
Abstract
In the present study, the influence of the land use intensity on the diversity of ammonia oxidizing bacteria (AOB) and archaea (AOA) in soils from different grassland ecosystems has been investigated in spring and summer of the season (April and July). Diversity of AOA and AOB was studied by TRFLP fingerprinting of amoA amplicons. The diversity from AOB was low and dominated by a peak that could be assigned to Nitrosospira. The obtained profiles for AOB were very stable and neither influenced by the land use intensity nor by the time point of sampling. In contrast, the obtained patterns for AOA were more complex although one peak that could be assigned to Nitrosopumilus was dominating all profiles independent from the land use intensity and the sampling time point. Overall, the AOA profiles were much more dynamic than those of AOB and responded clearly to the land use intensity. An influence of the sampling time point was again not visible. Whereas AOB profiles were clearly linked to potential nitrification rates in soil, major TRFs from AOA were negatively correlated to DOC and ammonium availability and not related to potential nitrification rates.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Since 2004, when the first evidence was provided that also organisms belonging to the kingdom of archaea are involved in ammonium oxidation [1–3], there is an ongoing debate to what extent these organisms contribute to nitrification and which factors drive their abundance, diversity, and activity in the environment. Today, it is generally acknowledged that ammonia oxidizing archaea (AOA) are ubiquitous [4] and several recently published studies assume a niche separation of AOA and their bacterial counterpart (AOB), which cannot be explained by a single environmental factor like ammonia availability, temperature or soil type [5–7]. Several studies based on the so far obtained isolates have also indicated a potential for the ammonia oxidation of AOA [8–11]. However, the turnover rates for ammonia do not only differ among AOB but also for the so far isolated AOA different ammonia oxidation rates have been measured. For AOB, ammonia oxidation rates between 21 and 43 fmol NO2 − cell−1 h−1 have been described [12, 13]. For AOA, the ammonia oxidation rates, which have been reported, were lower and ranged from 1.2-15 fmol NO2 − cell−1 h−1 [3, 10]. Thus, there is a need to measure not only the abundance of AOA and AOB in a particular environment, as it was done in a large number of studies in the last years, but also to monitor the dynamics in between both groups of ammonia oxidizers to link the population structure to a given function.
In a recent study, we could show that the land use intensity (LUI) of grassland ecosystems correlates positively with the abundance of AOA and AOB [14] as well as with potential nitrification rates (PNR). Whereas the amount of dissolved organic carbon present in soil could be identified as major driver for the abundance of denitrifiers and nitrogen-fixing microbes, we could identify inorganic nitrogen (mainly nitrate) concentrations closely related to the abundance of AOA and AOB in the investigated soil samples. Besides LUI also the time point of sampling (spring vs summer) had a clear effect on the abundance pattern mainly of AOB, with significant higher abundance on intensively used plots in summer. Overall, the observations on the effects of the LUI were more pronounced during in the middle of the vegetation period than in spring shortly after plant growth started.
In the present study, we show data as to how LUI changes the diversity pattern of AOA and AOB over time in the same ecosystems. Therefore, we used fingerprinting techniques of the bacterial and archaeal amoA gene and linked the obtained diversity pattern to major abiotic soil parameters of the given sites. We hypothesize that both the AOA and AOB diversity patterns are mainly driven by the time point of sampling as a response to the plant growth state and LUI only induces additional comparable smaller shifts in the structure of AOA and AOB.
Material and Methods
Site Description and Sampling
The present study is part of the German “Biodiversity Exploratories” [15], which forms an open multidisciplinary research platform to investigate the relationship between functional biodiversity, land use and ecosystem services. For the present study, soil samples were taken in the southernmost Exploratory “Schwäbische Alb” which covers more than 45,000 ha of the state of Baden-Württemberg in SW-Germany. Nine different grassland sites (AEG 1-9) were selected in this area. Due to long-term differences in the land use intensity, the selected sites could be categorized in three different land use intensity categories: intensely used meadow (IM, three times manure application and two times mown per season; AEG 1-3), intensely used mown pasture (IP, grazed by cattle and horses, mown once a year and two times manure application per season; AEG4-6) and extensively used pasture (EP, unfertilized and infrequently grazed by sheep; AEG7-9). The mean annual precipitation in this area ranged from 938–963 mm, whereas the annual mean temperature was around 7 °C. All sampled soils have been described as a Rendzic Leptosols with a clayey or loamy texture and a pH value between 5.7 and 6.9.
The number of plant species on the investigated sites ranged between 17 and 58. Significantly lower numbers were observed on the intensively managed plots (24 respectively 25 for IM and IP) compared to the EP plots where 49 different species were found (p = 0.019). While grasses were dominant on the intensely used meadows (58 to 100 % of coverage), the pastures (EP and IP) were more colonized by herbs (84 to 100 % of coverage).
From each site, five replicates were analyzed; each replicate consisted of five pooled bulk soil cores (d = 5.5 cm) from a soil depth of 0–10 cm. All samples were frozen directly after sampling for DNA based analyses at −20 °C as well as kept cool for enzyme activity and soil parameter measurements at 4 °C. Samples were taken in April and July 2008, representing the beginning of the vegetation period and the time point of flowering, where highest exudation rates could be expected.
