Silveira et al. 2005; Aziz et al. 2009; Guimarães et al. 2010; Rodrussamee et al. 2011), but why making a long way around from sugar to ethanol and from ethanol to ethyl acetate when sugar can be converted to ethyl acetate on a straight way by using yeasts (Fig. 2).
A historical survey of bulky formation of ethyl acetate by yeasts
Microbial synthesis of ethyl acetate by yeasts as an aroma compound, typically formed at fermentation of sugars in only small amounts (Jolly et al. 2006; Willaert and Nedovic 2006; Sumby et al. 2010; Cordente et al. 2012), is known for a long time.
In 1892, Beijerinck described a yeast which was characterized by extensive formation of a volatile product and named this isolate Saccharomyces acetaethylicus (Beijerinck 1892); the formed ethyl acetate was, however, only identified by its smell. This species proved to be a synonym of Saccharomyces anomalus which was described in 1891 by Hansen, then reassigned to the new genus Hansenula in 1919 by Sydow, and finally, moved to Pichia in 1984 by Kurtzman (Barnett 2004). Hansen also mentioned a strong fruity smell in cultures of Pichia anomala in his original description of this species. Takahashi and Satō (1911) described four ester-forming P. anomala strains which were isolated from sake. Gray (1949) mentioned similar observations by Seifert, Klöcker, Steuber, Guilliermond and Bedford for P. anomala and Williopsis saturnus in the period from 1897 to 1942. All these authors reported on ethyl acetate as the formed ester but nobody provided any evidence of this speculation.
Gray (1949) was the first who identified and quantified ethyl acetate, ethanol, acetaldehyde and acetate as the products of P. anomala in standing surface cultures through chemical analysis; he also quantified the formed ethyl acetate depending on some parameters like carbon source, initial pH, supplement of yeast extract and supply of oxygen.
Peel and co-workers confirmed synthesis of ethyl acetate by P. anomala by analysis of the ester with hydroxylamine (Peel 1951). Ethanol, acetaldehyde, and acetate were identified as co-metabolites of the glucose metabolism (Davies et al. 1951). It was shown that microbial synthesis of ethyl acetate as a bulk product requires oxygen and that ester synthesis fails at anaerobic conditions (Peel 1951; Davies et al. 1951). The use of rubber plugs for sealing the culture bottles (Peel 1951) refers to their awareness of the volatilization of formed ethyl acetate. Peel (1950, 1951) was the first who studied the mechanism of ester synthesis with resting P. anomala cells.
Tabachnick (1951) received a doctor’s degree for investigating the production of ethyl acetate by P. anomala and published the results in 1953 (Tabachnick and Joslyn 1953a, b). Tabachnick and Joslyn (1953a) identified formed ethyl acetate via physical parameters such as boiling point, refractive index, and density and by chemical analysis (ca. 9 g/L ester accumulated in the culture). Tabachnick and Joslyn (1953a, b) were the first who used shaken flasks instead of standing cultures and tested ethanol as a substrate. They also plausibly demonstrated degradation of accumulated ethyl acetate by P. anomala.
In 1981, Yong et al. observed significant synthesis of ethyl acetate in aged cultures of the soy yeast Saccharomyces rouxii and confirmed the requirement of oxygen for bulky ester formation. The ethyl acetate was obviously produced from ethanol which was previously formed from glucose at semi-anaerobic conditions.
Thomas and Dawson (1978) first described distinct formation of ethyl acetate by Candida utilis during aerobic phased cultivation in a stirred bioreactor and identified acetyl-CoA and ethanol as the two precursors for cell-free synthesis of ethyl acetate. These authors also recognized limitation of growth by iron as a prerequisite for bulky ester formation in this yeast, and they hypothesized that iron limitation restricts oxidation of acetyl-CoA in the citrate cycle and diverts it to ester formation.
Armstrong and co-workers (Armstrong et al. 1984a, b, 1988; Armstrong and Yamazaki 1984; Williams et al. 1988) quantitatively studied ester production by C. utilis with glucose or ethanol as a substrate. The use of Wheaton bottles avoided ester losses by evaporation and generated reliable data (Armstrong et al. 1984a). Depending on the ethanol concentration, ethyl acetate or acetaldehyde was the prevailing product (Armstrong et al. 1984b, 1988).
Willetts (1989) found bulky synthesis of ethyl acetate in Kluyveromyces marxianus. Aerobic conditions and iron limitation as a requirement for bulky ester formation also proved to be true for this yeast, but the yield of ethyl acetate, YEA/S (Table 1), was low.
Kallel-Mhiri and co-workers (Kallel-Mhiri et al. 1993; Kallel-Mhiri and Miclo 1993) first used whey-borne lactose as a renewable raw material to produce ethyl acetate by K. marxianus. However, the ester yield was low when lactose was the sole source of carbon.
Corzo et al. (1995) and Christen et al. (1999) studied formation of ethyl acetate by C. utilis based on ethanol, but the observed yields were quite small.
Then again, the working group of Passoth (Fredlund et al. 2004a, b; Druvefors et al. 2005; Passoth et al. 2006) studied synthesis of ethyl acetate by P. anomala with the intention to suppress mold growth by the formed ester for controlling spoilage of grain. The anti-mold activity of ethyl acetate was demonstrated but the presented data did not allow quantification of ester synthesis.
Since 2008, Bley, Löser, Urit and others (Löser et al. 2011, 2012, 2013; Urit et al. 2010, 2011, 2012, 2013a, b; Urit 2012) extensively studied formation of ethyl acetate from whey-borne lactose by K. marxianus DSM 5422 in many respects. Using one strain for diverse investigations enabled a deeper insight in the physiology and mechanism of ester synthesis. Quasi-continuous and precise analysis of formed ethyl acetate allowed model-based calculation of reliable yields and rates of ester synthesis. Process-relevant information was obtained by variation of technological parameters such as temperature, aeration, availability of iron and other trace elements. Precise data acquisition, the use of a renewable resource and application-oriented research should allow development of a large-scale process in the near future.
Microorganisms producing ethyl acetate
Bacteria and molds are in principle able to produce ethyl acetate but the detected amounts were quite low. Most of the published studies concentrated on microbial formation of this ester as a fragrance and aroma compound in fermented food; synthesis of ethyl acetate was, e.g., described for Lactobacillus strains in sourdough (Hansen and Schieberle 2005), for Acetobacter pusteurianus in vinegar (Kashima et al. 1998), and for molds of the genera Pinicillium and Geotrichum in cheese (Liu et al. 2004).
The most efficient producers of ethyl acetate are yeasts. Westall (1998) tested 169 yeast strains for their ability to produce volatile metabolites during aerobic emerged cultivation; two thirds of the 70 identified volatiles were esters, and ethyl acetate was often the predominating compound formed by yeasts belonging to Candida, Hanseniaspora, Kluyveromyces, Pichia, and other genera.
Most attention in this field has been paid to yeast-mediated formation of ethyl acetate in fermented beverages like beer (Verstrepen et al. 2003a; Willaert and Nedovic 2006; Saerens et al. 2008) and wine (Jolly et al. 2006; Sumby et al. 2010; Cordente et al. 2012). The ester was synthesized by Saccharomyces cerevisiae and so-called non-Saccharomyces wine yeasts of the genera Hanseniaspora, Hansenula, Kloeckera, Kluyveromyces, and Pichia (Rojas et al. 2001, 2003; Plata et al. 2003; Ciani et al. 2006). The amount of ethyl acetate in fermented beverages is typically <50 mg/L which represents a low yield relative to the sugar consumed. Some authors reported on higher concentrations of ethyl acetate in fermented media: 300 mg/L in artificial grape juice fermented with Kloeckera apiculata (Plata et al. 2003), 400 mg/L in grape must with Hanseniaspora guilliermondii (Rojas et al. 2003), 900 mg/L in YNB-glucose medium with P. anomala (Kurita 2008), 1,200 mg/L in grape must with P. anomala (Rojas et al. 2003), and even 1,600 mg/L in sucrose-supplemented grape must with Hanseniaspora uvarum (Ciani et al. 2006). These findings suggest bulky formation of ethyl acetate at anaerobic conditions but, in reality, oxygen had not been excluded completely from the cultivation systems. And the media contained much sugar so that the resulting ester yields were low in spite of quite high ester concentrations.
Table 2 gives an overview on bulky formation of ethyl acetate by yeasts. Only aerobic processes are included in this table since significant synthesis of this ester verifiably requires oxygen (details below). From the economic point of view, two aspects are of main concern in large-scale ester production: the yield of ester and the rate of the process. Formation rates of ethyl acetate have seldom been published so that evaluation is restricted to the yield. Only such publications are included which delivered the desired yield or allowed calculation of this parameter from published data. Ester formation was often only mentioned but not really quantified although title and abstract referred to significant synthesis (Peel 1950, 1951; Laurema and Erkama 1968; Murray et al. 1988; Fredlund et al. 2004b; Druvefors et al. 2005). The yields were used to decide if an ester synthesis is about bulky formation or not. Ester formation was regarded as bulky at YEA/S ≥ 0.01 g/g. The synthesis of ethyl acetate was often studied at varied culture conditions resulting in various ester yields; then, the maximum YEA/S value in the considered publication is given, or several yields are cited together with the specified conditions.
