
Abstract
The study was undertaken to unravel the culturable endophytic bacterial microbiome of Crocus sativus L. (saffron crocus) and consequently obtain potential leads to develop plant growth-promoting and biocontrol agents for increased productivity and sustainable cultivation. The endophytes formed 47 different operational taxonomic units (OTUs), spanning over 28 genera. The host was preferentially colonized by the genus Bacillus, followed by Burkholderia and Pantoea, respectively. Several endophytes possessed potential plant growth-promoting properties and inhibitory activities against the specific fungal pathogens of saffron. The endophytes, except for Microbacterium oxydans, did not cause any disease symptoms in the pot experiments. The selected cultures, Burkholderia gladioli, Streptomyces achromogenes, and three species of Bacillus, enhanced the host plant growth significantly. Based on the pot experiment results, two isolates, Bacillus mojavensis CS4EB32 and Burkholderia gladioli E39CS3, were selected for the field experiments. We obtained an increase of 67.5%, 69.8%, and 68.3% in the production of flowers with the individual and collective treatments, respectively. The treatments also enhanced the biomass of the plant and the length and weight of stigmas significantly. The endophyte treatments induced the expression of the pathway genes, resulting in a marked increase in the concentration of apocarotenoids. The study indicates that the dominant endophytes support plant growth and development in nature and present an opportunity for developing microbial formulations for the sustainability of saffron cultivation.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Saffron (Crocus sativus Linn.) is grown in several parts of the world, for centuries, in specific agro-climatic conditions, mainly in Spain, Iran, and India [1]. The stigmata of its crocuses are used as a valuable spice, and an important ingredient of traditional medicine owing to its unique constituents called apocarotenoids — crocin, picrocrocin, and safranal — that lend it the peculiar color, flavor, and aroma [2]. Thus, it is regarded as one of the most expensive spices worldwide [3]. The importance and demand of this medicinal herb have increased significantly due to the recent findings that saffron and its constituents, particularly crocin, are potential antioxidant, anticancer, antidepressant, gastro-protective, and neuroprotective agents [4]. However, the production and cultivation of saffron have declined worldwide for the last few decades due to various factors [5, 6]. The crop may be severely affected by low temperatures, drought, water-logging, and fungal pathogens, particularly, Fusarium oxysporum, Alternaria alternata, Epicoccum nigrum, Penicillium pinophilum, and Talaromyces cellulolyticus [7,8,9]. Therefore, it is essential to devise strategies for its disease management, stress tolerance, increased productivity, and sustainable cultivation.
Endophytes are microorganisms, bacteria as well as fungi, that colonize the internal plant tissues without producing any harmful effects or eliciting an immune response in the host [10]. The bacterial endophytes, exhibiting plant growth-promoting traits like the production of indole-3-acetic acid (IAA) and regulation of the levels of ethylene by 1-aminocyclopropane-1-carboxylic acid (ACC) deaminase, have been extensively reported [11]. Similarly, endophytic bacteria provide nutrients to the plants, like nitrogen, phosphorous and iron through nitrogen fixation, phosphate solubilization, and siderophore production, respectively [12, 13]. Moreover, bacterial endophytes may reduce the incidence of infections in plants through the production of bioactive metabolites, competition for space or nutrients, and the development of systematic resistance [14]. The bacterial endophytes also protect the plants from abiotic stress conditions [15]. Thus, from an agronomical perspective, bacterial endophytes have emerged as an important biological resource to enhance plant growth and control specific plant pathogens, thereby increasing the productivity and yield in several plants [16, 17]. Therefore, it is necessary to characterize the endophytic bacterial diversity of a particular host plant to develop sustainable cultivation strategies.
Previously, we reported the community structure of the endophytic fungal microbiome of C. sativus and reported thirteen new potential fungal pathogens of the plant [8]. Consequently, we also found that one of its endophytes, Mortierella alpina CS10E4, enhances plant growth, apocarotenoid content, and stress tolerance in saffron crocus [18]. In the current study, we characterized another vital component of the endophytic microbiome, the bacterial microbiome, and have identified the potential role of selected endophytes in regulating the plant growth, development, and secondary metabolism.
Materials and Methods
Biological Materials and the Study Sites
For the isolation and analysis of the endophytic bacterial population, saffron plants were collected from four locations—Pulwama (33° 58′ 33.9″ N, 74° 56′ 16.1″ E), Srinagar (34° 01′ 57.1″ N, 74° 47′ 56.7″ E), Budgam (33° 51′ 46.4″ N, 74° 45′ 58.6″ E), and Kishtwar (33° 20′ 22.0″ N, 75° 44′ 45.1″ E) districts of Jammu and Kashmir, India (Fig. S1). Disease-free plants were collected in August and December, 10 plants each, from each location and stored at 4 °C.
Purification of Bacterial Endophytes
The bacterial endophytes were retrieved from the corms and the shoots of C. sativus with modifications of a previously described protocol suitable for bacterial isolation [8]. The plant tissues were washed with 1% sodium hypochlorite (Himedia, Mumbai, India) and 70% ethanol (Merck, Darmstadt, Germany) for 5 and 2 min, respectively. Excess amounts of disinfectants were removed by three washes of sterile distilled water and surface-drying in a biosafety hood. Samples from the final wash were plated on nutrient agar (NA) and potato dextose agar (PDA) and incubated for 48–72 h to confirm the efficiency of surface sterilization. The surface-sterilized tissues were then snipped into smaller segments of approximate ~ 0.5 cm and transferred to NA, actinomycete isolation agar (AIA; Himedia, Mumbai, India), water agar (WA), and Kenknight-Munaier’s medium [19] and incubated at 25 °C for 1–2 weeks. Individual bacterial colonies were purified and preserved. The cultures were deposited in the microbial repository of the institute (WDCM 1117). The bacterial isolates were grouped based on morphological characteristics like colony color, texture, shape, size, surface, edge, opaqueness, and the Gram’s reaction. The endophytic bacterial isolates were clustered into 107 different morphotypes based on morphological characteristics, and typical isolates of each morphotype were identified by the 16S rRNA gene sequence analysis (Table S1).
Genomic DNA Extraction, Acquisition of 16S rRNA Gene Sequences, and Phylogenetic Characterization
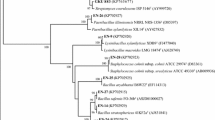
The genomic DNA of bacteria (gDNA) was extracted with Mericon DNA Bacterial plus kit (Qiagen, Hilden, Germany). The universal primers, 8F and 1492R, PCR reaction assay, and the thermal cycling program were used as described previously [20]. The PCR products were cleaned by QIAquick® PCR purification kit (Qiagen, Hilden, Germany) and custom-sequenced by the Eurofins Genomics, Bangalore, India, with both the primers. After assembling the sequences, the best-matching 16S rRNA gene sequences were determined by the BLAST analysis [21]. A phylogenetic tree was generated by the neighbor-joining method in MEGA (v 6) from closely related nucleotide sequences [22]
Diversity Analyses of Culturable Endophytes
The colonization frequency (CF) was calculated as the total number of plant segments colonized by endophytic bacteria divided by the total number of incubated plant segments, and the relative abundance, and species richness (S) were estimated as described previously [23]. The comparison of the bacterial diversity in different tissues and the comparative bacterial endophytic diversity at these sites was done using PAST (Version 3.04), [24].
