
Abstract
Mutation and polymorphism detection by nucleases has become a more important tool in clinical and biological researches. There are several kinds of single-stranded nucleases for detecting mismatched DNAs. One of them, CEL II, was isolated from Apium graveolens and cleaves DNA with high specificity at sites of mismatch. High-throughput mutation scanning requires large quantity of CEL II endonuclease. Here, we demonstrate high-level expression of CEL II using silkworm-baculovirus system. The recombinant CEL II secreted in silkworm hemolymph was glycosylated and susceptible to N-glycosidase F. Additionally, larger metal ions such as Ca2+ and Sr2+ were able to replace Mg2+ and enhanced mismatch cleavage activity of CEL II. These results indicate that the silkworm-baculovirus platform is a good alternative system to obtain the functional CEL II.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Nucleases are required for multiple processes of DNA metabolism, such as DNA replication, DNA repair, and recombination in vivo. They have been also widely used as experimental tools for in vitro and in vivo manipulation of DNA. Above all, single-stranded DNA-specific nucleases have been isolated from a wide variety of organisms. Some enzymes such as S1 nuclease (Aspergillus oryzae), P1 nuclease (Penicillium citrinum), BAL 31 nuclease (Alteromonas espejiana), Mung bean nuclease, and Micrococcal nuclease (Staphylococcus aureus) have been well characterized and are now available commercially (Desai and Shankar 2003). Unfortunately, these activities are too strong to detect small mismatches on a polyacrylamide gel.
There is another group of single-stranded DNA-specific nucleases, which effectively detect mismatches, insertion, and deletion of heteroduplex DNA by cutting of single-stranded regions in double-stranded nucleic acids. This approach is important to identify single nucleotide polymorphisms and disease-causing mutations in the genome. Recently, gene editing by zinc finger nucleases or transcription activator-like effector nucleases (TALENs) seems to be promising. It would be indispensable to construct a high-throughput validation system for isolating mutants without marker molecules or visible phenotypes. Several enzymes including T4 endonuclease VII, endonuclease V, T7 endonuclease I, CEL I, and Surveyor nuclease (CEL II) have been investigated so far. At present, CEL II was reported to show the best efficiency compared to T4E7, T7EI, and Endo V (Tsuji and Niida 2008).
The silkworm/baculovirus expression system (BES) is believed to be an excellent system to high-level production of recombinant protein (Lee et al. 2012b). It has some advantages with no limitation about the size of the protein, posttranslational modifications, disulfide bond formation, and proteolytic processing. In addition, we have reported that silkworm would be suitable for the large-scale production of proteins and several strains of silkworm which was highly effective to foreign protein expression by Bombyx mori nucleopolyhedrovirus (BmNPV) (Kawakami et al. 2008; Lee et al. 2012b). The native CEL I and CEL II have been used for detecting mutations so far, and the purification of the native proteins requires time and labor. Pimkin et al. (2007) demonstrated that the recombinant CEL I was expressed and purified using Autographa california nucleopolyhedrovirus and the insect cell line Sf9 from Spodoptera frugiperda. It has been reported that the CEL II protein has a secretion signal sequence and N-linked glycosylation, implying that BES would be an alternative way to produce the recombinant CEL II.
In this study, we carried out the production of the recombinant CEL II in the silkworm larvae using BES. The purified CEL II was functionally active in mutation detection. Further, we found that calcium and strontium ions enhanced the cleavage activity efficiently better than any of the ions tested. A BmNPV-based silkworm expression system is suitable for industrial expression of eukaryotic recombinant proteins.
Materials and methods
Cells and silkworms
Bme21 and BmN4-Luc5′3′DR cells were cultured in IPL-41 medium (Sigma no. I7760) supplemented with 10 % fetal bovine serum (Mon et al. 2011; Lee et al. 2012a). B. mori strain d17 used in this study was obtained from the Institute of Genetic Resources, Kyushu University Graduate School. The larvae were reared on mulberry leaves at 25 °C (Kawakami et al. 2008).