Several yeast species had been identified as potent producers of ethyl acetate (1st column in Table 2) where P. anomala was the first strain which was studied in this relation. This yeast was long time arranged in the genus Hansenula, but recently reclassified in the newly proposed genus Wickerhamomyces (Kurtzman 2011). The genetics, biotechnological potential, and physiology of P. anomala were reviewed by Passoth et al. (2006). The former Hansenula saturnus had been assigned to Williopsis (Kurtzman 1991). S. rouxii is important for flavor formation in soy-sauce fermentation (Yong et al. 1981). Saccharomyces kluyveri produced significant amounts of ethyl acetate (Møller et al. 2002), while its sister, S. cerevisiae, forms only traces of this ester (Kurita 2008). C. utilis is also an interesting candidate for bulky synthesis of ethyl acetate which was studied in the 1980ties and 90ties. Synthesis of ethyl acetate by Kluyveromyces marxianus was first described by Willetts (1989) since Willetts’ Candida pseudotropicalis is the anamorph of K. marxianus (Lachance 1998). In 1993, Kallel-Mhiri and co-workers published ester formation by Kluyveromyces fragilis which is a synonym of K. marxianus (Lachance 1998; Fonseca et al. 2008). Two strains of Kluyveromyces lactis also exhibited distinct formation of ethyl acetate (Löser et al. 2011).
Several substrates were used for producing ethyl acetate such as glucose, whey-borne lactose, ethanol on its own, or combined with sugars (2nd column). Mixed substrates make the evaluation of the process more difficult. Ethanol also appears as an intermediate of sugar consumption and had been postulated as the actual substrate for ester synthesis in P. anomala (Tabachnick and Joslyn 1953a), S. rouxii (Yong et al. 1981), C. utilis (Armstrong et al. 1984a; Armstrong and Yamazaki 1984; Williams et al. 1988), and K. marxianus (Willetts 1989; Kallel-Mhiri et al. 1993). On the contrary, K. marxianus DSM 5422 definitely produced ethyl acetate from lactose but hardly from ethanol (Löser et al. 2011; Urit et al. 2011, 2012).
Most experiments were done under conditions which allowed the yeasts to grow, while a few tests were conducted with resting cells (3rd column). These resting-cell experiments can be considered as whole-cell biotransformation of the substrate (usually ethanol) into ethyl acetate.
The cultivation systems and their modes of operation are also listed in Table 2 (4th column). The reaction vessel and its operation highly influence the course of the process and the rate and yield of microbial ester synthesis. The most crucial points were the handling of ester volatility (see below) and supply of oxygen. Table 2 only summarizes aerobic processes with bulky formation of ethyl acetate; but allowing the oxygen to enter the culture does not automatically mean that transferred oxygen covered the requirement of the yeasts. Attention was, therefore, paid to this subject (5th column). Oxygen limitation was often an unwanted effect resulting from absent shaking (standing cultures), insufficient shaking (temporal O2 limitation at high initial substrate concentrations), or low aeration and/or inadequate stirring. The following phenomena refer to oxygen limitation: a measured pO2 of zero, fermentation of sugars by Crabtree-negative yeasts, and a slow process (see footnotes of Table 2). On the other hand, fully aerobic conditions were confirmed by high pO2 values or absent ethanol formation. Some experiments were aimed at studying ester synthesis at a varied supply of oxygen (Gray 1949; Armstrong et al. 1984a; Bol et al. 1987; Kallel-Mhiri et al. 1993; Corzo et al. 1995). However, postulated stimulation of ester synthesis by oxygen limitation requires critical reviewing (see below).
Effectiveness of ester synthesis was in the first instance evaluated by the yield (6th column). YEA/S is defined as the mass of ethyl acetate formed divided by the mass of substrate consumed. But, strictly speaking, the given YEA/S values are based on masses of detected rather than formed ester. Losses of ester during an experiment result in under-determined yields. This fact explains why experiments with reliable ester analysis often gave higher yields (e.g., bioreactor experiments with K. marxianus DSM 5422). The yield with ethanol as a substrate was, on an average, higher than the yield with sugars, but this seeming advantage is only an illusion as being demonstrated below.
Absolute yields are not really suited for objective comparison since the highest possible yield depends on the substrate. Relative yields (7th column), being absolute yields divided by the theoretically maximum yield for the respective substrate, allow a more realistic view on this subject. The required maximum yields, YEA/S,max, are derived by stoichiometric considerations to a yeast metabolism solely aimed at conversion of the considered substrate into ethyl acetate (details below). A relative yield of 100 % is never obtained with growing yeasts since some substrate is used for biomass formation.
A total number of 23 Kluyveromyces strains were tested for their ability to form ethyl acetate from whey-borne lactose (Löser et al. 2011); 15 of them produced ethyl acetate (YEA/S from 0.07 to 0.19 g/g), while seven of the eight non-productive strains did not utilize lactose, and most probably, do not belong to this genus. Bulky synthesis of this ester by Kluyveromyces strains is thus not an exception but the rule.
Less attention has so far paid to the synthesis rate of ethyl acetate. Laurema and Erkama (1968) were the first who reported on this parameter for P. anomala, but the detected rates were low maybe because of using resting cells. Møller et al. (2002) depicted time-dependent rates for S. kluyveri; ethyl acetate was produced with a constant biomass-specific rate of 0.12 g/g/h and a maximum volume-specific rate of 0.64 g/L/h. Bol et al. (1987) studied synthesis of ethyl acetate by P. anomala at varied conditions and found a maximum productivity of 0.6 g/L/h. Kallel-Mhiri et al. (1993) reported on a maximum productivity of 0.7 g/L/h for K. maxianus in ethanol-supplemented whey medium. Several K. marxianus strains formed ethyl acetate from whey with a rate of 0.4 to 0.8 g/L/h (Löser et al. 2011). A volume-specific formation rate of ca. 3 g/L/h and a biomass-specific rate of 0.4 g/g/h were repeatedly observed during bioreactor cultivation of K. marxianus DSM 5422 in whey-based media (Löser et al. 2012, 2013; Urit et al. 2011, 2012, 2013a). An elevated temperature of 42 °C increased both rates to 4.23 g/L/h and 0.67 g/g/h (Urit et al. 2013b). The highest productivity ever observed amounted to 5.33 g/L/h in highly concentrated whey (Urit et al. 2011).
Comparison of the three most promising yeasts
The most promising candidates for production of ethyl acetate in an industrial scale are P. anomala, C. utilis, and K. marxianus. These yeasts are Crabtree-negative which enables effective cultivation at aerobic conditions.
K. marxianus utilizes a wide spectrum of sugars (Rodrussamee et al. 2011; Fonseca et al. 2013) and is the only of the three yeasts which metabolizes lactose so that whey can be used as a cheap nutrient (Silveira et al. 2005; Guimarães et al. 2010; Urit et al. 2011; Prazeres et al. 2012). Another outstanding characteristic of K. marxianus is its distinct thermal tolerance which allows cultivation at an elevated temperature (Aziz et al. 2009; Rodrussamee et al. 2011; Raimondi et al. 2013). Cultivation at a high temperature advances ester stripping which counteracts accumulation of ethyl acetate to toxic levels and promotes process-integrated product recovery (Urit et al. 2013b). And a high temperature enables non-sterile processes especially with lactose as a specific substrate at a low pH.
Ethyl acetate slows down growth and acts toxic at higher concentrations. The inhibition of growth was found between 12.3 and 17.6 g/L ethyl acetate for P. anomala (Tabachnick and Joslyn 1953b) and at 17 g/L for K. marxianus (Urit et al. 2013a), but no data are available in this respect for C. utilis.
Synthesis of ethyl acetate by C. utilis (Thomas and Dawson 1978; Armstrong and Yamazaki 1984; Armstrong et al. 1984a) and K. marxianus (Willetts 1989; Kallel-Mhiri et al. 1993; Löser et al. 2012; Urit et al. 2012) can be controlled by the level of iron in the culture medium, while for P. anomala, such a control mechanism has not yet been described. Oxygen limitation was believed to stimulate ester formation in P. anomala (Passoth et al. 2006), but this view could be a result of misinterpreted data.
The rate of ester production is also of practical interest. Batch processes with P. anomala (Gray 1949; Davies et al. 1951; Tabachnick and Joslyn 1953a) and C. utilis (Armstrong et al. 1984a; Armstrong and Yamazaki 1984; Williams et al. 1988) were slow and lasted several days, while processes with K. marxianus (Willetts 1989; Kallel-Mhiri et al. 1993; Urit et al. 2011, 2012, 2013a, b; Löser et al. 2013) required only hours. Oxygen limitation could have been a reason for low rates, but the published data do not allow definite evaluation.
Ethanol was repeatedly postulated as an essential intermediate for synthesis of ethyl acetate. P. anomala (Tabachnick and Joslyn 1953a), C. utilis (Armstrong et al. 1984a, b; Corzo et al. 1995; Christen et al. 1999), and K. marxianus (Willetts 1989) can convert ethanol into ethyl acetate, but most processes with significant ester synthesis were based on sugars (Table 2). The sugar was often partially fermented to ethanol where ethyl acetate was formed simultaneously with ethanol (described for P. anomala (Tabachnick and Joslyn 1953a) and C. utilis (Armstrong et al. 1984a; Armstrong and Yamazaki 1984; Williams et al. 1988)), or the ester was synthesized in a later stage from accumulated ethanol (described for P. anomala (Tabachnick and Joslyn 1953a), C. utilis (Armstrong and Yamazaki 1984), and K. marxianus (Willetts 1989; Kallel-Mhiri et al. 1993)). The emerging mode, simultaneous or successive, seems to depend on the supply of oxygen. Oxygen-limited conditions induce fermentation of sugar to ethanol and thus presumably favor ester synthesis from ethanol while fully aerobic conditions promote direct conversion of sugar into ethyl acetate. But some strain-specific characteristics may exist.
Formation of by-products means losses of substrate and pollution of the target product. P. anomala formed ethanol, acetate, and acetaldehyde from glucose (Gray 1949). C. utilis also exhibits a tendency to form these by-products (Armstrong et al. 1984a, b, 1988; Armstrong and Yamazaki 1984; Murray et al. 1988; Christen et al. 1999). Acetaldehyde even became the main product of C. utilis when ethanol was applied in high concentrations (Armstrong et al. 1984b, 1988). K. marxianus formed some ethanol and a little acetate together with ethyl acetate (Willetts 1989; Kallel-Mhiri et al. 1993; Urit et al. 2011, 2012, 2013a, b; Löser et al. 2013). Ethanol formation from sugars by Crabtree-negative yeasts refers to oxygen-limited growth (in most tests with P. anomala and C. utilis) or to deregulation of the yeast metabolism (observed for K. marxianus at adequate aeration (Urit et al. 2011, 2012, 2013a, b; Löser et al. 2013)).