In Vivo Test for Pathogenicity
The cultures were grown overnight at 25 °C in 15 mL nutrient broth (NB). Bacterial suspensions of 1.5 × 108 CFU mL−1 were prepared in normal saline solution [NSS, 0.85% NaCl (w/v)], used for the treatment of corms in all the experiments unless otherwise stated. Inoculums were mixed with 2% polyvinyl pyrrolidine (PVP, Sigma Aldrich, MO, USA), for coating. Autoclaved pot soil was spiked with 1.5 × 106 CFU g−1 cells of each culture. Surface-sterilized and injury imparted corms treated with F. oxysporum CSE15 were used as a positive control, whereas the negative control consisted of corms treated with NSS. Three corms per pot were used for each endophyte treatment and the experiment was carried out in triplicates. The corms were observed for symptoms of corm rot at 15 and 30 days post-inoculation (dpi) for the calculation of the Disease Index (D.I.) [8].
Evaluation of the Bacterial Endophytes for Plant Colonization Traits
The endophytes were screened for the plant cell wall degrading enzymes like lipase amylases, cellulases, and proteases [25]. Starch agar was used for the detection of amylase activity. Following bacterial inoculation and incubation at 25 °C for 72 h, the cultures were over-flown with Gram’s iodine. Amylase activity was recorded as the appearance of clear halos around the colonies. To test the cellulase activity of bacterial isolates, Czapek agar (Himedia, Mumbai, India) was supplemented with 0.5% (w/v) carboxymethylcellulose (CMC) sodium salt (Sigma Aldrich, MO, USA). The bacterial isolates demonstrating cellulase activity were visualized by treating the colonies with 0.2% congo red for 15 min followed by rinsing in 1 M NaCl twice to de-stain for 15 min. Colonies producing transparent hydrolytic zones were regarded as positive for cellulase production. For protease activity, skimmed milk agar (Himedia, Mumbai, India) plates inoculated with the cultures were placed at 25 °C for 72 h, and the development of clear halos around the colonies was taken as a positive reaction. Lipase activity was evaluated by the method of Sierra [26]. After incubation for 72 h at 25 °C, a visible halo formation was considered a positive test.
Assessment of the Bacterial Endophytes for Plant Growth-Promoting (PGP) Properties
The bacteria were cultured in 5 mL of NB overnight in a shaking incubator with 180 rpm at 25 °C. Five microliters of the culture were used in all the assays unless otherwise stated. IAA was quantified spectrophotometrically [27]. ACC deaminase activity was quantitatively measured by adopting the method of Penrose and Glick [28]. Further, endophytes were evaluated for their ability to fix nitrogen or survive on nitrogen-deficient medium [29], ammonia production [30], and inorganic phosphate solubilization [31]. Before the media preparation, all glassware was cleaned with 6 M HCl to remove any trace amounts of nitrogen-containing compounds. The endophytes were also evaluated for siderophore production [32].
Assessment of Antifungal Potential Against the Fungal Pathogens of Saffron
Chitinase activity was evaluated in a synthetic medium containing 0.2% (w/v) dry colloidal chitin powder [33]. Hydrogen cyanide production by the endophytes was assessed as described previously [34]. Further, the antifungal potential against the fungal pathogens of saffron as biocontrol agents was evaluated by the co-culture plate assay [8]. The fungal pathogens of saffron analyzed were Fusarium oxysporum CSE15, Fusarium oxysporum R1, Alternaria alternata CSE18, Talaromyces cellulolyticus CS3E6, Talaromyces pinophilus CSE29, Porostereum sp. CSE26, Penicillium pinophilum CSE20, and Acremonium sp. CSF4 [8].
Greenhouse Trial
Treatment of Saffron Corms with the Endophytes
Before the treatment, the corms were surface-sterilized [8]. Each endophyte suspension was mixed with 2% PVP as a coating agent. The surface-sterilized corms were submerged into the bacterial suspension for 2 h with continuous shaking. The endophyte-coated corms were transferred to the pots containing 100 g of autoclaved soil each and incubated in a greenhouse under the natural photoperiod of 10 h day/14 h night cycle at 25 °C. NSS-treated corms were sown in the soil as the control. We used one corm in each pot with five replicates for all the treatments for each endophyte treatment. An aliquot of bacterial suspension was inoculated at the time of corm sowing, while a similar inoculum was introduced after 7 days. In the control treatments, 1 mL of autoclaved NSS was used instead. The corms were sown in September 2018, and the saffron plants were harvested in the vegetative phase in January 2019.
Estimation of the Flavonoid and Phenolic Content
Briefly, 5 mL of 80% methanol was added to 1 g of tissue macerated in liquid nitrogen, followed by incubation with 37 °C with continuous shaking for 8 h. Extracts were prepared thrice from the same tissue, filtered, lyophilized, and finally dissolved in methanol. The total flavonoid and phenolic contents were measured using the aluminum chloride and the Folin–Ciocalteu reagent, respectively [35, 36].
Field Experiments
Bacterial Compatibility Assay
The compatibility of the selected endophytes developed as a consortium was evaluated by co-culture on NA medium. Each endophyte was placed on the agar surface. The strains were inoculated 0.5 cm apart and allowed to grow at 25 °C for 72 h. The colony interfaces were visually observed for growth inhibition.
Inoculum Preparation and Treatment of Saffron Corms with the Endophytes
The field experiments were carried out in collaboration with GloBiL’s Agri and Food Enterprises, our industrial partner, in their farms at Lassipora, Pulwama, J&K, India. The inoculums were prepared, and the corms were treated as described above. The consortium was developed by adding an equal number of cells of both endophytes. The corms were planted on raised (7 inches) beds of 4.3 × 1.5 m for each treatment (Fig. S2). Each bed was treated with 2.5 g NPK (DuraTec® Top 12, COMPO EXPERT, Germany) and 2.5 kg manure (Vermicompost), before plantation. Each treatment group, planted in a single bed, consisted of 277 corms with an average weight of 12.8 g. For the negative control, corms were treated with NSS, and the rest of the treatments were the same. The corms were sown in September 2019 and the flowers were collected in October and November 2019. However, flowering was stopped abruptly due to heavy and unprecedented snowfall on 6th November 2019, after which no flowers were collected.
Influence of Endophytes on Plant Growth, and Chlorophyll Content in the Host
Various morphological parameters like the number of flowers, fresh weight and length of stigmata, plant height, and fresh biomass, number and length of roots, the number of apical buds, and the number of daughter corms produced were recorded. A total of thirty saffron plants were harvested in the vegetative phase in December 2019 for the analyses of morphological data. The total chlorophyll content was estimated using a previously described method [37].
Quantification of the Apocarotenoids
Crocin was extracted from stigmata as described previously [18]. For the quantification safranal, 100 mg of dried stigmas were crushed with the help of liquid nitrogen and dissolved in 1 mL of chloroform. The mixture was placed in an ice bath for 10 min and centrifuged at 5000 g for 5 min in a refrigerated centrifuge. The supernatant was evaporated to obtain the extracts, dissolved in methanol, and analyzed by HPLC [18]. Standard markers of crocin and safranal were procured from Sigma Aldrich.
Comparative Expression by Quantitative Reverse Transcriptase PCR Analyses
The RNA was isolated from crocus flowers using RNeasy® Plant Mini Kit (Qiagen, Hilden, Germany) and the complementary DNA were synthesized by the Easy script™ plus kit (Applied Biological Materials Inc. Richmond, BC, Canada). Three replicates were obtained and the qRT-PCR assays were run in an Mx3000p QPCR System (Stratagene). The reaction mixture consisted of 5 μL of 2 × KAPA SYBR FAST Master Mix, 0.2 pmol primers, and 100 ng of the template cDNA in a total assay of 10 μL. The thermal conditions were 95 °C for 3 min, 40 cycles of 95 °C for 15 s, and 60 °C for 1 min. The melting curves were obtained by collecting fluorescence from 60 to 95 °C. The 18S rRNA gene was used as an endogenous normalizing gene. The relative gene expression was estimated using the 2−ΔΔCt method [38]. The sequences of the oligonucleotide primers are given in Table S2.