Construction of the Cel II expression vector
Total RNA was extracted from leaves of commercially grown celery (Apium graveolens) using ISOGEN (Nippon Gene) according to the manufacturer’s instruction. cDNA was synthesized from the total RNA using the Superscript III (Life Technologies). The complete ORF region of CEL II was amplified with the primers AgCel-IIatg (5′-GGTCCATGGGTATGTTGACTTATACTGGAATTTATTTTCTGC-3′) and AgCel-IInostop (5′-CGACTCGAGACTATTTCAATATTGTTACTGCTTCTATGTCC-3′) using KOD-Plus-Neo polymerase (TOYOBO). The modified pENTR11 (pENTR11L21TEVH8) was used for the addition of a poly-histidine tag at the C-terminus of CEL II. The nucleotide sequence of pENTR11L21TEVH8 is available upon request. The amplified PCR product was digested with NcoI and XhoI and ligated with the NcoI/XhoI cleaved pENTR11L21TEVH8 plasmid. The resulting plasmid was recombined with the Gateway destination vector pDEST8 using the LR Clonase II Enzyme mix (Life Technologies). The resultant plasmid was designated pDEST8/CEL-IItevH8.
Construction of the recombinant baculovirus
Baculovirus carrying the CEL II gene was constructed as described elsewhere (Ono et al. 2007; Lee et al. 2012a). Briefly, Escherichia coli BmT3DH10Bac containing bacmid and helper plasmid (pMON7124) was transformed with pDEST8/CEL-IItevH8. Colonies containing the bacmid for expression of CEL II were identified by colony PCR using primers LacM13 (5′-CACACAGGAAACAGCTATGACCATGATTAC-3′) and AgCel-IIstop5 (5′-CCAGCGATCTCACAAGATTGAATATGC3′). The bacmid DNA was purified using the QIAGEN Large-Construct kit. The bacmid was transfected into Bme21 cells using Fugene HD (Promega) to yield BmNPV/T3 carrying the CEL II expression cassette (BmNPV/T3-CEL-II). High-titer virus stock was prepared to infect the silkworm larvae by subcutaneous injection.
Purification of recombinant CEL II
The baculovirus was injected directly into the third day of fifth-instar silkworm larvae. The hemolymph was collected at 84 hpi by cutting the caudal leg in a tube containing 5 μl of 200 mM 1-phenyl-2-thiourea. The hemolymph was centrifuged at 8,500 × g for 10 min at 4 °C. The supernatant was further centrifuged at 90,000 × g for 30 min, and the 90,000 × g supernatant fraction was diluted with 70 ml of buffer A (20 mM Hepes, pH 7.4, 500 mM NaCl, 10 % glycerol) containing a protease inhibitor cocktail (Complete, Roche). The sample was applied to a 5-ml HisTrap HP (GE) column, followed by washing the column with 50 ml of buffer A containing 40 mM imidazole. CEL II was eluted with a linear gradient of imidazole 40–300 mM in buffer A. CEL II-containing fractions were pooled and dialyzed overnight against dialysis buffer (50 mM Tris–HCl, pH 7.5, 100 mM KCl, 0.01 % Triton X-100, 10 μM ZnCl2, 50 % glycerol). The purity of protein was analyzed by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) followed by silver staining (Silver Stain II kit, Wako).
Deglycosylation of the recombinant CEL II
Approximately 1 μg of CEL II was denatured with 1× glycoprotein denaturing buffer at 95 °C for 10 min. After the addition of NP-40 and G7 reaction buffer, CEL II was incubated with PNGase F (New England Biolabs) at 37 °C for 18 h. Likewise, non-denatured CEL II was subjected to deglycosylation with PNGase F. The protein sample without PNGase F was used as a control. The deglycosylated protein was subjected to SDS-PAGE followed by Western blot analysis using HisProbe-HRP (Pierce). The presence of PNGase F was confirmed by Ponceau S staining of the PVDF membrane.