Microbial degradation of formed ethyl acetate would be harmful for the process. If an observed decline in ester concentration was reasoned by degradation or volatilization is often difficult to decide, losses of ester by evaporation or stripping were oftentimes not cared or were tried to avoid by improper measures (details below). The sake-brewing yeast Hansenula mrakii could verifiably utilize ethyl acetate as the sole source for carbon (Inoue et al. 1994). Tabachnick and Joslyn (1953a, b) proved microbial conversion of ethyl acetate into acetate by P. anomala. Disappearance of ethyl acetate and accumulation of acetate were also found with C. utilis (Armstrong and Yamazaki 1984). K. marxianus DSM 5422, on the contrary, never consumed formed ethyl acetate (Löser et al. 2012).
Taken all facts together, K. marxianus seems to be the best candidate for bulky production of ethyl acetate due to its proven ability for direct conversion of sugar into the ester with a high rate and without forming much by-product (aerobic conditions provided), its outstanding thermal tolerance, ability for lactose utilization, and high resistance to ethyl acetate. The ester synthesis is easy to control by the level of iron, and the produced ethyl acetate is obviously not consumed again by this yeast. Moreover, K. marxianus is a dairy yeast with GRAS status. The shadowy existence of K. marxianus, compared to its sister species, K. lactis, is surprising since the properties of this yeast should facilitate its application in biotechnology (Fonseca et al. 2008; Lane and Morrissey 2010).
Mechanisms for synthesis of ethyl acetate
This consideration is restricted to yeasts and concentrates on the crucial reaction step, while the pathways of substrate catabolism for feeding required precursors are not included. Most research in this field were done with S. cerevisiae, although this yeast forms only traces of ethyl acetate (Mason and Dufour 2000; Saerens et al. 2010; Cordente et al. 2012). The findings for S. cerevisiae do not inevitably apply to bulky formation of ethyl acetate by P. anomala, C. utilis, and K. marxianus. Three mechanisms are mainly discussed for yeasts (Liu et al. 2004; Park et al. 2009): esterification, alcoholysis, and the hemiacetal reaction (Fig. 3) which will be considered in general next.

Schemes of microbial synthesis of ethyl acetate in yeasts
Yeasts are believed to synthesize esters by the reversed reaction of esterases known as esterification (Fig. 3a). But the ester-synthesizing esterase activity seems often to be much lower than the ester-hydrolyzing activity (Inoue et al. 1997; Kurita 2008) so that hydrolysis, but not synthesis of esters, is the predominating reaction (Cordente et al. 2012). H. mrakii not only produces ethyl acetate, but also consumes this ester as a substrate; esterases are seemingly involved in both processes (Inoue et al. 1994). Interesterification is a related mechanism of ester formation being catalyzed by lipases or esterases and means that ester neogenesis is based on conversion of another ester in three possible modes: alcoholysis (the ester reacts with an alcohol), acidolysis (the ester reacts with a carboxylic acid), and transesterification (reaction between two esters). The importance of this mechanism for synthesis of ethyl acetate in yeasts is not yet clear (Liu et al. 2004; Sumby et al. 2010).
The formation of ethyl acetate by alcoholysis means reaction of acetyl-CoA with ethanol (Fig. 3b). The involvement of acetyl-CoA in synthesis of ethyl acetate was first described for S. cerevisiae by Nordström (1962). Howard and Anderson (1976) found cell-free synthesis of ethyl acetate from acetyl-CoA and ethanol in S. cerevisiae cell extracts. Alcoholysis is catalyzed by alcohol acetyltransferase (AATase) which was first isolated from S. cerevisiae (Yoshioka and Hashimoto 1981). This mechanism was also found in other yeasts (Thomas and Dawson 1978; Kallel-Mhiri and Miclo 1993; Kusano et al. 1999; Plata et al. 2003). The synthesis of esters by AATase in yeasts was reviewed by Mason and Dufour (2000) and Park et al. (2009).
Distinct ester formation by the hemiacetal reaction was first described by Kusano et al. (1999). Hemiacetals are formed by abiotic condensation of an aldehyde with an alcohol. The hemiacetal is then enzymatically oxidized to an ester where the hydrogen is transferred to NAD(P)+ (Park et al. 2009). The enzymatic reaction is catalyzed by alcohol dehydrogenases, or more specifically, by hemiacetal dehydrogenases (Kusano et al. 1998, 1999; Park et al. 2007). The synthesis of ethyl acetate requires acetaldehyde and ethanol as the precursors (Fig. 3c). The hemiacetal dehydrogenase of Candida utilis exhibited a high potential for synthesis of ethyl acetate (Kusano et al. 1999) while alcohol dehydrogenase of S. cerevisiae (Kusano et al. 1998) and the ADH1 alcohol dehydrogenase of the filamentous fungus Neurospora crassa (Park et al. 2007) synthesized this ester from the respective hemiacetal only with a low rate.
There is another mechanism of ester synthesis from ketones by Baeyer-Villiger monooxygenases which obviously plays only a role in bacteria (Park et al. 2009). Onaca et al. (2007) demonstrated synthesis of ethyl acetate from methylethylketone by Pseudomonas veronii.
Various methods were used to explore the mechanism of ester synthesis. Substrate-conversion studies with intact cells are simple to perform but can produce defective results since the cell metabolism is a network; formation of ethyl acetate from ethanol and acetate, for example, does not inevitably prove esterase activity since the assimilated ethanol could also have been transformed to other metabolites like acetyl-CoA and acetaldehyde which enable ester formation via the other two pathways. Another, more reliable method comprises analysis of enzyme activities in cell extracts with specific substrates but, here, the inducibility of enzymes has to bear in mind at previous yeast cultivation.
The earliest study on the mechanism of ester synthesis in P. anomala was performed with resting cells (Peel 1951); ethyl acetate was formed from ethanol alone or in combination with acetate at aerobic conditions while no ester was produced at anaerobic conditions which rules out ester synthesis via the reversed esterase reaction. Tabachnick and Joslyn (1953b) intensified ester accumulation in resting-cell cultures of P. anomala by esterase inhibitors; this result and the requirement of oxygen let the authors speculate about synthesis of ethyl acetate via an energy-coupled reaction rather than an esterase reaction. Yoshioka and Hashimoto (1981) again observed formation of ethyl acetate by resting P. anomala cells from ethanol plus acetate or from ethanol plus acetyl-CoA and concluded catalysis of ester synthesis by both esterase and AATase, but intact cells produce doubtful results. Bol et al. (1987) presented a hypothetical pathway for synthesis of ethyl acetate in P. anomala from acetyl-CoA and ethanol based on literature where one of the two cited references actually concerns ester synthesis in brewers yeast while the other is missing in the reference list. Kurita (2008) lastly tested enzyme activities in P. anomala cell extracts; the AATase activity was high, the ester-synthesizing esterase activity was low, while the ester-hydrolyzing esterase activity was very high. H. mrakii as a relative of P. anomala gave similar results (Inoue et al. 1997). P. anomala obviously forms ethyl acetate via AATase while the esterases result in net hydrolysis but not synthesis of this ester.
C. utilis was at first believed to produce ethyl acetate in an AATase-catalyzed alcoholysis reaction; in cell extracts of C. utilis, the ester was formed from acetyl-CoA and ethanol but not from acetate and ethanol (Thomas and Dawson 1978). Armstrong and Yamazaki (1984) also assumed exclusive ester formation in this way for C. utilis. Murray et al. (1988) and Corzo et al. (1995) depicted this pathway in detail. Kusano et al. (1999) lastly demonstrated some AATase activity but primarily hemiacetal dehydrogenase activity for several C. utilis strains (the latter was 15 to 100 times higher), i.e., C. utilis produces ethyl acetate mainly via the hemiacetal reaction.
The knowledge for K. marxianus in this regard is comparably low. Synthesis of ethyl acetate by K. marxianus was studied with cell-free extracts (Kallel-Mhiri and Miclo 1993); a constitutive esterase and an inducible AATase were identified, but these findings are doubtful since the activities were, for incomprehensible reason, measured with isoamyl alcohol instead of ethanol (i.e., it was a test for synthesis of isoamyl acetate rather than ethyl acetate). Plata et al. (2003) tested wine yeasts for AATase and ester-synthesizing esterase activities in cell extracts; K. marxianus exhibited both activities where the esterase activity predominated in the early stage and then gradually disappeared, but the results are questionable since the yeasts were cultivated semi-anaerobically. K. marxianus DSM 2254 is believed to synthesize ethyl acetate via AATase since much ester was formed from lactose as a precursor for acetyl-CoA while nearly no ester was produced from ethanol (Löser et al. 2011; Urit et al. 2012).
Hypothetical reasons for synthesis of ethyl acetate
The published ideas regarding the cellular function of synthesis of ethyl acetate and other esters are more or less speculative. Some theories were developed for S. cerevisiae (Peddie 1990; Jain 2010; Saerens et al. 2010) although this yeast produces only traces of esters. Here, only such hypotheses are considered which could by relevant for bulky formation of ethyl acetate at aerobic conditions.
Ester formation for detoxification: This hypothesis has been repeatedly discussed for acetate (Tabachnick and Joslyn 1953a; Fredlund et al. 2004a; Jain 2010), aldehydes (Park et al. 2009; Jain 2010), and medium-chain fatty acids (Peddie 1990; Verstrepen et al. 2003b; Jain 2010). This mechanism could apply to acetaldehyde being eliminated by the hemiacetal reaction (Kusano et al. 1999).