Statistical Analyses
The statistical analyses were carried out by GraphPad Prism version 5.00 (GraphPad Software, San Diego, CA, USA). The data from the greenhouse pot and the field experiments were analyzed with one-way ANOVA and Bonferroni’s tests. The data on gene expression were evaluated by two-way ANOVA with Bonferroni’s tests and a heatmap was generated using the NCSS 2007 version 07.1.14 software. At a probability level of p < 0.05, the differences between the treatments were regarded as significant. ***, **, *, and ns indicate significant difference at p < 0.001, p < 0.01, p < 0.05, and non-significant, respectively, with reference to the untreated control.
Results
Phylogenetic Affinity and Diversity of Bacterial Endophytes Retrieved from C. sativus
A collection of 306 cultures of bacterial endophytes were purified from 2408 fragments of corms and shoot from four different sites. Precisely, 203 isolates from the corms and 103 from the shoot were recovered and stored. The analyses of these endophytes from the saffron tissues for different locations are presented in Fig. S3a. The colonization frequency was higher in the corm tissues (54.6%) relative to the shoot tissues (30.9%) (Fig. S3b). The colonization frequency of the endophytes varied from 35.8 to 48.2% at the four different locations (Fig. S3c). The 16S rRNA gene sequence analyses distributed the 107 morphotypes into 47 different OTUs comprising 28 genera, identified at a threshold of 99% sequence similarity (SS) except the OTU 19 (Table 1). These 47 different OTUs were used for further diversity analysis and plant growth-promoting properties.
The representative OTUs were assigned to Proteobacteria, Firmicutes, Actinobacteria, and Bacteroidetes. Proteobacteria was the most dominant phylum consisting of about 51% of the OTUs. Firmicutes constituted the second-largest phylum followed by the Actinobacteria and Bacteroidetes with about 36.1%, 10.6%, and 2.0% OTUs, respectively (Fig. S4). The most dominant genus within the bacterial endophytes was Bacillus (66 independent isolates), followed by Burkholderia (57 independent isolates) and Pantoea (20 independent isolates). Several other genera had more than three independent isolates while two genera possessed singletons (Fig. 1a). Bacillus and Burkholderia were shared by all four locations, occupying 40.1% of all the bacterial genera (Fig. S3d). Both the genera were also widely distributed in the corms as well as the shoot tissues of the host. The Venn diagram also depicted that the number of unique bacterial genera in each location was in low abundance.

The diversity of the culturable bacterial endophytes of C. sativus. a The relative abundance of 28 different genera that represent the endophytic bacterial community of saffron crocus. b The diversity profile graph of the bacterial endophytes of C. sativus at the tissue level. The endophytes were more diverse in the the corms than the shoot. c The diversity analyses of saffron showed a higher colonization frequency in the corm than the shoot. A total of 13 bacterial genera were specific to the corm, whereas only 4 bacterial genera were specific to the shoot of the plant. The remaining 11 genera were found in both the corm and shoot tissues
The level of bacterial diversity differed significantly between the two tissue types. The Simpson, Shannon, and Chao1 observed values for the corm tissues were 0.89, 2.69, and 24.33 and for the shoot tissues, 0.87, 2.31, and 16.5, respectively. The species richness (S) was also higher in the corms. These findings are also evident in the diversity profile graph (Fig. 1b). A total of 13 bacterial genera, Enterobacter, Microbacterium, Paraburkholderia, Xanthomonas, Streptomyces, Obesumbacterium, Sphingomonas, Methylobacterium, Micrococcus, Alcaligenes, Lysinibacillus, Paenibacillus, and Flavobacterium, were specific to the corms, whereas only four bacterial genera, Klebsiella, Erwinia, Pseudomonas, and Kocuria, were specific to the shoot tissues. The remaining 11 genera were common to both the corm and shoot tissues (Fig. 1c).
Further, the diversity of endophytes as estimated by Simpsons, Shannon, and Chao1 diversity indices (Table S3) indicated that it was similar in all the locations but slightly lower at the location 4. The diversity profile graph also exhibits similar patterns of diversity at these locations (Fig. S3e).
Bacterial Endophytes Do not Cause Corm Rot in Saffron
Bacterial endophytes from the C. sativus were tested to determine whether any of the endophytes could cause any symptom disease (corm rot) on inoculation in healthy saffron corms. We found that none of the bacterial endophytes caused corm rot except Microbacterium oxydans E108CS6, which induced some visible corm rot symptoms with a low D.I. value of 0.66 ± 0.16. However, the known pathogen, F. oxysporum, used as a positive control, caused the disease in all the corms with a D.I. value of 2.7 ± 0.28 (Table S4).
Bacterial Endophytes Show Plant Colonization Traits
A total of 21 isolates produced extracellular proteases. Among these isolates, 11 produced > 10 mm clearance zones on skimmed milk agar. Additionally, 24 isolates produced lipases, 14 isolates produced amylases, and 16 cultures produced the cellulase enzymes (Table 2).
Bacterial Endophytes Exhibit In Vitro Plant Growth Promotion
PGP properties tested included nitrogen fixation, phosphate solubilization, the production of IAA and ACC deaminase, ammonia, and siderophores (Table 2). All the 47 endophytic strains produced IAA under axenic conditions. However, out of the 47 endophytic strains, 9 strains produced more than 100 mg L−1 of IAA in liquid broth. These strains included Pantoea eucalypti E62CS3, Pantoea conspicua E66CS3, Obesumbacterium proteus E92CS4, Ba. megaterium E67CS3, K. oxytoca E105CS6, R. aquatilis E48CS3, Paraburkholderia phenazinium CSEB1, Ba. stratosphericus E96CS5, and A. xylosoxidans CSEB4. Additionally, 8 bacterial strains produced 50–100 mg L−1 IAA, whereas most bacterial endophytes (63.8%) produced < 50 mg L−1 IAA. Eleven bacterial isolates (23.4%) produced the ACC deaminase enzyme. The quantity of α-ketobutyrate released from the ACC substrate indicated the ACC deaminase production, ranging between 5.0 and 60.2 μmol α-KB mg−1 h−1. Three isolates, Erwinia persicina E127CS7, Bu. gladioli E39CS3, and Ba. megaterium E67CS3, produced the highest amounts of ACC deaminase, 60.2, 53.4, and 50.0 μmol α-KB mg−1 h−1, respectively.
A total of 35 endophytes out of the 47 OTUs were able to grow on a nitrogen-deficient medium, with 14 of them showing luxuriant growth. A total of 30 bacterial endophytes secreted ammonia into the medium in the in vitro conditions. The inorganic phosphate was solubilized by thirteen cultures, among which R. aquatilis E48CS3, Bu. gladioli E39CS3, and Obesumbacterium proteus E92CS4 were the most efficient (Table 2). Most of the isolates (68.0%) produced siderophores. Potential siderophore producers were Phyllobacterium ifriqiyense B2B8, Bu. gladioli E39CS, Paraburkholderia soli CSEB14, and Pantoea conspicua E66CS3.
Bacterial Endophytes Exhibit a Broad Range of Antifungal Activity Against the Fungal Pathogens of Saffron
Only two isolates, Bu. gladioli E39CS3 and S. marcescens CSEB46, could degrade colloidal chitin. Bu. gladioli E39CS3 was observed to possess higher chitinase activity as compared to S. marcescens CSEB46. Further, each strain of Pseudomonas produced HCN (Table 2).