RF-I nicking assays
The nicking activity of CEL II was examined using supercoiled plasmid DNA. Supercoiled plasmid DNA (RF-I; 0.5 μg) was incubated with the designated amounts of CEL II for 30 min at 37 °C in 20 μl of buffer B (20 mM HEPES, pH 7.5, 10 mM KCl, 3 mM MgCl2). To stop the enzyme reaction, 2 μl of stop solution (50 mM EDTA, 1 μg/μl Proteinase K) was added to each reaction. The reaction mixtures were analyzed by agarose gel electrophoresis.
Mismatch cleavage assays
Two plasmids containing either wild-type or 6-bp deletion mutant of human KNL1 gene (GenBank accession number NM170589) were amplified using KOD-Plus-NEO polymerase (TOYOBO). For preparation of heteroduplex DNA substrates, the PCR products without any purification were mixed at a 1:1 ratio and incubated at 95 °C for 5 min, then gradually cooled to room temperature with a cooling rate of 1 °C/min. The other two types of DNA substrates (G/C mismatch, 1-bp deletion) were prepared in the same manner as described above. For the detection of single-nucleotide polymorphisms (SNPs), substrates were amplified by PCR using purified genomic DNA from Metarhizium and cDNA from the silkworm strains k45 and d17. The sequences of primers used to amplify the Metarhizium 28S ribosomal DNA (rDNA) were Metarhizium-rDNAF1 (5′- ATCCAGCGGTTCCCGTTGGTGCACTTTG-3′) and Metarhizium-rDNAR1 (5′- GCCCTGGAGGGCTCTCACCTGCGTTCAC-3′). The sequences of primers used to amplify the silkworm Atonal-like protein 8 gene (BmATOH8, accession number KC118544) were BmATOH8F1 (5′- CGAACCGCAAGGACTTCCGAGAGTGCC-3′) and BmATOH8R1 (5′- CTGTCCGGCTCGATGACTGCGGAAAGTG-3′).
For mismatch endonuclease assay, the substrate DNA (350 ng) was mixed with 5 ng of the purified CEL II in 15 μl of reaction buffer (20 mM Tris–HCl, pH 7.5, 25 mM KCl) with 2–50 mM of the indicated divalent ion. The standard enzyme reaction was performed at 45 °C for 60 min and stopped by adding 2 μl of stop solution (50 mM EDTA, 1 μg/μl Proteinase K) at 37 °C for 30 min. The incubation time and the reaction temperature were changed as indicated in the legend of Fig. 3. The reaction products were fractionated by 8 % native polyacrylamide gel electrophoresis in TBE.
Design and construction of TALEN
To generate the TALENs targeting the modified luciferase gene (Luc5′ΔC) (Mon et al. 2011), TALE repeats were assembled using the Golden Gate cloning (Cermak et al. 2011). The vectors used for the assembly were obtained from Addgene (http://www.addgene.org). The sequences targeted by the TALENs were as follows: DR5 (5′-CCATTTTTTGAAGCGAAGGT-3′) and DR3 (5′-CCTGCAGATTACCCTGTTAT-3′). The spacer length between the TALEN-binding sites was 16 bp.
To perform rapid and convenient construction of many different types of expression vector, we generated the Gateway entry plasmid, pENTR11m-TALEN, which contains the N- and C-terminal flanking regions of TALE and the catalytic domain of FokI. First, the Esp3I site within the kanamycin-resistant gene in pENTR11 (Life Technologies) was deleted to create pENTR11m. Second, the pTAL3-derived fragment (the TALE flanking sequences and the lacZ gene) was cloned into pENTR11m. Finally, a sequence encoding a nuclear localization signal was added into the downstream of the attL1 site. The resulting plasmid, pENTR11m-TALEN, was used for the step 2 ligation in the Golden Gate assembly (Cermak et al. 2011). pENTR11m-TALEN/DR5 and pENTR11m-TALEN/DR3 were recombined with the insect expression plasmid pie2FW (Yamashita et al. 2007; Tatsuke et al. 2009).