Generation of free coenzyme A: This hypothesis assumes AATase-catalyzed acetate ester formation from acetyl-CoA and alcohols for maintaining a balance between acetyl-CoA and free CoA in the yeast cell (Thomas and Dawson 1978; Verstrepen et al. 2003b; Fredlund et al. 2004a; Saerens et al. 2010). This mechanism could be effective when normal acetyl-CoA regeneration is impaired by a limited acetyl-CoA flux into the citrate cycle, e.g., due to iron limitation.
Anti-fungal activity of esters: suppression of mold growth with P. anomala was explained by its potential for ethyl acetate formation (Fredlund et al. 2004a, b; Druvefors et al. 2005; Passoth et al. 2006; Schnürer and Jonsson 2011). The anti-fungal activity of this ester was demonstrated for the grain-spoilage mold Penicillium roqueforti (Fredlund et al. 2004b; Druvefors et al. 2005).
Ester formation for yeast dissipation: This uncommon hypothesis postulates dissipation of yeasts in nature by insects like Drosophila flies which are attracted by the odor of fermented fruits especially by microbially formed esters (Saerens et al. 2010).
Considerations to the maximum yield of ethyl acetate
The yield of ethyl acetate, YEA/S, is an important parameter for evaluating microbial ester formation. Such yields are useful to compare several processes being performed with the same substrate, but they do not allow realistic comparison when done with various substrates as, e. g., with lactose or ethanol. A more objective base for such examination are relative yields namely the absolute yields related to the respective theoretical maximum yields, YEA/S/YEA/S,max. This ratio represents an index reaching from zero to one, or from 0 to 100 %.
Maximum yields can be derived from the stoichiometry of a yeast metabolism which is assumedly aimed at most effective conversion of the provided substrate into ethyl acetate. The substrate takes thus influence on the YEA/S,max value. Glucose, lactose, and ethanol were typical substrates for bulky formation of ethyl acetate (Table 2). Sugar catabolism results in pyruvate (two moles from glucose, four moles from lactose) and, independently of the special pathway of ester synthesis, two moles pyruvate are required to form one mol ethyl acetate. And two moles ethanol (fed directly or produced from sugar by fermentation) are needed for one mol ethyl acetate. Then, overall balance equations are formulated for the three substrates as previously done for lactose (Urit et al. 2011):
The mass ratios of formed ester and consumed substrate represent the desired yields: YEA/Glucose,max = 0.489 g/g, YEA/Lactose,max = 0.515 g/g, YEA/EtOH,max = 0.958 g/g.
Sugar-ethanol mixtures (Yong et al. 1981; Kallel-Mhiri et al. 1993; Kallel-Mhiri and Miclo 1993) require average maximum yields: YEA/(S1+S2),max = xS1·YEA/S1,max + xS2·YEA/S2,max with xS1 and xS2 being the mass fractions of the two consumed substrates.
The thus-obtained maximum yields were used to evaluate published or from released data derived yields (Table 2). The highest relative ester yield was obtained with ethanol (Bol et al. 1987) but this observation should not be overrated. The maximum yield for ethanol is almost twice as high as the maximum yield for sugars, but this fact is only an advantage at the first view. A two-stage process, where sugar is fermented to ethanol and the ethanol thereafter converted to ester (Fig. 2), exhibits an overall yield of YEtOH/Sugar·YEA/EtOH and a maximum overall yield of YEtOH/Sugar,max·YEA/EtOH,max. For lactose, one gets a maximum yield of 0.538 g/g·0.958 g/g = 0.515 g/g for the total process which is identical with the maximum yield for direct conversion of lactose into ethyl acetate. A high ester yield for ethanol is thus only a pretended benefit. Production of ethyl acetate by a two-stage process (sugar → ethanol, ethanol → ethyl acetate) is actually disadvantageous since it generates higher losses and is typically more cost-intensive than a single-stage process (sugar → ethyl acetate).
Microbial production of ethyl acetate from sugar is actually a superimposition of several sugar-consuming sub-processes: generation of energy by respiration and fermentation, yeast growth, and ester synthesis. These sub-processes can be combined to overall balance equations (Castrillo and Ugalde 1992; Hensing et al. 1995; Mazutti et al. 2010) or, like here, formulated individually. With glucose as a substrate and ammonium as a nitrogen source one gets the following equations:
Published biomass formulae fluctuate since yeast composition varies with conditions (Fonseca et al. 2007): CH1.78O0.75N0.16 (Castrillo and Ugalde 1992), CH1.54O0.54N0.15 (Ghaly and El-Taweel 1997) and CH1.94O0.76N0.17 (Silva-Santisteban et al. 2006) for K. marxianus and CH1.83O0.56N0.17 for K. lactis (Hensing et al. 1995). The source of nitrogen also takes some influence on the balance for yeast growth (Hensing et al. 1995).
Distribution of the available sugar over these sub-processes depends on process conditions: yeast growth was limited by nitrogen, sulfur, or iron (Thomas and Dawson 1977); fermentation was avoided by supply of enough oxygen (Kiers et al. 1998; Parrondo et al. 2009); or ester synthesis was initiated by iron limitation (Thomas and Dawson 1978; Willetts 1989; Löser et al. 2012; Urit et al. 2012).
Anaerobic fermentation of sugars often results in ethanol yields near to the theoretical maximum as observed at fermentation of whey-borne lactose with K. marxianus (Ghaly and El-Taweel 1997; Silveira et al. 2005; Sansonetti et al. 2009; Christensen et al. 2011). Aerobic production of ethyl acetate is less efficient since the oxygen enables other sub-processes such as respiratory energy metabolism and enhanced yeast growth (Urit et al. 2012, 2013b; Löser et al. 2012). These concomitant sub-processes imply losses of sugar in reference to the ester yield resulting in YEA/S values being distinctly smaller than YEA/S,max. This explains why YEA/S–YEA/S,max ratios were rarely larger than 50 % (Table 2).
The highest relative ester yield with whey amounted to 56 % (Urit et al. 2013b). Whey is a waste of milk processing and represents a renewable resource. Whey-borne lactose is therefore an interesting substrate in biotechnology and subject of numerous reviews (Mawson 1994; González Siso 1996; Pesta et al. 2007; Smithers 2008; Guimarães et al. 2010; Prazeres et al. 2012). Conversion of whey-borne sugar into ethyl acetate has never been mentioned there, although such a process was already described in 1993 (Kallel-Mhiri et al. 1993; Kallel-Mhiri and Miclo 1993).
Regulation of ester synthesis
Oxygen is definitely required for bulky formation of ethyl acetate as becoming visible from the overall balance equations of ester synthesis. Much ethyl acetate was formed by P. anomala (Davies et al. 1951; Tabachnick and Joslyn 1953a, b), C. utilis (Willetts 1989), S. rouxii (Yong et al. 1981), and K. marxianus (Kallel-Mhiri and Miclo 1993) at aerobic conditions, while ester production was negligible at anaerobic conditions. Aeration stimulated ester formation by P. anomala (Gray 1949; Laurema and Erkama 1968; Bol et al. 1987; Rojas et al. 2001), C. utilis (Armstrong et al. 1984a; Armstrong and Yamazaki 1984; Corzo et al. 1995), and K. marxianus (Kallel-Mhiri et al. 1993), but intensive aeration seemingly diminished ester synthesis again (Gray 1949; Armstrong et al. 1984a; Bol et al. 1987; Kallel-Mhiri et al. 1993). Corzo et al. (1995) presumed stripping as the actual reason for decreasing ester concentrations at high aeration. Or the yeast metabolism was maybe inhibited by using pure oxygen (Kallel-Mhiri et al. 1993; Kallel-Mhiri and Miclo 1993) since a high pO2 value causes cell stress and a reduced metabolic activity in K. marxianus (Pinheiro et al. 2002, 2003; Urit et al. 2013a).
Availability of oxygen is, in fact, an obligatory but not sufficient precondition for significant formation of ethyl acetate. One well-known trigger of ester formation is iron; ethyl acetate was formed by C. utilis (Thomas and Dawson 1978; Armstrong and Yamazaki 1984; Armstrong et al. 1984a) and K. marxianus (Willetts 1989; Kallel-Mhiri et al. 1993; Löser et al. 2012; Urit et al. 2012) at iron limitation while supplementing the medium with iron repressed this ester formation. Iron-dependent ester synthesis by P. anomala has not yet been studied. The formation of ethyl acetate by S. kluyveri was not suppressed by iron (Møller et al. 2002); ester formation in this yeast may occur due to metabolic overflow since no ester appeared at glucose-limited cultivation.
Armstrong and Yamazaki (1984) were the first to report on synthesis of ethyl acetate by C. utilis at various iron concentrations in the medium, but the experimental conditions were not well-defined, the iron varied only a little, and neither yeast growth nor iron uptake was measured. Löser et al. (2012, 2013) studied iron-dependent formation of ethyl acetate by K. marxianus DSM 5422 extensively; at low doses, the iron was entirely absorbed and the amount of formed biomass was governed by a minimum iron content in the yeasts (8.8 μg/g), while at higher doses, sugar was the growth-limiting factor, iron was more or less taken up and the biomass formation was directed by the available sugar. Intensive ester synthesis required a low content of iron in the yeast biomass (Löser et al. 2012, 2013).
Thomas and Dawson (1978) explained ester formation at iron-limited growth of C. utilis with a reduced flux of acetyl-CoA into the citrate cycle due to a diminished activity of aconitase and succinate dehydrogenase (both enzymes depend on iron) followed by diversion of acetyl-CoA for synthesis of ethyl acetate. On the other hand, C. utilis seems to produce ethyl acetate mainly via the hemiacetal pathway rather than from acetyl-CoA in an AATase catalyzed reaction (Kusano et al. 1999). Thomas and Dawson (1978) also speculated on involvement of the electron-transferring proteins since iron limitation reduced the available energy. Armstrong and Yamazaki (1984) observed formation of ethyl acetate by C. utilis even at higher levels of iron when EDTA was added to the medium; they speculated about an increased membrane permeability for ethanol as a precursor for ethyl acetate but, more likely, formation of EDTA-iron chelates prevents iron absorption by the yeasts (Kosman 2003) and iron-limited growth finally results in ester formation.