Among the endophytes, a total of six, Ba. amyloliquefaciens E87CS4, Ba. mojavensis CS4EB32, Ba. siamensis E80CS4, S. achromogenes E91CS4, Ba. halotolerans E79CS3, and Bu. gladioli E39CS3, exhibited a broad range of antifungal activity against the fungal pathogen of saffron (Table S5). Ba. amyloliquefaciens E87CS4 and Ba. mojavensis CS4EB32 inhibited all the fungal pathogens by more than 50%. Ba. siamensis E80CS4 and S. achromogenes E91CS4 inhibited seven pathogens whereas Ba. halotolerans E79CS3 and Bu. gladioli E39CS3 inhibited six pathogens by more than 50% (Fig. S5). However, Bu. gladioli E39CS3 inhibited both Fusarium oxysporum strains CSE15 and R1 most efficiently.
Several Bacterial Endophytes Promote Plant Growth and Secondary Metabolite Content of C. sativus in the Greenhouse Pot Experiments
Five bacterial endophytes, Ba. siamensis E80CS4, Ba. halotolerans E79CS3, Bu. gladioli E39CS3, Ba. mojavensis CS4EB32, and S. achromogenes E91CS4, were selected for their potential to facilitate plant growth in the greenhouse. The treatments exhibited a noteworthy increase in growth parameters of the plant (Fig. 2a). The biomass and the height of the endophyte-treated plants increased significantly as compared to that in the control plants (Fig. 2b, c). The results indicated that the impact of the endophyte, Bu. gladioli E39CS3, Ba. mojavensis CS4EB32, and S. achromogenes E91CS4 on the plant rooting and shooting systems was significant. As a consequence, the length of the adventitious roots and their numbers increased in the endophyte-treated plants (Fig. 2d, e). Out of the five endophytes, Ba. halotolerans E79CS3 and Ba. mojavensis CS4EB32 significantly increased the number of apical buds sprouting per corm (Fig. 2f). However, the biomass of the corms increased only with the treatment of Bu. gladioli E39CS3 to an average of 5.3 g corm−1 in comparison to 2.0 g corm−1 for the untreated control (Fig. 2g). Besides, all the endophyte treatments significantly increased the flavonoid contents (Fig. S6a). However, there was no significant increase in the phenolic content except for the plants treated with Bu. gladioli E39CS3 (Fig. S6b).

The effect of the endophytes on the root and shoot development of C. sativus in the greenhouse experiments. a Impact of the endophytes, Ba. siamensis E80CS4, Ba. halotolerans E79CS5, Bu. gladioli E39CS3, Ba. mojavensis CS4EB32, and S. achromogenes E91CS4 on the root and shoot development. Plots illustrating the statistical analysis of the plant growth parameters such as b fresh biomass gain, c plant height, d length of adventitious roots, e number of adventitious roots per corm, f number of apical buds sprouting per corm, and g biomass of the corms. Values are the means of 5 biological replicates ± S.D
Bacterial Endophytes Promote Early Flowering and Root and Shoot Development in the Field Experiments
In the compatibility assay, Ba. mojavensis CS4EB32 and Bu. gladioli E39CS3 stains were able to grow without inhibiting each other when co-cultivated in vitro (Fig. S7). The cultures CS4EB32, E39CS3, and the consortium of both (CS4EB32 + E39CS3) were applied in the field conditions. Interestingly, a 10-day early flowering was observed in the plants treated with Bu. gladioli E39CS3 and the consortium, while 8-day early flowering was obtained in the plants seeded with Ba. mojavensis CS4EB32. The number of flowers obtained from the endophyte-treated plants increased exceptionally in comparison to the untreated plants. A total of 211, 214, and 212 flowers were obtained from CS4EB32, E39CS3, and the consortium treatments, respectively, in contrast to the control, which produced 126 flowers (Fig. 3a). This indicates an increase of 67.5%, 69.8%, and 68.3% in the production of flowers in the plants treated with CS4EB32, E39CS3, and the consortium, respectively. However, there was an unprecedented heavy snowfall in the Kashmir valley in the first week of November, which arrested the flowering abruptly.

The effect of the endophytes on the flowering characteristics of C. sativus. a The number of flowers obtained, b the length of stigma, c fresh weight of stigma, and d pictorial view of stigma. The values are the means of 30 biological replicates ± S.D
The treatments increased the length and the fresh weight of the stigmas significantly, with E39CS3 producing the maximum increase of 26.2% and 45.6%, respectively (Fig. 3b-d).
The aerial parts of the endophyte inoculated plants also displayed a marked increase in the growth parameters (Fig. 4a), such as height (Fig. 4b) and fresh biomass (Fig. 4c). Further, a remarkable improvement was visible in the rooting systems like the length (29.6–79.6%) and number of adventitious roots (61.4–82.7%) (Fig. 4d, e). The number of apical buds and secondary cormlets increased significantly only in the E39CS3 treatments relative to un-inoculated control.

The effect of the endophytes on the root and shoot development of C. sativus in the field experiments. a Impact of the endophytes, Bu. gladioli E39CS3, Ba. mojavensis CS4EB32, and a consortium of both on the root and shoot development. The endophytes visibly improved the root and shoot growth in the plants, b fresh biomass gain, c plant height, d length of the adventitious roots, e number of adventitious root per corm. Values are the mean of 30 biological replicates ± S.D
Bacterial Endophytes Improve the Secondary Metabolite Content in the Host Plant
During the vegetative phase, we observed that the endophytes resulted in increased total flavonoid concentration of the plant in all the treatments significantly. However, the total phenolic content increased significantly only with E39CS3 treatments. The flavonoid content increased more than 10% in all the endophyte treatments (Fig. 5a), whereas the phenolic content increased more than 10% only with the E39CS3 treatments (Fig. 5b). Further, the total chlorophyll content increased significantly in E39CS3 and the consortium-treated plants (Fig. 5c).

The effect of the endophytes on important secondary metabolites of C. sativus in the field experiments. Impact of the endophytes Ba. mojavensis CS4EB32, Bu. gladioli E39CS3, and the consortium on the a flavonoid content, b phenolic content, c total chlorophyll content, d crocin content, e safranal content. The values are the mean of 3 biological replicates ± S.D
Bacterial Endophytes Enhance the Apocarotenoid Content in Saffron
The HPLC quantification of crocin in the stigmas revealed that E39CS3 and the consortium treatments increased the crocin content by 69.3% and 47.3%, respectively (Fig. 5d). In contrast, CS4EB32 did not influence the production of crocin, significantly. Further, safranal content was found to increase by 246.8%, 102.2%, and 155.7% in E39CS3, CS4EB32, and the consortium-treated plants (Fig. 5e).
Endophyte Treatments Induce the Expression of the Key Pathway Genes
To understand the mechanism of increased production of apocarotenoid by the endophyte treatments, expression of key genes involved in the carotenoid and apocarotenoid synthesis was measured by qRT-PCR. The expression profiles of ten important genes, PSY, PDS, ZDS, CRTISO, LYC, CCD4a, BCH2C, CCD2, ALDH, and UGT, were analyzed. The expression of the carotenoid and apocarotenoid pathway genes was upregulated in the treated plants (Fig. 6a, b). The mRNA levels of the phytoene synthase (PSY) gene, which carries out the first step in the carotenoid biosynthesis, showed 8.7, 2.2, and 4.4 fold increase in the E39CS3, CS4EB32, and the consortium inoculated plants, respectively. The intermediate pathway genes of carotenoid biosynthesis like phytoene desaturase (PDS), zeta carotene desaturase (CaZDS), carotenoid isomerase (CRTISO), and lycopene cyclase (LYC) also showed upregulation in all the endophyte-treated plants. Further, the beta-carotene hydroxylase gene (BCH2C), which performs β-carotene hydroxylation and generates zeaxanthin, the precursor of apocarotenoids (crocin, picrocrocin, and safranal), showed 6.5, 1.4, and 6.7 fold induction by E39CS3, CS4EB32, and the consortium treatments, respectively. The CCD2 gene responsible for cleavage of zeaxanthin into hydroxyl β-cyclocitral and crocetin dialdehyde showed 2.9, 2.2, and 1.3 fold increase, whereas the gene for CCD4a that catalyzes the breakdown of beta-carotene into beta-ionone and cyclocitral showed 5.4, 7.4, and 2.4 fold increase in CS4EB32, E39CS3, and the consortium treatments, respectively. Further, the aldehyde dehydrogenase gene (ALDH), which converts crocetin dialdehyde into crocetin, showed 11.2, 1.4, and 6.7 fold induction in E39CS3, CS4EB32, and consortium treatments, respectively. Finally, the expression of UDP-glucosyltransferases gene (UGT) which converts hydroxyl-β-cyclocitral into picrocrocin, the immediate precursor of safranal, showed 3.6, 2.9, and 4.0 fold increase in E39CS3, CS4EB32, and the consortium treatments, respectively.