Mutation detection by CEL II
The TALEN-DR5 and TALEN-DR3 expression vectors were transfected into BmN4-Luc5′3′DR cells using Fugene HD (Promega), according to the manufacturer’s instructions. The cells were harvested 10 days after transfection and lysed in 50 mM NaOH solution. The extracts were incubated for 10 min at 95 °C, followed by neutralization with Tris–HCl (pH 8.0). Total genomic DNAs were used for PCR with the KOD FX DNA polymerase (TOYOBO). Primer sequences were as follows: primer A (5′-AGGCCCGGCGCCATTCTATCCTCTAG-3′) and primer B (5′-AGTAAAACCTCTACAAATGTGGTAAAATCG-3′). The amplified PCR products were used for the nested PCR. The nested PCR primers were as follows: primer C (5′-ACCCTATTTTCATTCTTCGCCAAAAGCACTCTGATTG-3′) and primer D (5′- ATCCTTTGTATTTAATTAAAGACTTCAAGCG-3′). As a control, the pLuc5′3′DR plasmid DNA was used (Mon et al. 2011).
The second PCR products were heated to 95 °C and gradually cooled to 4 °C to allow formation of heteroduplexes. The resulting products were mixed with 10 ng of the purified CEL II in 20 μl of reaction buffer (20 mM Tris–HCl, pH 7.5, 25 mM KCl, 10 mM SrCl2). The reaction was conducted at 45 °C for 60 min and stopped by adding 2 μl of stop solution at 37 °C for 30 min. The reaction products were fractionated by 8 % native PAGE. As a positive control of homologous recombination, the intact luciferase sequence and the modified luciferase sequence (Luc5′ΔC) were amplified by PCR, followed by CEL II digestion.
Nucleotide sequence submission
Nucleotide sequence data of CEL II have been submitted to the GenBank database with the accession number AB746184.
Results
Single-step purification of the recombinant CEL II endonuclease
Recombinant protein expression system using baculovirus/silkworm is an extremely valuable tool for the large-scale production of proteins with posttranslational modifications such as glycosylation, phosphorylation, disulfide bond formation, and proteolytic processing (Kato et al. 2009; Lee et al. 2012b). The celery mismatch-specific endonuclease, CEL II, has a putative signal peptide for secretion and asparagine-linked (N-linked) glycosylation (Yang et al. 2000). These properties encouraged us to produce the recombinant CEL II in silkworm larvae. For ease of purification, an 8xHis-tag was added to the 3′ end of the CEL II ORF. The B. mori nucleopolyhedrovirus bacmid to express CEL II was produced by transposition in E. coli (Fig. 1a). To manipulate B. mori nucleopolyhedrovirus (BmNPV/T3-CEL-II), we used the Bme21 cell line, which is appropriate for large amounts of high-titer virus (Lee et al. 2012a).

Purification and characterization of the recombinant CEL II nuclease. a Schematic representation of BmNPV/T3. The bacmid DNA contained the CEL II expression cassette under the control of the strong polyhedrin promoter. The CEL II gene was fused with a poly-histidine tag (His8) at the C-terminus. b Purification and SDS-PAGE analysis of the recombinant CEL II. The purified protein was analyzed by 10 % SDS-PAGE followed by silver staining. The asterisk indicates a background band. c SDS-PAGE analysis of the deglycosylated CEL II after incubation with (plus sign) and without (minus sign) PNGase F. Left panel, Ponceau stain of the membrane; right panel, Western blotting with HisProbe. Lanes 1 and 5, PNGase F treated CEL II under denaturing conditions; 2 and 6, PNGase F treated CEL II under non-denaturing conditions; 3 and 7, untreated CEL II; 4 and 8, PNGase F control. d Single-strand-specific nuclease activity detected by RF-I nicking assay. The supercoiled plasmid was incubated with or without CEL II as indicated. The terms RF I and RF II denote supercoiled and nicked plasmid DNA, respectively
To produce recombinant CEL II, BmNPV/T3-CEL-II was injected into silkworm larvae. Because the infected silkworms began to die 96 h after injection, the hemolymph from silkworm larvae was harvested 84 h after injection. The supernatant of the hemolymph was purified by nickel-affinity chromatography. The fraction containing CEL II was dialyzed and stored at −80 °C. About 400 μg of protein was obtained from 20 ml of the hemolymph (Fig. 1b).