Synthesis of ethyl acetate by K. marxianus DSM 5422 was also triggered by copper limitation (Urit et al. 2010, 2012) which supports the postulated involvement of the respiratory chain. The electron-transferring proteins depend on iron (Levi and Rovida 2009) and complex IV also requires copper (Cuillel 2009). A deficit in one of these two metals makes the electron transport and NADH oxidation slow. Diminished oxidation of mitochondrial NADH produces a lack of mitochondrial NAD+ which is required for several citrate-cycle reactions. Lacking NAD+ finally reduces the flux of acetyl-CoA into the citrate cycle and diverts this compound to ester synthesis. This view is in accordance with metabolite-profiling studies during cultivation of K. marxianus at various levels of iron limitation (Löser et al. 2012). Involvement of the respiratory chain has also been demonstrated by initiating ester synthesis in K. marxianus by carboxin, antimycin A, or cyanide, which are specific inhibitors for the electron-transferring proteins (Löser et al., unpublished results). Oxygen-limited cultivation of K. marxianus DSM 5422 also induced some ester synthesis since oxygen is the terminal electron acceptor (Löser et al., unpublished results).
Iron is considered as the best factor for controlling ester synthesis in K. marxianus compared to other principles; oxygen limitation favors ethanol synthesis, only traces of copper are required so that copper limitation is difficult to adjust (Urit et al. 2012), and respiratory inhibitors are highly toxic substances. The intracellular storage of iron complicates matters: the rate of ester synthesis actually depends on the intracellular iron concentration which, in turn, is controlled by both the applied iron and the produced biomass (Löser et al. 2012, 2013).
Volatility of ethyl acetate and consequences
The microbial synthesis of ethyl acetate usually occurs in aqueous culture media, but the ester is highly volatile and easily migrates to the gas phase. This phase transfer has to take into account during lab-scale experiments in shaken bottles but also during processes in aerated bioreactors. The volatility of the ester is very important in three respects: (a) improper experimentation results in losses of the ester, (b) precise quantification of ester synthesis must take the evaporation of ester into account, and (c) volatility of ethyl acetate enables process-integrated product recovery.
Most of such laboratory experiments were performed in culture bottles. The sealing of such bottles was often not described (Gray 1949; Armstrong et al. 1984a, b; Willetts 1989; Christen et al. 1999; Rojas et al. 2001, 2003). The use of cotton stoppers (Tabachnick and Joslyn 1953b) allows the formed ester to disappear via diffusion (Fig. 4). On the other hand, Tabachnick and Joslyn (1953a) were the first who debated the problem of evaporation and tested the disappearance of the ester and ethanol from shaken flasks; they stated that evaporation was not relevant, but their data are doubtful since ethanol was identified to be more volatile than ethyl acetate, but the opposite is true. The use of rubber stoppers (Peel 1951; Tabachnick and Joslyn 1953b; Laurema and Erkama 1968) bear the risk that ester is absorbed by the stopper material where silicone rubber is most problematical. Yong et al. (1981) used cotton plugs and tried to absorb formed ethyl acetate from the headspace gas by hydroxylamine but they did not test the efficiency of this method. Christen et al. (1999) again used ethanol as a substrate and tested its evaporation from shake flasks but they did not care evaporation of formed ethyl acetate.

Superimposition of microbial synthesis of ethyl acetate in the liquid phase, phase transfer to the gas phase and discharge from the cultivation system results in uncontrollable losses of ester (cotton-plugged shake flasks) or in ester stripping (aerated bioreactors)
There are also some examples of reliably sealed culture vessels. Davies et al. (1951) were the first who used a sealed system with absorption of formed carbon dioxide and recharge of consumed oxygen. The use of Wheaton bottles is another positive example (Armstrong et al. 1984a; Armstrong and Yamazaki 1984), but the sampling during the experiment is hindered. Septum-sealed bottles allows repeated sampling of the headspace and/or the liquid medium by syringes pierced through the septum without the risk of ester losses (Löser et al. 2011, 2012). The operational reliability of this system was proven by repeated analysis of shaken aqueous ester solutions (Urit et al. 2013a).
In such sealed flasks, formed ethyl acetate partially migrates from the liquid to the headspace gas. The mass of formed ester is then the sum of dissolved and evaporated ethyl acetate both being calculated from the volume and concentration in the respective phase (Löser et al. 2011; the symbols of this and the following equations are explained in Table 1):
This method requires time-consuming ester analyses in both phases. Intensive shaking, however, causes a quick phase transfer of ethyl acetate (quick relative to the synthesis rate) and results in quasi-equilibrium conditions and thus produces a constant relation between the gas and liquid-phase ester concentration which is described by the partition coefficient (Urit et al. 2011):
The partition coefficient for ethyl acetate in a water-air system was estimated from the saturation concentrations in both phases (Urit et al. 2011); the KEA,L/G value highly depends on the temperature since the water solubility decreases while the vapor pressure (determining the maximum gas-phase content) decreases with growing temperature. KEA,L/G values were also determined experimentally at low ester concentrations in equilibrated systems (Löser et al. 2011; Urit et al. 2011) and by stripping tests (Urit et al. 2011); the obtained KEA,L/G values were somewhat smaller than the calculated data. Liquid-media constituents like sugar and mineral salts distinctly reduce both the ester solubility and partition coefficient (Covarrubias-Cervantes et al. 2004, 2005; Löser et al. 2011; Urit et al. 2011).
In quasi-equilibrated systems, the liquid-phase concentration can be substituted by the KEA,L/G value. The mass of formed ester is so simply calculable from measured headspace concentrations (Löser et al. 2011):
Synthesis of ethyl acetate was also studied in aerated bioreactors where aeration inevitably results in a discharge of synthesized ester (stripping; Fig. 4). Bol et al. (1987) were the first who studied microbial ester synthesis in such a system but did not mention the problem of stripping at all. Kallel-Mhiri et al. (1993) and Kallel-Mhiri and Miclo (1993) placed a condenser at the gas outlet for minimizing stripping of volatiles, but calculations and experiments (Urit et al. 2011) clearly demonstrated the inefficiency of this measure. Even a refrigerant of −10 °C (Corzo et al. 1995) condenses gaseous ester only in part. But a freezing trap kept at −80 °C (Murray et al. 1988) should capture nearly all stripped ethyl acetate. Fredlund et al. (2004a) tried to trap stripped ester in decane but their data refer to very incomplete absorption. Incomplete capture of ethyl acetate from the exhaust gas was also observed with solid adsorbents such as activated carbon and resins (Medeiros et al. 2006).
Stripping of ethyl acetate from aerated cultivation systems can hardly be prevented but requires adequate handling. One way to create reliable data is the regular quantification of ethyl acetate in the exhaust gas and culture medium (Löser et al. 2012, 2013; Urit et al. 2011, 2012, 2013a, b). Exclusive quantification of ester in the exhaust gas (Thomas and Dawson 1978; Møller et al. 2002) allows calculation of the mass of stripped ester but is insufficient for calculating ester formation rates since this approach ignores temporal accumulation of ethyl acetate in the liquid phase.
Studying the ester stripping from aerated bioreactors was aimed at identifying key factors which influence this process. A model was derived for several reasons: for understanding the stripping, for quantifying the ester formation, and for model-based process design (Urit et al. 2011, 2013a). The presented equations are valid for the batch mode but are modifiable for continuous processes (Löser et al. 2012). This modeling starts with two balance equations describing the changes of the mass of ester in both phases (Urit et al. 2011). After transforming the masses into concentrations one gets:
The liquid concentration changes due to microbial formation with the rate REA and due to phase transfer to the headspace by evaporation (the second term) while CEA,G changes by this phase transfer too and by discharge of the ester with the exhaust gas (Fig. 4). The temporal change of CEA,G is usually small and, with dCEA,G/dt ≈ 0, Eq. (5) is transformed into the following equation:
Combining Eqs. (4) and (6) gives:
In aerated stirred reactors, the kEA,La value (the phase-transfer coefficient) is typically much higher than the (FG/VL)-KEA,L/G ratio so that Eq. (7) can be simplified again:
Abiotic stripping tests (REA = 0) in an aerated stirred reactor clearly demonstrated the validity of this simplified equation (Urit et al. 2011); the stripping rate was proportional to the gas-flow rate while a varied phase-transfer coefficient did not influence the stripping. The same was observed during microbial ester synthesis in aerated bioreactors (Urit et al. 2013a). The stripping of ethyl acetate was governed by the absorption capacity of the exhaust gas but not by the phase transfer since a high phase-transfer coefficient produces near-equilibrium conditions (CEA,L ≈ KEA,L/G·CEA,G). Some deviation from this equilibrium was only observed in slowly shaken culture bottles (Löser et al. 2011). Installation of an exhaust-gas condenser neither retained any ester nor changed the course of stripping (Urit et al. 2011).
The temperature also influences the ester stripping since the partition coefficient highly depends on this parameter. Urit et al. (2013b) studied the formation of ethyl acetate by K. marxianus in a stirred reactor at 32 and 42 °C; the temperature distinctly affected the CEA,L-CEA,G ratio as expected; the high temperature facilitated ester stripping and counteracted accumulation of ester to inhibitory amounts. This is relevant since ethyl acetate distinctly inhibits yeast growth (Tabachnick and Joslyn 1953b; Urit et al. 2013a).