The effect of the endophytes on gene expression of C. sativus in the field experiments. a Influence of the endophyte Ba. mojavensis CS4EB32, Bu. gladioli E39CS3, and the consortium on important carotenoid and apocarotenoid biosynthesis genes. Relative gene expression of phytoene synthase (PSY), phytoene desaturase (PDS), zeta carotene desaturase (CaZDS), carotenoid isomerase (CRTISO), lycopene cyclase (LYC), carotenoid cleavage dioxygenase 4 (CCD4a), beta-carotene hydroxylase (BCH2C), carotenoid cleavage dioxygenase 2 (CCD2), aldehyde dehydrogenase (ALDH), and UDP-glucosyltransferase (UGT). b The heatmap showing the expression profile of the carotenoid and apocarotenoid biosynthetic pathway genes in different treatments. The significantly upregulated genes are highlighted in red and yellow colors and the non-significant changes in gene expression are represented in the blue color. The values are the mean of 3 biological replicates ± S.D
Discussion
Despite the importance of C. sativus as a medicinal plant, an exotic spice, and a precious cash crop, its endophytic bacterial community is largely unexplored. Considering the ecological roles of endophytes in plant health, yield, mitigation of environmental stresses, and diversification, it is imperative to characterize the host microbiomes for sustainable cultivation of saffron [12, 14]. Therefore, we characterized the endophytic bacterial microbiome of the saffron plant through culture-dependent methods and obtained leads for the enhancement of growth and yield of this crop.
The results indicate that bacterial endophytes more abundantly colonized corm tissues compared to the shoot tissues. As the corms are fleshy, nutrient-rich, and underground plant parts, the conditions are favorable for the colonization of endophytes. However, compared to the corms, shoot development occurs at the short vegetative phase of the saffron life cycle, thus supporting the growth of fewer endophytes [1]. Also, shoots are exposed to stronger physicochemical variations, which generally reduce the colonization of endophytes [39]. Various reports suggest that plant tissues impact the diversity and composition of endophytic communities due to their specificity and adjustment to different anatomical and physiological conditions of the plant micro-environments [10, 40]. As saffron is propagated through corms, the vertical transfer of endophytes for hundreds of years of cultivation and the horizontal transfer from the environment, especially soil, have specifically evolved its endophytic bacterial microbiome.
In other plants also Proteobacteria, Firmicutes, Actinobacteria, and Bacteroidetes were reported as the most dominant bacterial endophytic phyla [41]. The most prevalent species that makes an endophytic association with saffron crocus is Burkholderia gladioli, but the most dominant genus as its endophyte is Bacillus. Previously, six genera, Pseudomonas, Bacillus, Paenibacillus, Staphylococcus, Brevibacterium, and Enterobacter, were reported as the endophytes of C. sativus, but the sample size was minimal, and the plant samples were collected from a single site [42]. Here, we report twenty-two additional genera, including Burkholderia, the second most dominant genus, as the endophytes of C. sativus. The data suggest that C. sativus harbors a more diverse array of bacterial endophytes than that reported earlier. The core microbiome analysis of saffron showed that Bacillus and Burkholderia were shared among all the four locations, widely distributed in the corm and the shoot tissues. Thus, the association of Bacillus and Burkholderia with saffron may have a far-reaching influence on plant growth and development, as envisaged by this study. The core microbiome plays a critical role in plant development and distribution, as found in this case [43].
The type of plant tissues, physiology, geographical locations, and ecological factors influence the diversity and community structure of the endophytes of various plants [10]. The analyses across different sites suggested that because of the same environmental, climatic conditions, and genetic factors of the host, the diversity of the endophytic bacteria was almost uniform at all the locations. The results align with our study on the fungal endophytes of C. sativus from the same locations [8].
Plant-associated endophytes are beneficial for the plants, but some endophytes can act as latent or opportunistic pathogens [44]. During in vivo pathogenicity tests, we observed that none of the culturable endophytes showed corm rot in the saffron corms except a mild rot caused only by M. oxydans. The corm rot in saffron is primarily caused by fungal pathogens, particularly Fusarium oxysporum [7, 8]. Endophytes possess potential sustainable agricultural applications as they can significantly contribute to plant health, productivity, and stress tolerance [11, 14]. Bacterial endophytes regulate plant growth directly or indirectly by enhancing nutrient availability, production of phytohormones, modulating the ethylene levels within the plants, etc. Cellulase and amylase enzymes may help them colonize in plant tissues due to their capability to degrade the cell wall components [12]. Bacterial proteases and lipases are also believed to suppress a wide range of plant pathogens, thus helping the plant evade phytopathogens [45].
Plant-associated bacteria use IAA to colonize the plant tissues [46]. We found that all bacterial endophytes produced IAA in vitro, though in varying concentrations. Previous studies on fungal endophytes of Crocus sativus and Glycyrrhiza glabra from our lab also reported that all the endophytes were IAA producers [8, 19]. Phytohormones increase water and nutrient uptake and enhance plant growth and development in the host plants [47]. Thus, the IAA produced by endophytes could play an important role in plant colonization and plant growth promotion. Plants tend to synthesize ethylene to stimulate fruit-ripening, and it also acts as a vital stress hormone to sense the onset of abiotic and biotic stresses. However, the second peak of ethylene is harmful to plant growth and development [48]. ACC deaminase produced by bacteria breaks down ACC, the precursor of ethylene, into α-ketobutyrate and ammonia, thus preventing its deleterious effects [15]. Therefore, the endophytes Bacillus, Burkholderia, Citrobacter, Enterobacter, Erwinia, Rahnella, and Serratia can protect the plant from abiotic stresses.
In this study, we found that most endophytes proliferated on nitrogen-deficient media and produced ammonia in the in vitro conditions, indicating that several of these isolates may be able to fix the atmospheric nitrogen. However, five bacterial endophytes growing on the nitrogen-free medium could not secrete ammonia, which is in conformity with the previous findings [49]. Similarly, a total of thirteen endophytic bacterial isolates exhibited phosphate solubilization activity. Interestingly, all the species belonging to Pantoea could mineralize the phosphate, which is supported by previous studies [50]. Besides, bacterial endophytes may facilitate iron acquisition through siderophore production, contributing to pathogen inhibition [51]. The majority of the endophytes (68.0%) produced siderophores, but Phyllobacterium ifriqiyense and Bu. gladioli were the most efficient producers.