Since native CEL II has been reported to possess N-linked glycosylation, deglycosylation analysis of the recombinant CEL II was performed using N-glycosidase F (PNGase F), which cleaves N-linked oligosaccharides from glycoproteins. Treatment with PNGase F reduced the molecular mass (Fig. 1c), indicating that the recombinant CEL II produced in silkworm larvae was indeed glycosylated with N-linked oligosaccharides. We could not find any difference in the degree of deglycosylation under either the native or denaturing condition (Fig. 1c, lanes 5 and 6).
CEL family proteins, CEL I and CEL II, have been reported that they cleave heteroduplex DNA specifically. The nuclease activity of purified recombinant CEL II was monitored by a replicative form I (RF-I) nicking assay. As shown in Fig. 1d, we could detect the nicking activity of recombinant CEL II. Supercoiled RF-I DNA was efficiently converted to nicked circular DNA (RF-II) by even 2.5 ng of the enzyme. These results demonstrated that BES was an alternative method to produce CEL II.
Alkaline earth metal ions, calcium, and strontium activate the mismatch nuclease activity of CEL II
CEL II belongs to S1/P1 nuclease family and has five putative Zn2+-binding sites (Romier et al. 1998). CEL I also require Mg2+ for activity. Because divalent metal ions appear to be required for efficient DNA mismatch nuclease activity, we explored the possibility that other divalent metal ions could replace Zn2+ and Mg2+ and increase its activity. The survey of alternative cofactors indicated that Ca2+ and the heavier alkaline earth metal ion Sr2+ were effective cofactors, stimulating mismatch-specific incisions (Fig. 2a). Cleavage products were not observed when Zn2+, Cu2+, Ni2+, and Co2+ were used. We also found that higher concentration of the divalent metal ions, Ca2+ and Sr2+, inhibited the nuclease activity, and the maximum permissive concentration of Sr2+ was a little higher than that of Ca2+ (Fig. 2b). In the experiments described below, we used the divalent metal ion Sr2+ in the reaction buffer.

Calcium and strontium activate the mismatch nuclease activity of CEL II. All reactions were performed for 60 min at 45 °C. a The reannealed PCR products were incubated with CEL II in reaction buffer with the indicated ions at a concentration of 10 mM. b The reannealed PCR products were incubated with CEL II in reaction buffer with the indicated concentrations of Ca2+ or Sr2+
Optimum incubation time and temperature for the CEL II activity
Reaction conditions for CEL II were optimized for cleavage efficiency. Optimum incubation time was 60 min (Fig. 3a), and background smears were found with a longer time of incubation for 90 min. CEL II was efficient at mismatch cleavage between 45 and 50 °C (Fig. 3b).

The optimum incubation time and temperature for the CEL II nuclease activity. a The reannealed PCR products were incubated with CEL II for the indicated incubation time. All reactions were performed at 45 °C with 10 mM strontium. b The reannealed PCR products were incubated with CEL II at the indicated temperature. All reactions were performed for 60 min with 10 mM strontium
Examples of mutation detection with CEL II
Substrate DNAs containing different types of mismatches were tested for further application of the recombinant CEL II. PCR products with a G to C base substitution or an A extrahelical loop were cleaved by CEL II, respectively (Fig. 4a, b). To further explore the utility of CEL II for more practical applications, we used SNPs in the ribosomal DNA sequences of Metarhizium spp. and in the atonal-like protein 8 (BmATOH8) sequences of the silkworm strains k45 and d17 (Lee et al. 2007; Kawaguchi et al. 2009; Nishi et al. 2011). The rDNA sequence has been widely used to evaluate the genetic diversity in fungi. The region of 28S rDNA from two isolates (Hkd25-1 and Oit8-3) was amplified by PCR, and the SNPs were detected by CEL II (Fig. 4c). In addition, we utilized CEL II to the detection of SNPs in the BmATOH8 gene (Fig. 4d). The amplified fragment which contained three SNPs was cleaved by CEL II. These results indicate that the recombinant CEL II could be applied for mutation detection and SNP genotyping.