Produced ethyl acetate migrates from the liquid to the headspace and is then discharged with the exhaust gas. The mass of ester which is stripped till a given time is calculated as follows (Urit et al. 2011):
In the case of discrete CEA,G measurement, the integral is replaced by a sum of FG·CEA,G(t)·Δt terms. Precise calculation also regards the temporal variance of the exhaust-gas flow (FG); FG is derived from the supplied gas flow and the exhaust-gas composition (Duboc and von Stockar 1998; Löser et al. 2011).
The mass of formed ethyl acetate represents the ester which has been stripped till a given time plus the ester which has already been formed but not yet stripped (ester still dissolved in the liquid phase):
In systems with a high VG/VL ratio, the ester in the headspace (evaporated but not yet discharged) can become relevant; then a third term must be added to Eq. (10) namely VG·CEA,G(t) (Löser et al. 2011).
The volume-specific rate of ester synthesis is obtained by differentiation of Eq. (10) and subsequent division by the liquid volume (assuming an invariant VL):
This volume-specific formation rate is the mass of ester synthesized per liter reaction volume and per hour and characterizes the efficiency of the process. The biomass-specific synthesis rate of ethyl acetate is, in contrast, a biological variable which describes the efficiency of the yeasts and is given in grams of ester formed per hour and per gram of biomass (rEA = REA/CX).
Volatility characterizes the disappearance of a considered compound from a given system. Ethyl acetate and ethanol possess nearly the same boiling point (77.2 and 78.3 °C at 1 bar) and their vapor pressure–temperature curves are similar (http://ddbonline.ddbst.de) but the ester disappears much quicker. This is surprising at first view but becomes clear when taking into account that the volatility is also determined by the solubility in the liquid. Ethanol is limitless mixable with water while ethyl acetate is less water-soluble. The partition coefficient is therefore better suited for rating the volatility since it depends on both the vapor pressure and solubility (Urit et al. 2011). The behavior of ethyl acetate or ethanol in water–air and whey–air systems was comparatively studied (Löser et al. 2011); KEA,L/G was circa 30 times smaller than KEtOH,L/G or, in other words, ethyl acetate was 30 times more volatile than ethanol. In stripping tests (0.8 L liquid aerated with 30 L/h air), ethanol disappeared with a specific rate of 0.0142 h−1 (Löser et al. 2005), while the ester was stripped with a rate of 0.43 h−1 (Urit et al. 2011) which gives a ratio of 1:30.
Process-integrated product recovery
The stripping of ethyl acetate complicates quantification of ester synthesis, but it also offers a chance for process-integrated product recovery at future large-scale ester production. Such a product recovery generally improves economy by accelerating the process and by omission of process stages (here, by combining fermentation and product extraction).
In situ product recovery was extensively studied at 2-phenylethanol synthesis by biotransformation with yeasts, but the cytotoxic 2-phenylethanol is hardly volatile and therefore preferably extracted during the fermentation process (Mei et al. 2009; Hua and Xu 2011). In situ product recovery by evaporation was proposed for microbially formed acetaldehyde but not tested (Armstrong et al. 1984b). Process-integrated recovery of microbially produced butanol and by-products via stripping had been repeatedly tested (Qureshi and Blaschek 2001; Lu et al. 2012; de Vrije et al. 2013; Setlhaku et al. 2013; Xue et al. 2013). Abdel-Banat et al. (2010) studied the recovery of ethanol, high ethanol concentrations, an elevated temperature, and a reduced pressure improved evaporation, but this was not enough for practical application.
Armstrong et al. (1984a) studied microbial conversion of ethanol into ethyl acetate and proposed solvent extraction of the ester, but the high volatility of ethyl acetate predestines its process-integrated recovery by evaporation and subsequent separation from the gas phase. The stripped ester could be separated by adsorption (Manjare and Ghoshal 2006a, b, c) or perhaps by membrane processes.
References
Abdel-Banat BMA, Hoshida H, Ano A, Nonklang S, Akada R (2010) High-temperature fermentation: how can processes for ethanol production at high temperatures become superior to the traditional process using mesophilic yeast? Appl Microbiol Biotechnol 85:861–867
Armstrong DW, Yamazaki H (1984) Effect of iron and EDTA on ethyl acetate accumulation in Candida utilis. Biotechnol Lett 6:819–824
Armstrong DW, Martin SM, Yamazaki H (1984a) Production of ethyl acetate from dilute ethanol solutions by Candida utilis. Biotechnol Bioeng 26:1038–1041
Armstrong DW, Martin SM, Yamazaki H (1984b) Production of acetaldehyde from ethanol by Candida utilis. Biotechnol Lett 6:183–188
Armstrong DW, Martin SM, Yamazaki H (1988) Selective production of ethyl acetate and acetaldehyde by microorganisms. U.S. Pat 4,720,457
Arpe H-J (2007) Industrielle Organische Chemie, Bedeutende Vor- und Zwischenprodukte. Wiley-VCH, Weinheim
Atkinson R (2000) Atmospheric chemistry of VOCs and NOx. Atmos Environ 34:2063–2101
Atkinson R (2007) Gas-phase tropospheric chemistry of organic compounds: a review. Atmos Environ 41:S200–S240
Aziz S, Memon HUR, Shah FA, Rajoka MI, Soomro SA (2009) Production of ethanol by indigenous wild and mutant strain of thermotolerant Kluyveromyces marxianus under optimized fermentation conditions. Pak J Anal Environ Chem 10(1+2):25–33
Barnett JA (2004) A history of research on yeasts 8: taxonomy. Yeast 21:1141–1193
Beijerinck MW (1892) Zur Ernährung des Kahmpilzes. Zentbl Bakt Parasitenk Abt I 11:68–75
Bol J, Knol W, ten Brik B (1987) Optimization of the production of ethyl acetate from ethanol by Hansenula anomala. Dechema Monogr 105:235–236
Castrillo JI, Ugalde UO (1992) Energy metabolism of Kluyveromyces marxianus in deproteinated whey. Chemostat studies. Modelling. J Biotechnol 22:145–152
Chan WC, Su MQ (2008) Biofiltration of ethyl acetate and amyl acetate using a composite bead biofilter. Bioresour Technol 99:8016–8021
Christen P, Domenech F, Páca J, Revah S (1999) Evaluation of four Candida utilis strains for biomass, acetic acid and ethyl acetate production from ethanol. Bioresour Technol 68:193–195
Christensen AD, Kádár Z, Oleskowicz-Popiel P, Thomsen MH (2011) Production of bioethanol from organic whey using Kluyveromyces marxianus. J Ind Microbiol Biotechnol 38:283–289
Ciani M, Beco L, Comitini F (2006) Fermentation behaviour and metabolic interactions of multistarter wine yeast fermentations. Int J Food Microbiol 108:239–245
Colley SW, Fawcett CR, Rathmell C, Marshall MW (2004) Process for the preparation of ethyl acetate. US Pat 6,809,217
Cordente AG, Curtin CD, Varela C, Pretorius IS (2012) Flavour-active wine yeasts. Appl Microbiol Biotechnol 96:601–618
Corzo G, Revah S, Christen P (1995) Effect of oxygen on the ethyl acetate production from continuous ethanol stream by Candida utilis in submerged cultures. Dev Food Sci 37B:1141–1154
Covarrubias-Cervantes M, Champion D, Debeaufort F, Voilley A (2004) Aroma volatility from aqueous sucrose solutions at low and subzero temperatures. J Agric Food Chem 52:7064–7069
Covarrubias-Cervantes M, Bongard S, Champion D, Voilley A (2005) Temperature effect on solubility of aroma compounds in various aqueous solutions. Lebensm-Wiss Technol 38:371–378
Cuillel M (2009) The dual personality of ionic copper in biology. J Incl Phenom Macrocycl Chem 65:165–170
Davies R, Falkiner EA, Wilkinson JF, Peel JL (1951) Ester formation by yeasts 1. Ethyl acetate formation by Hansenula species. Biochem J 49:58–61
de Vrije T, Budde M, van der Wal H, Claassen PAM, López-Contreras AM (2013) “In situ” removal of isopropanol, butanol and ethanol from fermentation broth by gas stripping. Bioresour Technol 137:153–159
Druvefors UÄ, Passoth V, Schnürer J (2005) Nutrient effects on biocontrol of Penicillium roqueforti by Pichia anomala J121 during airtight storage of wheat. Appl Environ Microbiol 71:1865–1869
Duboc P, von Stockar U (1998) Systematic errors in data evaluation due to ethanol stripping and water vaporization. Biotechnol Bioeng 58:428–439
Fonseca GG, Gombert AK, Heinzle E, Wittmann C (2007) Physiology of the yeast Kluyveromyces marxianus during batch and chemostat cultures with glucose as the sole carbon source. FEMS Yeast Res 7:422–435
Fonseca GG, Heinzle E, Wittmann C, Gombert AK (2008) The yeast Kluyveromyces marxianus and its biotechnological potential. Appl Microbiol Biotechnol 79:339–354
Fonseca GG, de Carvalho NMB, Gombert AK (2013) Growth of the yeast Kluyveromyces marxianus CBS 6556 on different sugar combinations as sole carbon and energy source. Appl Microbiol Biotechnol 97:5055–5067