The rot caused by Fusarium oxysporum is the most common disease in saffron resulting in severe losses in the yield [9]. In a previous study, we have also reported some other fungal pathogens of saffron capable of producing rot in corm [8]. Moreover, a recent study reported that the corm rot was also caused by Penicillium solitum [52]. The emerging literature suggests that bacterial endophytes possess enormous potential to control plant pathogenic fungi, thus protecting the host from fungal diseases [53, 54]. Bacterial endophytes may suppress the pathogens by the synthesis of hydrogen cyanide (HCN) and other allelochemicals, antifungal metabolites, chitinase, and through the induction of plant systematic resistance [14, 55]. We found that the endophytes belonging to the genus Burkholderia and Serratia produced chitinases. Chitinase-producing Bu. gladioli are also reported to control various fungal pathogens in other plants [56]. Several bacterial endophytes, including the genera Bacillus, Streptomyces, and Burkholderia, exhibited a broad range of antifungal activity against the specific fungal pathogen of saffron. However, Burkholderia and Bacillus demonstrated potential as biocontrol agents, which may be employed to manage the corm rot in the saffron crocus.
The aim of using plant growth-promoting bacterial endophytes in agriculture is to reduce the use of harmful fertilizers and pesticides [57]. Thus, the selected bacterial endophytes are potential candidates for the enhancement of plant growth of the host. In the field experiments, we found that the endophyte treatments had a more prominent effect on the number of flowers and length and weight of stigma, which improved the yield of saffron remarkably. The improvement of plant growth parameters should ultimately translate into higher yields, which was achieved perfectly in this study. Since the ultimate product is the stigma obtained from saffron plants, the yield further improved due to increased stigmata length and weight. If similar trends are obtained by these microbial formulations in large-scale field applications, it could have an unprecedented effect on the yield of saffron in Kashmir and worldwide. Such agrotechnologies can have an enormous influence on the sustainable cultivation of saffron and its productivity and thus may enable the farmers to meet the ever-growing demand for this valuable commodity. Consequently, it will also result in huge economic gains for saffron growers. Therefore, specific endophytes, selected based on plant growth-promoting traits, can increase the yields of a crop significantly. In plants, phenolic and flavonoids play multiple roles, including UV protection, defense mechanism, and antioxidant activity [58]. Thus, the endophytes help the plants to survive under stress conditions. All these results validate our pot experiment results in the field settings.
Interestingly, the inoculation of endophytes increased the crocin and safranal content due to the upregulation of the apocarotenoid biosynthetic pathway genes in Crocus plants in the field experiment. An increase in key secondary metabolite content of other medicinal plants by specific endophytes has been reported previously [59]. However, the expression profiling shows that each endophyte treatment differentially upregulates the transcription of key genes involved in the carotenoids and apocarotenoid biosynthesis. Phytoene synthase, which E39CS3 and the consortium treatments strongly upregulate, catalyzes the foremost step of carotenoid biosynthesis. Similarly, the intermediate carotenoid biosynthetic pathway genes also showed potential upregulation in both treatments. Further, BCH2C, involved in zeaxanthin production, the precursor of apocarotenoids of the saffron crocus (crocin, picrocrocin, and safranal), was also strongly induced by E39CS3 and the consortium treatments. The downstream core gene in the apocarotenoid pathway, ALDH, was also strongly upregulated in E39CS3 and the consortium. However, UGT was upregulated in all endophyte-treated plants. Thus, the upregulation of the carotenoid biosynthetic genes increases the production of crocin, picrocrocin, and safranal in the endophyte-treated plants.
The selection of two bacterial endophytes was rationalized because both strains are dominant endophytes of the C. sativus, having potential plant growth-promoting and antifungal activities. The choice of developing a consortium is to study the complementary or synergistic effect of endophytes on plant growth promotion and metabolism of C. sativus, as earlier reported by several researchers in different plants [60]. In our study, we found that all the treatments had an almost similar influence on the number of flowers, fresh weight, and length of stigma. However, Bu. gladioli E39CS3 had a more prominent effect in terms of potentiating the growth, secondary metabolites content, and expression levels genes of the apocarotenoid pathway compared to the consortium. Burkholderia species, including Bu. gladioli, have emerged as potential biostimulants and biocontrol agents in several plants [61]. However, Burkholderia spp. have also been reported as opportunistic human pathogens and causative agents of several plant diseases [62, 63]. However, recent advances in biological science, including the high throughput genome sequencing and detection of virulence genes, may be applied for a clear distinction between beneficial Burkholderia and its pathogenic strains [64]. Since Bu. gladioli is the most dominant endophyte of the saffron plant, its association may have significant implications on the plant growth and development, which is essentially the case as found in this study. Further, Bu. gladioli E39CS3 is a potential biocontrol agent as it induces host resistance against the major saffron pathogen, F. oxysporum [65]. However, it may be approved for regular use only after the safety of the strain is unambiguously established.
Conclusion
In this study, we explored the endophytic bacterial microbiome of the saffron plant. Although the endophytes consisted of diverse lineages, the host recruits several species of Bacillus and Burkholderia gladioli, preferentially as its symbiotic partners. The pot and field experiments demonstrated that several of these endophytes enhanced plant growth substantially. The selected endophytes, Ba. mojavensis, and Bu. gladioli increased the yield of the flowers significantly and also shifted the metabolic flux towards the synthesis of the key components of the host, the apocarotenoids. In this particular case, we found that the most dominant endophytes of the saffron plant demonstrated the most profound effects on plant health and yield. Thus, the saffron plant has evolved its endophytic microbiome to obtain the most beneficial traits for better survival and growth. It may be interesting to investigate if the same principle applies to other crops. These findings hold promise for the development of commercial formulations for increased productivity and sustainable cultivation of saffron. The association of these endophytes with the saffron plant may be further studied at the molecular level to understand the molecular mechanism of plant growth promotion.
Data Availability
The pure cultures were deposited in the microbial repository of the institute (WDCM 1117). The 16S rRNA gene sequence data were submitted to the GenBank under the accession numbers MK472702–MK472705, MK474934–MK475031, MK377246, MK419120, MK583724, MK621297, and MK621285.
Code Availability
Not applicable.