CEL II digestion products of heteroduplex DNA containing several mismatches. a, b Single mismatch or A extrahelical loop substrates were digested using CEL II nuclease and analyzed by agarose gel electrophoresis. All reactions were performed at 45 °C with 10 mM strontium. c The detection of two SNPs (two T to C base substitutions) in the 283-bp PCR product. The substrate is the 28S rDNA of Metarhizium with the two substitutions at position 114 and 121. d The detection of three SNPs (two C to T base substitutions and one T to C base substitution) in the 285-bp PCR product. The substrate is the atonal-like protein 8 gene in B. mori
CEL II detected mutations mediated by TALEN
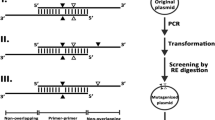
Recently, technologies for targeted genome editing have been developed, and mismatch-specific endonucleases are widely used for the screening of genomic mutations (Yeung et al. 2005; Kim et al. 2009). To investigate the capability of recombinant CEL II for detecting mutations, we used the TALENs targeted to the modified luciferase gene (Cermak et al. 2011; Mon et al. 2011). In the BmN4-Luc5′3′DR cells, the inactive luciferase would recover its activity through double-stranded break (DSB) which induced repair as described in Fig. 5a. One of the TALEN target was located upstream of the I-SceI recognition site, and the other was within the I-SceI recognition sequence (Fig. 5a). DSBs induced by the TALENs could be repaired by homologous recombination (HR) or nonhomologous end joining (NHEJ) (Mon et al. 2011).

CEL II detected mutations mediated by TALEN. a The recombination repair substrate in BmN4Luc5′3′DR. It contains two mutated luciferase genes, Luc5′ΔC and Luc3′ΔA as direct repeats under the control of SK8 promoter. When a pair of TALENs is expressed in cells, DSBs are introduced around the I-SceI site. The two red arrows indicate the primers for the nested PCR. b Schematic diagram of the heteroduplex formation and the enzyme mismatch cleavage. DSBs repair by HR or NHEJ generates three types of heteroduplex DNA molecules in theory. HR causes loss of the I-SceI site and gain of the intact luciferase sequence. NHEJ mainly causes deletion of some base pairs around the I-SceI site. The predicted sizes of the digested DNA fragments are shown. c The nested PCR products were incubated with CEL II, and the extent of cleavage was analyzed by agarose gel electrophoresis
We introduced the pair of the TALEN expression plasmids into BmN4-Luc5′3′DR and extracted genomic DNA (Mon et al. 2011). After PCR amplification of the genomic DNA, PCR products around the target site were heat denatured and slowly annealed to allow the formation of heteroduplex DNA. We speculated that heat shock procedure generated three kinds of reannealed PCR products (Fig. 5b). The reannealed PCR products were digested by CEL II at 45 °C for 60 min, and three cleaved fragments were detected (Fig. 5c, lane 3). TALENs utilize the catalytic domain of FokI endonuclease and seem to cleave the spacer region between the target sites. Imprecise NHEJ causes the addition or deletion of nucleotides prior to ligation. Therefore, it is indeed difficult to estimate the accurate size of cleaved products. We speculated that the approximately 199-bp product was the diagnostic indicator of NHEJ, and the 160-bp products were derived from the fragments repaired by HR and NHEJ (Fig. 5c, lane 1 and 3). These results demonstrated that it appeared to be practical to detect mismatch heteroduplex DNAs using recombinant CEL II purified from silkworm larvae.
Discussion
The preparation of native enzymes usually requires several steps of purification, which demand significant time and labor. Pimkin et al. (2007) successfully purified recombinant CEL I using the baculovirus Sf9 insect cell expression system. Recently, insect larvae have been used as biofactories instead of cultured cells. The advantages of using insect larvae are the reduction in costs, the increase in recombinant protein yields, and the easy and inexpensive scale-up (Kato et al. 2009; Lee et al. 2012b). In this study, we produced recombinant CEL II from silkworm larvae using BES. The recombinant CEL II expressed in silkworm was N-linked glycosylated similarly to the native CEL II (Yang et al. 2000). Silkworm is known to have high-mannose or paucimannose-type glycans (Kost et al. 2005; Lee et al. 2012b). Although the detailed N-glycan structures of native and recombinant CEL II are unknown so far, the recombinant CEL II produced in silkworm larvae was functional in the mismatch cleavage assay. More than half of the CEL II proteins could not be secreted and remained in silkworm cells (data not shown). The protein processing pathway in the endoplasmic reticulum and Golgi network would be potential targets for modification to improve the recovery of recombinant proteins.