Fredlund E, Blank LM, Schnürer J, Sauer U, Passoth V (2004a) Oxygen- and glucose-dependent regulation of central carbon metabolism in Pichia anomala. Appl Environ Microbiol 70:5905–5911
Fredlund E, Druvefors UÄ, Olstorpe MN, Passoth V, Schnürer J (2004b) Influence of ethyl acetate production and ploidy on the anti-mould activity of Pichia anomala. FEMS Microbiol Lett 238:133–137
Ghaly AE, El-Taweel AA (1997) Continuous ethanol production from cheese whey fermentation by Candida pseudotropicalis. Energy Source 19:1043–1063
González Siso MI (1996) The biotechnological utilization of cheese whey: a review. Bioresour Technol 57:1–11
Gray WD (1949) Initial studies on the metabolism of Hansenula anomala (Hansen) Sydow. Am J Bot 36:475–480
Guimarães PMR, Teixeira JA, Domingues L (2010) Fermentation of lactose to bio-ethanol by yeasts as part of integrated solutions for the valorisation of cheese whey. Biotechnol Adv 28:375–384
Hansen A, Schieberle P (2005) Generation of aroma compounds during sourdough fermentation: applied and fundamental aspects. Trends Food Sci Technol 16:85–94
Hensing MCM, Bangma KA, Raamsdonk LM, de Hulster E, van Dijken JP, Pronk JT (1995) Effects of cultivation conditions on the production of heterologous α-galactosidase by Kluyveromyces lactis. Appl Microbiol Biotechnol 43:58–64
Howard D, Anderson RG (1976) Cell-free synthesis of ethyl acetate by extracts from Saccharomyces cerevisiae. J Inst Brew 82:70–71
Hua D, Xu P (2011) Recent advances in biotechnological production of 2-phenylethanol. Biotechnol Adv 29:654–660
Hwang S-CJ, Lee C-M, Lee H-C, Pua HF (2003) Biofiltration of waste gases containing both ethyl acetate and toluene using different combinations of bacterial cultures. J Biotechnol 105:83–94
Inoue Y, Fukuda K, Wakai Y, Sudsai T, Kimura A (1994) Ester formation by a yeast Hansenula mrakii IFO 0895: contribution of esterase for iso-amyl acetate production in sake brewing. Lebensm-Wiss Technol 27:189–193
Inoue Y, Trevanichi S, Fukuda K, Izawa S, Wakai Y, Kimura A (1997) Roles of esterase and alcohol acetyltransferase on production of isoamyl acetate in Hansenula mrakii. J Agric Food Chem 45:644–649
Inui K, Kurabayashi T, Sato S (2002) Direct synthesis of ethyl acetate from ethanol carried out under pressure. J Catal 212:207–215
Jain VK (2010) Modifying redox potential and its impact on metabolic fluxes in Saccharomyces cerevisiae. Dissertation, Stellenbosch University, Cape Town, South Africa
Janssens L, De Pooter HL, Schamp NM, Vandamme EJ (1992) Production of flavours by microorganisms. Process Biochem 27:195–215
Jolly NP, Augustyn OPH, Pretorius IS (2006) The role and use of non-Saccharomyces yeasts in wine production. S Afr J Enol Vitic 27:15–39
Kallel-Mhiri H, Miclo A (1993) Mechanism of ethyl acetate synthesis by Kluyveromyces fragilis. FEMS Microbiol Lett 111:207–212
Kallel-Mhiri H, Engasser J-M, Miclo A (1993) Continuous ethyl acetate production by Kluyveromyces fragilis on whey permeate. Appl Microbiol Biotechnol 40:201–205
Kam S-K, Kang K-H, Lee M-G (2005) Removal characteristics of ethyl acetate and 2-butanol by a biofilter packed with jeju scoria. J Microbiol Biotechnol 15:977–983
Kashima Y, Iijima M, Okamoto A, Koizumi Y, Udaka S, Yanagida F (1998) Purification and characterization of intracellular esterases related to ethylacetate formation in Acetobacter pasteurianus. J Ferment Bioeng 85:584–588
Kiers J, Zeeman A-M, Luttik M, Thiele C, Castrillo JI, Steensma HY, van Dijken JP, Pronk JT (1998) Regulation of alcoholic fermentation in batch and chemostat cultures of Kluyveromyces lactis CBS 2359. Yeast 14:459–469
Kim S-J, Jung S-M, Park Y-C, Park K (2007) Lipase catalyzed transesterification of soybean oil using ethyl acetate, an alternative acyl acceptor. Biotechnol Bioprocess Eng 12:441–445
Kosman DJ (2003) Molecular mechanisms of iron uptake in fungi. Mol Microbiol 47:1185–1197
Koutinas M, Peeva LG, Livingston AG (2005) An attempt to compare the performance of bioscrubbers and biotrickling filters for degradation of ethyl acetate in gas streams. J Chem Technol Biotechnol 80:1252–1260
Kurita O (2008) Increase of acetate ester-hydrolysing esterase activity in mixed cultures of Saccharomyces cerevisiae and Pichia anomala. J Appl Microbiol 104:1051–1058
Kurtzman CP (1991) DNA relatedness among saturn-spored yeasts assigned to the genera Williopsis and Pichia. Antonie Van Leeuwenhoek 60:13–19
Kurtzman CP (2011) Phylogeny of the ascomycetous yeasts and the renaming of Pichia anomala to Wickerhamomyces anomalus. Antonie Van Leeuwenhoek 99:13–23
Kusano M, Sakai Y, Kato N, Yoshimoto H, Sone H, Tamai Y (1998) Hemiacetal dehydrogenation activity of alcohol dehydrogenases in Saccharomyces cerevisiae. Biosci Biotechnol Biochem 62:1956–1961
Kusano M, Sakai Y, Kato N, Yoshimoto H, Tamai Y (1999) A novel hemiacetal dehydrogenase activity involved in ethyl acetate synthesis in Candida utilis. J Biosci Bioeng 87:690–692
Lachance MA (1998) Kluyveromyces van der Walt emend. van der Walt. In: Kurtzman CP, Fell JW (eds) The yeasts, a taxonomic study, 4th edn. Elsevier, Amsterdam, pp 227–247
Lane MM, Morrissey JP (2010) Kluyveromyces marxianus: a yeast emerging from its sister’s shadow. Fungal Biol Rev 24:17–26
Laurema S, Erkama J (1968) Formation of ethyl acetate in Hansenula anomala. Acta Chem Scand 22:1482–1486
Levi S, Rovida E (2009) The role of iron in mitochondrial function. Biochim Biophys Acta 1790:629–636
Lin T-B, Chuang KT, Tsai K-Y, Chang J-R (1998) Process for ethyl acetate production. US Pat 5,770,761
Liu S-Q, Holland R, Crow VL (2004) Esters and their biosynthesis in fermented dairy products: a review. Int Dairy J 14:923–945
Longo MA, Sanromán MA (2006) Production of food aroma compounds: microbial and enzymatic methodologies. Food Technol Biotechnol 44:335–353
Löser C, Schröder A, Deponte S, Bley T (2005) Balancing the ethanol formation in continuous bioreactors with ethanol stripping. Eng Life Sci 5:325–332
Löser C, Urit T, Nehl F, Bley T (2011) Screening of Kluyveromyces strains for the production of ethyl acetate: design and evaluation of a cultivation system. Eng Life Sci 11:369–381
Löser C, Urit T, Förster S, Stukert A, Bley T (2012) Formation of ethyl acetate by Kluyveromyces marxianus on whey during aerobic batch and chemostat cultivation at iron limitation. Appl Microbiol Biotechnol 96:685–696
Löser C, Urit T, Stukert A, Bley T (2013) Formation of ethyl acetate from whey by Kluyveromyces marxianus on a pilot scale. J Biotechnol 163:17–23
Lu C, Zhao J, Yang S-T, Wei D (2012) Fed-batch fermentation for n-butanol production from cassava bagasse hydrolysate in a fibrous bed bioreactor with continuous gas stripping. Bioresour Technol 104:380–387
Manjare SD, Ghoshal AK (2006a) Comparison of adsorption of ethyl acetate on activated carbon and molecular sieves 5A and 13X. J Chem Eng Data 51:1185–1189
Manjare SD, Ghoshal AK (2006b) Adsorption equilibrium studies for ethyl acetate vapor and E-Merck 13X molecular sieve system. Sep Purif Technol 51:118–125
Manjare SD, Ghoshal AK (2006c) Studies on adsorption of ethyl acetate vapor on activated carbon. Ind Eng Chem Res 45:6563–6569
Mason AB, Dufour J-P (2000) Alcohol acetyltransferases and the significance of ester synthesis in yeast. Yeast 16:1287–1298
Mawson AJ (1994) Bioconversions for whey utilization and waste abatement. Bioresour Technol 47:195–203
Mazutti MA, Zabot G, Boni G, Skovronski A, de Oliveira D, Di Luccio M, Rodrigues MI, Maugeri F, Treichel H (2010) Mathematical modeling of Kluyveromyces marxianus growth in solid-state fermentation using a packed-bed bioreactor. J Ind Microbiol Biotechnol 37:391–400
Medeiros ABP, Pandey A, Vandenberghe LPS, Pastore GM, Soccol CR (2006) Production and recovery of aroma compounds produced by solid-state fermentation using different adsorbents. Food Technol Biotechnol 44:47–51
Mei J, Min H, Lü Z (2009) Enhanced biotransformation of L-phenylalanine to 2-phenylethanol using an in situ product adsorption technique. Process Biochem 44:886–890
Modi MK, Reddy JRC, Rao BVSK, Prasad RBN (2007) Lipase-mediated conversion of vegetable oils into biodiesel using ethyl acetate as acyl acceptor. Bioresour Technol 98:1260–1264
Møller K, Christensen B, Förster J, Piškur J, Nielsen J, Olsson L (2002) Aerobic glucose metabolism of Saccharomyces kluyveri: growth, metabolite production, and quantification of metabolic fluxes. Biotechnol Bioeng 77:186–193
Murray WD, Duff SJB, Lanthier PH, Armstrong DW, Welsh FW, Williams RE (1988) Development of biotechnological processes for the production of natural flavors and fragrances. Dev Food Sci 17:1–18
Nordström K (1962) Formation of ethyl acetate in fermentation with brewer’s yeast. III. Participation of coenzyme A. J Inst Brew 68:398–407