References
Kumar R, Singh V, Devi K, Sharma M, Singh MK, Ahuja PS (2008) State of art of saffron (Crocus sativus L.) agronomy: a comprehensive review. Food Rev Int 25:44–85. https://doi.org/10.1080/87559120802458503
Ahrazem O, Rubio-Moraga A, Nebauer SG, Molina RV, Gomez-Gomez L (2015) Saffron: its phytochemistry, developmental processes, and biotechnological prospects. J Agric Food Chem 63:8751–8764. https://doi.org/10.1021/acs.jafc.5b03194
Mzabri I, Addi M, Berrichi A (2019) Traditional and modern uses of saffron (Crocus Sativus). Cosmetics 6:63. https://doi.org/10.3390/cosmetics6040063
Bukhari SI, Manzoor M, Dhar MK (2018) A comprehensive review of the pharmacological potential of Crocus sativus and its bioactive apocarotenoids. Biomed Pharmacother 98:733–745. https://doi.org/10.1016/j.biopha.2017.12.090
Gresta F, Lombardo GM, Siracusa L, Ruberto G (2008) Saffron, an alternative crop for sustainable agricultural systems. A review Agron Sustain Dev 28:95–112. https://doi.org/10.1051/agro:2007030
Menia M, Iqbal S, Zahida R, Tahir S, Kanth RH, Saad AA, Hussian A (2018) Production technology of saffron for enhancing productivity. J Pharmacognos Phytochem 7:1033–1039
Palmero D, Rubio-Moraga A, Galvez-Patón L, Nogueras J, Abato C, Gómez-Gómez L, Ahrazem O (2014) Pathogenicity and genetic diversity of Fusarium oxysporum isolates from corms of Crocus sativus. Ind Crops Prod 61:186–192. https://doi.org/10.1016/j.indcrop.2014.06.051
Wani ZA, Mirza DN, Arora P, Riyaz-Ul-Hassan S (2016) Molecular phylogeny, diversity, community structure, and plant growth promoting properties of fungal endophytes associated with the corms of saffron plant: an insight into the microbiome of Crocus sativus Linn. Fungal Biol 120:1509–1524. https://doi.org/10.1016/j.funbio.2016.07.011
Cappelli C (1994) Occurrence of Fusarium oxysporum f. sp. gladioli on saffron in Italy. Phytopathol Mediterr 33:93–94. https://doi.org/10.1023/A:1021204022787
Compant S, Cambon MC, Vacher C, Mitter B, Samad A, Sessitsch A (2020) The plant endosphere world–bacterial life within plants. Environ Microbiol. https://doi.org/10.1111/1462-2920.15240
Santoyo G, Moreno-Hagelsieb G, del Carmen O-M, Glick BR (2016) Plant growth-promoting bacterial endophytes. Microbiol Res 183:92–99. https://doi.org/10.1016/j.micres.2015.11.008
Hardoim PR, Van Overbeek LS, Berg G, Pirttilä AM, Compant S, Campisano A, Döring M, Sessitsch A (2015) The hidden world within plants: ecological and evolutionary considerations for defining functioning of microbial endophytes. Microbiol Mol Biol Rev 79:293–320. https://doi.org/10.1128/MMBR.00050-14
Afzal I, Shinwari ZK, Sikandar S, Shahzad S (2019) Plant beneficial endophytic bacteria: mechanisms, diversity, host range and genetic determinants. Microbiol Res 221:6–49. https://doi.org/10.1016/j.micres.2019.02.001
Liu H, Carvalhais LC, Crawford M, Singh E, Dennis PG, Pieterse CM, Schenk PM (2017) Inner plant values: diversity, colonization and benefits from endophytic bacteria. Front Microbiol 8:2552. https://doi.org/10.3389/fmicb.2017.02552
Ali S, Charles TC, Glick BR (2014) Amelioration of high salinity stress damage by plant growth-promoting bacterial endophytes that contain ACC deaminase. Plant Physiol Biochem 80:160–167. https://doi.org/10.1016/j.plaphy.2014.04.003
Card S, Johnson L, Teasdale S, Caradus J (2016) Deciphering endophyte behaviour: the link between endophyte biology and efficacious biological control agents. FEMS Microbiol Ecol 92:8. https://doi.org/10.1093/femsec/fiw114
Harman GE, Uphoff N (2019) Symbiotic root-endophytic soil microbes improve crop productivity and provide environmental benefits. Scientifica 2019. https://doi.org/10.1155/2019/9106395
Wani ZA, Kumar A, Sultan P, Bindu K, Riyaz-Ul-Hassan S, Ashraf N (2017) Mortierella alpina CS10E4, an oleaginous fungal endophyte of Crocus sativus L. enhances apocarotenoid biosynthesis and stress tolerance in the host plant. Sci Rep 7:1–11. https://doi.org/10.1038/s41598-017-08974-z
Arora P, Wani ZA, Ahmad T, Sultan P, Gupta S, Riyaz-Ul-Hassan S (2019) Community structure, spatial distribution, diversity and functional characterization of culturable endophytic fungi associated with Glycyrrhiza glabra L. Fungal Biol 123:373–383. https://doi.org/10.1016/j.funbio.2019.02.003
Turner S, Pryer KM, Miao VP, Palmer JD (1999) Investigating deep phylogenetic relationships among cyanobacteria and plastids by small subunit rRNA sequence analysis 1. J Eukaryot Microbiol 46:327–338. https://doi.org/10.1111/j.1550-7408.1999.tb04612.x
Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ (1990) Basic local alignment search tool. J Mol Biol 215:403–410. https://doi.org/10.1016/S0022-2836(05)80360-2
Tamura K, Stecher G, Peterson D, Filipski A, Kumar S (2013) MEGA6: molecular evolutionary genetics analysis version 6.0. Mol Biol Evol 30:2725–2729. https://doi.org/10.1093/molbev/mst197
Qadri M, Rajput R, Abdin MZ, Vishwakarma RA, Riyaz-Ul-Hassan S (2014) Diversity, molecular phylogeny, and bioactive potential of fungal endophytes associated with the Himalayan blue pine (Pinus wallichiana). Microb Ecol 67:877–887. https://doi.org/10.1007/s00248-014-0379-4
Hammer O, Harper DA, Ryan PD (2001) Palaeontological statistics software package for education and data analysis. Palaeontol Electron 4:9
Kumar G, Kanaujia N, Bafana A (2012) Functional and phylogenetic diversity of root-associated bacteria of Ajuga bracteosa in Kangra valley. Microbiol Res 167:220–225. https://doi.org/10.1016/j.micres.2011.09.001
Sierra G (1957) A simple method for the detection of lipolytic activity of micro-organisms and some observations on the influence of the contact between cells and fatty substrates. Antonie Van Leeuwenhoek 23:15–22. https://doi.org/10.1007/BF02545855
Gordon SA, Weber RP (1951) Colorimetric estimation of indoleacetic acid. Plant Physiol 26:192. https://doi.org/10.1104/pp.26.1.192
Penrose DM, Glick BR (2003) Methods for isolating and characterizing ACC deaminase-containing plant growth-promoting rhizobacteria. Physiol Plant 118:10–15. https://doi.org/10.1034/j.1399-3054.2003.00086.x
Jensen HL (1942) Nitrogen fixation in leguminous plants. II. Is symbiotic nitrogen fixation influenced by Azotobacter. Proc Linn Soc NSW 67:205–212
Cappuccino JG, Sherman N (1996) Instructor’s guide for microbiology: a laboratory manual, 10th Edition, Pearson, UK
Pikovskaya RI (1948) Mobilization of phosphorus in soil in connection with vital activity of some microbial species. Mikrobiologiya 17:362–370
Andrews MY, Santelli CM, Duckworth OW (2016) Digital image quantification of siderophores on agar plates. Data in Brief 6:890–898. https://doi.org/10.1016/j.dib.2016.01.054
Murthy N, Bleakley B (2012) Simplified method of preparing colloidal chitin used for screening of chitinase-producing microorganisms. Internet J Microbiol 10:2
Lorck H (1948) Production of hydrocyanic acid by bacteria. Physiol Plant 1:142–146
Singleton VL, Orthofer R, Lamuela-Raventos RM (1999) Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-Ciocalteau reagent. Method Enzymol 299:152–178. https://doi.org/10.1016/S0076-6879(99)99017-1
Akkol EK, Goger F, Kosar M, Başer KH (2008) Phenolic composition and biological activities of Salvia halophile and Salvia virgate from Turkey. Food Chem 108(3):942–949. https://doi.org/10.1016/j.foodchem.2007.11.071
Porra RJ, Thompson WA, Kriedemann PE (1989) Determination of accurate extinction coefficients and simultaneous equations for assaying chlorophylls a and b extracted with four different solvents: verification of the concentration of chlorophyll standards by atomic absorption spectroscopy. Biochim Biophys Acta 975:384–394. https://doi.org/10.1016/S0005-2728(89)80347-0
Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 25:402–408. https://doi.org/10.1006/meth.2001.1262
Compant S, Clément C, Sessitsch A (2010) Plant growth-promoting bacteria in the rhizo-and endosphere of plants: their role, colonization, mechanisms involved and prospects for utilization. Soil Biol Biochem 42:669–678. https://doi.org/10.1016/j.soilbio.2009.11.024
Gaiero JR, McCall CA, Thompson KA, Day NJ, Best AS, Dunfield KE (2013) Inside the root microbiome: bacterial root endophytes and plant growth promotion. Am J Bot 100:1738–1750. https://doi.org/10.3732/ajb.1200572
Chimwamurombe PM, Grönemeyer JL, Reinhold-Hurek B (2016) Isolation and characterization of culturable seed-associated bacterial endophytes from gnotobiotically grown Marama bean seedlings. FEMS Microbiol Ecol 92, fiw083. https://doi.org/10.1093/femsec/fiw083
Sharma T, Kaul S, Dhar MK (2015) Diversity of culturable bacterial endophytes of saffron in Kashmir. India SpringerPlus 4:661. https://doi.org/10.1186/s40064-015-1435-3
Shade A, Handelsman J (2012) Beyond the Venn diagram: the hunt for a core microbiome. Environ Microbiol 14:4–12. https://doi.org/10.1111/j.1462-2920.2011.02585.x
Brader G, Compant S, Vescio K, Mitter B, Trognitz F, Ma LJ, Sessitsch A (2017) Ecology and genomic insights into plant-pathogenic and plant-nonpathogenic endophytes. Annu Rev Phytopathol 55:61–83. https://doi.org/10.1146/annurev-phyto-080516-035641
Glick BR (2012) Plant growth-promoting bacteria: mechanisms and applications. Scientifica 2012. https://doi.org/10.6064/2012/963401
Hilbert M, Voll LM, Ding Y, Hofmann J, Sharma M, Zuccaro A (2012) Indole derivative production by the root endophyte Piriformospora indica is not required for growth promotion but for biotrophic colonization of barley roots. New Phytol 196:520–534. https://doi.org/10.1111/j.1469-8137.2012.04275.x
Zúñiga A, Poupin MJ, Donoso R, Ledger T, Guiliani N, Gutiérrez RA, González B (2013) Quorum sensing and indole-3-acetic acid degradation play a role in colonization and plant growth promotion of Arabidopsis thaliana by Burkholderia phytofirmans PsJN Mol Plant Microbe Interact 26:546–553. https://doi.org/10.1094/MPMI-10-12-0241-R
Glick BR (2014) Bacteria with ACC deaminase can promote plant growth and help to feed the world. Microbiol Res 169:30–39. https://doi.org/10.1016/j.micres.2013.09.009
Dariush S, Emtiazi G (2010) Ammonium production during the nitrogen-fixing process by wild Paenibacillus strains and cell-free extract adsorbed on nano TiO2 particles. J Microbiol Biotechn 20:1251–1258. https://doi.org/10.4014/jmb.1003.03002
Castagno LN, Estrella MJ, Sannazzaro AI, Grassano AE, Ruiz OA (2011) Phosphate-solubilization mechanism and in vitro plant growth promotion activity mediated by Pantoea eucalypti isolated from Lotus tenuis rhizosphere in the Salado River Basin (Argentina). J Appl Microbiol 110:1151–1165. https://doi.org/10.1111/j.1365-2672.2011.04968.x
Loaces I, Ferrando L, Scavino AF (2011) Dynamics, diversity and function of endophytic siderophore-producing bacteria in rice. Microb Ecol 61:606–618. https://doi.org/10.1007/s00248-010-9780-9
Zhang T, Huang C, Deng C, Zhang Y, Feng Y, Hu J, Wang R, Zhao L, Wang Y, Kai G (2020) First report of corm rot on saffron caused by Penicillium solitum in China. Plant Dis 104:579–579. https://doi.org/10.1094/PDIS-09-19-1927-PDN
Mousa WK, Shearer C, Limay-Rios V, Ettinger CL, Eisen JA, Raizada MN (2016) Root-hair endophyte stacking in finger millet creates a physicochemical barrier to trap the fungal pathogen Fusarium graminearum. Nat Microbiol 1:1–12. https://doi.org/10.1038/nmicrobiol.2016.167
Swain DM, Yadav SK, Tyagi I, Kumar R, Kumar R, Ghosh S, Das J, Jha G (2017) A prophage tail-like protein is deployed by Burkholderia bacteria to feed on fungi. Nat Commun 8:1–9. https://doi.org/10.1038/s41467-017-00529-0
Conrath U, Beckers GJ, Langenbach CJ, Jaskiewicz MR (2015) Priming for enhanced defense. Annu Rev Phytopathol 53:97–119. https://doi.org/10.1146/annurev-phyto-080614-120132
Shimosaka M, Fukumori Y, Narita T, Zhang XY, Kodaira R, Nogawa M, Okazaki M (2001) The bacterium Burkholderia gladioli strain CHB101 produces two different kinds of chitinases belonging to families 18 and 19 of the glycosyl hydrolases. J Biosci Bioeng 91:103–105. https://doi.org/10.1016/S1389-1723(01)80123-7
Singh BK, Trivedi P, Egidi E, Macdonald CA, Delgado-Baquerizo M (2020) Crop microbiome and sustainable agriculture. Nat Rev Microbiol 18:601–602. https://doi.org/10.1038/s41579-020-00446-y
Shahidi F, Ambigaipalan P (2015) Phenolics and polyphenolics in foods, beverages and spices: antioxidant activity and health effects—a review. J Funct Foods 18:820–897. https://doi.org/10.1016/j.jff.2015.06.018
Pandey SS, Singh S, Pandey H, Srivastava M, Ray T, Soni S, Pandey A, Shanker K, Babu CV, Banerjee S, Gupta MM (2018) Endophytes of Withania somnifera modulate in planta content and the site of withanolide biosynthesis. Sci Rep 8:1–19
Ray T, Pandey SS, Pandey A, Srivastava M, Shanker K, Kalra A (2019) Endophytic consortium with diverse gene-regulating capabilities of benzylisoquinoline alkaloids biosynthetic pathway can enhance endogenous morphine biosynthesis in Papaver somniferum. Front Microbiol 10:925. https://doi.org/10.3389/fmicb.2019.00925
Shehata HR, Raizada MN (2017) A Burkholderia endophyte of the ancient maize landrace Chapalote utilizes c-di-GMP-dependent and independent signaling to suppress diverse plant fungal pathogen targets. FEMS Microbiol Lett 364:138. https://doi.org/10.1093/femsle/fnx138
Fiori M, Ligios V, Schiaffino A (2011) Identification and characterization of Burkholderia isolates obtained from bacterial rot of saffron (Crocus sativus L.) grown in Italy. Phytopathol Mediterr 50:450–461. https://www.jstor.org/stable/26556466
Eberl L, Vandamme P (2016) Members of the genus Burkholderia: good and bad guys. F1000Research 5. https://doi.org/10.12688/f1000research.8221.1
Mannaa M, Park I, Seo YS (2019) Genomic features and insights into the taxonomy, virulence, and benevolence of plant-associated Burkholderia species. Int J Mol Sci 20:121. https://doi.org/10.3390/ijms20010121
Ahmad T, Bashir A, Farooq S, Riyaz-Ul-Hassan S (2021) Burkholderia gladioli E39CS3, an endophyte of Crocus sativus Linn., induces host resistance against corm-rot caused by Fusarium oxysporum. J Appl Microbiol. https://doi.org/10.1111/jam.15190
Acknowledgements
TA is grateful to the University Grant Commission, New Delhi, India, for the Senior Research Fellowship. SF is supported by the Department of Science and Technology, New Delhi, India, through INSPIRE Senior Research Fellow. The article was approved by the institutional IPR committee as Manuscript no. CSIR-IIIM/IPR/00270.
Funding
This work was supported by the Council of Scientific and Industrial Research (CSIR), New Delhi, India, through the Major Lab Projects (MLP1002 and MLP1008) of the institute.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Ethics Approval
Not applicable.
Conflict of Interest
The authors declare no competing interests.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Ahmad, T., Farooq, S., Mirza, D.N. et al. Insights into the Endophytic Bacterial Microbiome of Crocus sativus: Functional Characterization Leads to Potential Agents that Enhance the Plant Growth, Productivity, and Key Metabolite Content. Microb Ecol 83, 669–688 (2022). https://doi.org/10.1007/s00248-021-01810-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00248-021-01810-y