The purified CEL II could digest heteroduplex DNA efficiently, and its activity was strongly enhanced by heavy metal ions, calcium, and strontium. Crystal structures of P1 nuclease was determined, and P1 nuclease contains three zinc ions per monomer (Romier et al. 1998). Analysis of amino acids sequence indicated that all residues, which are important for binding of the three Zn2+ atoms, are well conserved in CEL II. Substitution of zinc ions by calcium or strontium ions is effective for mutation detection even though we have no idea to explain the results.
The heteroduplex DNA is usually incubated at 42 °C with the commercially available Surveyor nuclease (CEL II) according to the manual. In our assays, the optimum temperature for the nuclease activity was between 45 and 50 °C, which was similar to the best condition of Surveyor nuclease (Vogiatzakis et al. 2007). Several reports showed that the efficiency of CEL I mutation detection was increased with thermostable DNA polymerase or thermostable DNA ligase (Oleykowski et al. 1999; Yeung et al. 2005; Huang et al. 2012). The activity of CEL II mutation detection might be enhanced synergistically by either a polymerase or ligase enzyme.
We showed that the recombinant CEL II could detect SNPs and mutations generated by TALENs in vivo. The TALEN technology is becoming a powerful tool in gene editing, and highly reliable methods for detecting mutations would be indispensable. The CEL II expression system described here is easy to scale up for the larger amount of CEL II, which enables us to perform the high-throughput mutation detection assay.
References
Cermak T, Doyle EL, Christian M, Wang L, Zhang Y, Schmidt C, Baller JA, Somia NV, Bogdanove AJ, Voytas DF (2011) Efficient design and assembly of custom TALEN and other TAL effector-based constructs for DNA targeting. Nucleic Acids Res 39:e82–e82
Desai NA, Shankar V (2003) Single-strand-specific nucleases. FEMS Microbiol Rev 26:457–491
Huang MC, Cheong WC, Lim LS, Li M-H (2012) A simple, high sensitivity mutation screening using Ampligase mediated T7 endonuclease I and Surveyor nuclease with microfluidic capillary electrophoresis. Electrophoresis 33:788–796
Kato T, Kajikawa M, Maenaka K, Park EY (2009) Silkworm expression system as a platform technology in life science. Appl Microbiol Biotechnol 85:459–470
Kawaguchi Y, Kusakabe T, Lee JM, Koga K (2009) Characteristics of egg trait mutations in the silkworm, Bombyx mori. J Insect Biotechnol Sericology 78:113–126
Kawakami N, Lee JM, Mon H, Kubo Y, Banno Y, Kawaguchi Y, Maenaka K, Park EY, Koga K, Kusakabe T (2008) Efficient protein expression in Bombyx mori larvae of the strain d17 highly sensitive to B. mori nucleopolyhedrovirus. Mol Biotechnol 40:180–185
Kim HJ, Lee HJ, Kim H, Cho SW, Kim J-S (2009) Targeted genome editing in human cells with zinc finger nucleases constructed via modular assembly. Genome Res 19:1279–1288
Kost TA, Condreay JP, Jarvis DL (2005) Baculovirus as versatile vectors for protein expression in insect and mammalian cells. Nat Biotechnol 23:567–575
Lee JM, Kawakami N, Mon H, Mitsunobu H, Iiyama K, Ninaki S, Maenaka K, Park EY, Kusakabe T (2012a) Establishment of a Bombyx mori nucleopolyhedrovirus (BmNPV) hyper-sensitive cell line from the silkworm e21 strain. Biotechnol Lett 34:1773–1779
Lee JM, Mon H, Banno Y, Iiyama K, Kusakabe T (2012b) Bombyx mori strains useful for efficient recombinant protein production using a baculovirus vector. J Biotechnol Biomaterial