Onaca C, Kieninger M, Engesser K-H, Altenbuchner J (2007) Degradation of alkyl methyl ketones by Pseudomonas veronii MEK700. J Bacteriol 189:3759–3767
Orlando JJ, Tyndall GS (2010) The atmospheric oxidation of ethyl formate and ethyl acetate over a range of temperatures and oxygen partial pressures. Int J Chem Kinet 42:397–413
Park Y-C, San K-Y, Bennett GN (2007) Characterization of alcohol dehydrogenase 1 and 3 from Neurospora crassa FGSC2489. Appl Microbiol Biotechnol 76:349–356
Park YC, Shaffer CEH, Bennett GN (2009) Microbial formation of esters. Appl Microbiol Biotechnol 85:13–25
Parrondo J, García LA, Díaz M (2009) Nutrient balance and metabolic analysis in a Kluyveromyces marxianus fermentation with lactose-added whey. Braz J Chem Eng 26:445–456
Passoth V, Fredlund E, Druvefors UÄ, Schnürer J (2006) Biotechnology, physiology and genetics of the yeast Pichia anomala. FEMS Yeast Res 6:3–13
Peddie HAB (1990) Ester formation in brewery fermentations. J Inst Brew 96:327–331
Peel JL (1950) Formation of ethyl acetate by yeast. J Gen Microbiol 4:iv–v, Communication
Peel JL (1951) Ester formation by yeasts. 2. Formation of ethyl acetate by washed suspensions of Hansenula anomala. Biochem J 49:62–67
Pesta G, Meyer-Pittroff R, Russ W (2007) Utilization of whey. In: Oreopoulou V, Russ W (eds) Utilization of by-products and treatment of waste in the food industry. Springer, New York, pp 193–207
Pinheiro R, Belo I, Mota M (2002) Oxidative stress response of Kluyveromyces marxianus to hydrogen peroxide, paraquat and pressure. Appl Microbiol Biotechnol 58:842–847
Pinheiro R, Belo I, Mota M (2003) Growth and β-galactosidase activity in cultures of Kluyveromyces marxianus under increased air pressure. Lett Appl Microbiol 37:438–442
Plata C, Millán C, Mauricio JC, Ortega JM (2003) Formation of ethyl acetate and isoamyl acetate by various species of wine yeasts. Food Microbiol 20:217–224
Posada JA, Patel AD, Roes A, Blok K, Faaij APC, Patel MK (2013) Potential of bioethanol as a chemical building block for biorefineries: preliminary sustainability assessment of 12 bioethanol-based products. Bioresour Technol 135:490–499
Prazeres AR, Carvalho F, Rivas J (2012) Cheese whey management: a review. J Environ Manag 110:48–68
Qureshi N, Blaschek HP (2001) Recovery of butanol from fermentation broth by gas stripping. Renew Energy 22:557–564
Raimondi S, Zanni E, Amaretti A, Palleschi C, Uccelletti D, Rossi M (2013) Thermal adaptability of Kluyveromyces marxianus in recombinant protein production. Microb Cell Factories 12(34):1–7
Rodrussamee N, Lertwattanasakul N, Hirata K, Suprayogi, Limtong S, Kosaka T, Yamada M (2011) Growth and ethanol fermentation ability on hexose and pentose sugars and glucose effect under various conditions in thermotolerant yeast Kluyveromyces marxianus. Appl Microbiol Biotechnol 90:1573–1586
Rojas V, Gil JV, Piñaga F, Manzanares P (2001) Studies on acetate ester production by non-Saccharomyces wine yeasts. Int J Food Microbiol 70:283–289
Rojas V, Gil JV, Piñaga F, Manzanares P (2003) Acetate ester formation in wine by mixed cultures in laboratory fermentations. Int J Food Microbiol 86:181–188
Röttig A, Wenning L, Bröker D, Steinbüchel A (2010) Fatty acid alkyl esters: perspectives for production of alternative biofuels. Appl Microbiol Biotechnol 85:1713–1733
Saerens SMG, Verstrepen KJ, Thevelein JM, Delvaux FR (2008) Ethyl ester production during brewery fermentation: a review. Cerevisia 33(2):82–90
Saerens SMG, Delvaux FR, Verstrepen KJ, Thevelein JM (2010) Production and biological function of volatile esters in Saccharomyces cerevisiae. Microb Biotechnol 3:165–177
Sansonetti S, Curcio S, Calabrò V, Iorio G (2009) Bio-ethanol production by fermentation of ricotta cheese whey as an effective alternative non-vegetable source. Biomass Bioenergy 33:1687–1692
Schnürer J, Jonsson A (2011) Pichia anomala J121: a 30-year overnight near success biopreservation story. Antonie Van Leeuwenhoek 99:5–12
Setlhaku M, Heitmann S, Górak A, Wichmann R (2013) Investigation of gas stripping and pervaporation for improved feasibility of two-stage butanol production process. Bioresour Technol 136:102–108
Silva-Santisteban BOY, Converti A, Filho FM (2006) Intrinsic activity of inulinase from Kluyveromyces marxianus ATCC 16045 and carbon and nitrogen balances. Food Technol Biotechnol 44:479–483
Silveira WB, Passos FJV, Mantovani HC, Passos FML (2005) Ethanol production from cheese whey permeate by Kluyveromyces marxianus UFV-3: a flux analysis of oxido-reductive metabolism as a function of lactose concentration and oxygen levels. Enzym Microb Technol 36:930–936
Smithers GW (2008) Whey and whey proteins—from ‘gutter-to-gold’. Int Dairy J 18:695–704
Sumby KM, Grbin PR, Jiranek V (2010) Microbial modulation of aromatic esters in wine: current knowledge and future prospects. Food Chem 121:1–16
Tabachnick J (1951) The chemistry and physiology of ester production by Hansenula anomala. Dissertation, University of California, Berkeley
Tabachnick J, Joslyn MA (1953a) Formation of esters by yeast. I. The production of ethyl acetate by standing surface cultures of Hansenula anomala. J Bacteriol 65:1–9
Tabachnick J, Joslyn MA (1953b) Formation of esters by yeast. II. Investigations with cellular suspensions of Hansenula anomala. Plant Physiol 28:681–692
Takahashi T, Satō H (1911) Some new varieties of Willia anomala as aging yeast of saké. J Coll Agric 1:227–269
Thomas KC, Dawson PSS (1977) Variations in the adenylate energy charge during phased growth (cell cycle) of Candida utilis under energy excess and energy-limiting growth conditions. J Bacteriol 132:36–43
Thomas KC, Dawson PSS (1978) Relationship between iron-limited growth and energy limitation during phased cultivation of Candida utilis. Can J Microbiol 24:440–447
Urit T (2012) Ethyl acetate formation by Kluyveromyces marxianus on whey. Dissertation, Dresden University of Technology, Dresden
Urit T, Löser C, Stukert A, Bley T (2010) Bildung von Ethylacetat durch Kluyveromyces marxianus aus Molke bei aerober batch-Kultivierung unter Spurenelemente-Limitation. Chem Ing Technik 82:1550
Urit T, Löser C, Wunderlich M, Bley T (2011) Formation of ethyl acetate by Kluyveromyces marxianus on whey: studies of the ester stripping. Bioprocess Biosyst Eng 34:547–559
Urit T, Löser C, Stukert A, Bley T (2012) Formation of ethyl acetate by Kluyveromyces marxianus on whey during aerobic batch cultivation at specific trace-element limitation. Appl Microbiol Biotechnol 96:1313–1323
Urit T, Manthey R, Bley T, Löser C (2013a) Formation of ethyl acetate by Kluyveromyces marxianus on whey: influence of aeration and inhibition of yeast growth by ethyl acetate. Eng Life Sci 13:247–260
Urit T, Li M, Bley T, Löser C (2013b) Growth of Kluyveromyces marxianus and formation of ethyl acetate depending on temperature. Appl Microbiol Biotechnol 97:10359–10371
Uthoff S, Bröker D, Steinbüchel A (2009) Current state and perspectives of producing biodiesel-like compounds by biotechnology. Microb Biotechnol 2:551–565
Verstrepen KJ, Derdelinckx G, Dufour J-P, Winderickx J, Thevelein JM, Pretorius IS, Delvaux FR (2003a) Flavor-active esters: adding fruitiness to beer. J Biosci Bioeng 96:110–118
Verstrepen KJ, Van Laere SDM, Vanderhaegen BMP, Derdelinckx G, Dufour J-P, Pretorius IS, Winderickx J, Thevelein JM, Delvaux FR (2003b) Expression levels of the yeast alcohol acetyltransferase genes ATF1, Lg-ATF1, and ATF2 control the formation of a broad range of volatile esters. Appl Environ Microbiol 69:5228–5237
Westall S (1998) Characterisation of yeast species by their production of volatile metabolites. J Food Mycol 1:187–201
Willaert R, Nedovic VA (2006) Primary beer fermentation by immobilised yeast—a review on flavour formation and control strategies. J Chem Technol Biotechnol 81:1353–1367
Willetts A (1989) Ester formation from ethanol by Candida pseudotropicalis. Antonie Van Leeuwenhoek 56:175–180
Williams RE, Armstrong DW, Murray WD, Welsh FW (1988) Enzyme and whole cell production of flavor and fragrance compounds. Ann N Y Acad Sci 542:406–412
Xue C, Zhao J, Liu F, Lu C, Yang S-T, Bai F-W (2013) Two-stage in situ gas stripping for enhanced butanol fermentation and energy-saving product recovery. Bioresour Technol 135:396–402
Yong FM, Lee KH, Wong HA (1981) The production of ethyl acetate by soy yeast Saccharomyces rouxii NRRL Y-1096. J Food Technol 16:177–184
Yoshioka K, Hashimoto N (1981) Ester formation by alcohol acetyltransferase from brewers’ yeast. Agric Biol Chem 45:2183–2190
Declaration
The authors declare that they have no conflict of interest.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Löser, C., Urit, T. & Bley, T. Perspectives for the biotechnological production of ethyl acetate by yeasts. Appl Microbiol Biotechnol 98, 5397–5415 (2014). https://doi.org/10.1007/s00253-014-5765-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00253-014-5765-9