Lee JM, Mon H, Takahashi M, Kawakami N, Mitsunobu H, Banno Y, Koga K, Uchino K, Kawaguchi Y, Kusakabe T (2007) Screening of high-permissive silkworm strains for efficient recombinant protein production in Autographa californica nuclear polyhedrosis virus (AcNPV). J Insect Biotechnol Sericology 76:101–105
Mon H, Lee J, Kawaguchi Y, Kusakabe T (2011) Double-strand breaks repair by gene conversion in silkworm holocentric chromosomes. Mol Genet Genomics 286:215–224
Nishi O, Hasegawa K, Iiyama K, Yasunaga-Aoki C, Shimizu S (2011) Phylogenetic analysis of Metarhizium spp. isolated from soil in Japan. Appl Entomol Zool 46:301–309
Oleykowski CA, Bronson Mullins CR, Chang DW, Yeung AT (1999) Incision at nucleotide insertions/deletions and base pair mismatches by the SP nuclease of spinach. Biochemistry 38:2200–2205
Ono C, Nakatsukasa T, Nishijima Y, Asano S-I, Sahara K, Bando H (2007) Construction of the BmNPV T3 bacmid system and its application to the functional analysis of BmNPV he65. J Insect Biotechnol Sericology 76:161–167
Pimkin M, Caretti E, Canutescu A, Yeung JB, Cohn H, Chen Y, Oleykowski C, Bellacosa A, Yeung AT (2007) Recombinant nucleases CEL I from celery and SP I from spinach for mutation detection. BMC Biotechnol 7:29
Romier C, Dominguez R, Lahm A, Dahl O, Suck D (1998) Recognition of single-stranded DNA by nuclease P1: high resolution crystal structures of complexes with substrate analogs. Proteins 32:414–424
Tatsuke T, Hong S, Tobata H, Mon H, Lee J, Kawaguchi Y, Kusakabe T (2009) Construction of piggyBac-based vectors using visible and drug-resistance marker for introducing foreign genes into silkworm cultured cells. J. Fac. Agr. Kyushu Univ 54:397–400
Tsuji T, Niida Y (2008) Development of a simple and highly sensitive mutation screening system by enzyme mismatch cleavage with optimized conditions for standard laboratories. Electrophoresis 29:1473–1483
Vogiatzakis N, Kekou K, Sophocleous C, Kitsiou S, Mavrou A, Bakoula C, Kanavakis E (2007) Screening human genes for small alterations performing an enzymatic cleavage mismatched analysis (ECMA) protocol. Mol Biotechnol 37:212–219
Yamashita J, Miyagawa Y, Sugahara R, Mon H, Mitsunobu H, Lee JM, Kawaguchi Y, Kusakabe T (2007) Molecular cloning of silkworm Cdc37 and its interaction with Hsp90 chaperone. J Insect Biotechnol Sericology 76:137–143
Yang B, Wen X, Kodali NS, Oleykowski CA, Miller CG, Kulinski J, Besack D, Yeung JA, Kowalski D, Yeung AT (2000) Purification, cloning, and characterization of the CEL I nuclease. Biochemistry 39:3533–3541
Yeung AT, Hattangadi D, Blakesley L, Nicolas E (2005) Enzymatic mutation detection technologies. Biotechniques 38:749–758
Acknowledgments
We thank Dr. Susumu Shimizu for the gift of reagents. This work was supported in part by a grant from the Ministry of Agriculture, Forestry and Fisheries of Japan (Integrated Research Project for Plant, Insect and Animal using Genome Technology INSECT-1201), and KAKENHI nos. 22248003, 22248004, and 23580077 from the Japan Society for the Promotion of Science.
Conflict of interest
None.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Mon, H., Lee, J., Fukushima, M. et al. Production and characterization of the celery mismatch endonuclease CEL II using baculovirus/silkworm expression system. Appl Microbiol Biotechnol 97, 6813–6822 (2013). https://doi.org/10.1007/s00253-012-4583-1
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00253-012-4583-1