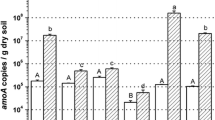
Major abiotic labile and stable soil parameters, the abundance of AOA and AOB as well as potential nitrification rates have been described elsewhere [14] and have been summarized in Table 1.
Nucleic Acid Extraction and Fingerprinting of amoA Amplicons using TRFLP
Genomic DNA was extracted from 0.5 g bulk soil (wet weight) using FastDNA Spin Kit for soil (MP Biomedicals, Germany) according to the manufacturer’s protocol. Quality and quantity of DNA extracts were determined with Nanodrop 1000 Spectrophotometer (Peqlab, Germany).
For the amplification of the bacterial amoA gene, the primer pair amoA 1 F (5′-GGGGTTTCTACTGGTGGT-3′) and amoA 2R (5′-CCCCTCKGSAAAGCC TTCTTC-3′) with an optimal annealing temperature of 60 °C were used generating a 491 bp fragment; for the archaeal amoA gene the primer pair 19F (5′-ATGGTCTGGCTWAGACG-3′) and CrenamoA616r48x (5′-GCCATCCABCKRTANGTCCA-3′) with an optimal annealing temperature of 58 °C were used amplifying a 624 bp fragment. Each forward primer was labeled at the 5′end with Carboxyfluorescein. Details on the PCR conditions can be found elsewhere [16].
The amoA amplicons were purified with Qiaquick PCR Purification Kit (Qiagen, Germany) and quantified with Nanodrop 1000 Spectrophotometer (Peqlab, Germany). The selection of digestion enzymes was done by the open source software Restriction Enzyme Picker Online, v.1.3. Three different enzymes for AOA (HhaI, Tsp509I and HpyCH4III) and two different enzymes for AOB (HhaI and TaqαI) were used; 200 ng of purified amoA amplicon were digested in a final volume of 25 μl for 2.5 h according to the instructions provided by the manufacturer. The enzymatic reaction was purified with Minelute Reaction Clean Up Kit (Qiagen, Germany). Approximately 5 ng of the digested amplicons were added to 13 μl of HiDi Formamide (Applied Biosystems, Germany) containing a 1:400 dilution of 6-carboxy-X rhodamine-labeled MapMarker 1000 ROX Size ladder (Bioventures, USA). After denaturation at 95 °C for 5 min, a capillary electrophoresis was carried out on an ABI 3730 DNA analyzer (Applied Biosystems) under the following conditions: 10 s injection time, 2 kV injection voltage, 7 kV run voltage, 66 °C
Statistical Analysis
TRFLP patterns of the amoA gene of both the AOA and AOB were dissected using Correspondence Analysis (CA), taking the respective peak area as a measure for the abundance of a particular OTU. Fragments smaller than 30 bases and TRFs contributing with <1 % of the total signal height were excluded from the statistical analysis. Detrended correspondence analysis was used to examine whether a linear or unimodal response of the data should be considered. The ordination was performed using a unimodal model to maximize the amount of explained variance in the TRFLP patterns with focus scaling on interspecies distances using Hill’s scaling and log-transformed species data. Leave-one-out cross-validation (LOOCV) rates were calculated to test if samples were classified in the right LUI based on the obtained TRFLP data. LOOCV involves using a single observation from the original sample as the validation data and the remaining observations as the training data. This is repeated in a way that each observation in the sample is used once as the validation data.
Results and Discussion
Diversity of AOA and AOB as affected by land use intensity and sampling time point
For AOB TRF 283B was the most dominant OTU present in each sample with a relative abundance up to almost 90 %. Based on in silico analysis, this peak could be identified as Nitrosospira (data not shown). TRF 281B was the second prevalent OTU and was mainly present in samples from IM and IP, whereas TRF 50 was typically for samples from EP plots. Surprisingly, overall, nearly no influence of the season was found and the TRFLP pattern between April and July did not differ significantly. TRF 220B was only detectable in July on soils characterized as EP. Due to the stable AOB pattern based on the LOOCV method, no clear assignment of the TRFLP profiles of AOB to the three land use intensity levels studied in this paper was possible (LOOCV rate 0.66667)
More than 50 % of the archaeal ammonia oxidizers were represented by TRF 374B and TRF 222B. The in silico analysis showed that TRF 374B was closely related to the fosmid clone 54d9, which belongs to the Crenarchaeal group I.1b [2] whereas TRF 222B could be assigned to the soil cluster of ammonia oxidizing archaea closely related to Nitrosopumilus and was found in almost all sampled plots at both time points. TRF 167B, TRF 368B and TRF 524B were only presented in IM and in EP but not in IP. Interestingly AEG3, which differed also in soil properties compared to the other IM plots, built up a different AOA community than the other IM plots. TRF 602B, TRF 522B, and TRF 220B exclusively occurred on this plot. TRF 502B and TRF 230B were typical for samples derived from IP and IM plots; TRF 66B occurred only in samples from IP plots, whereas TRF 38B exclusively appeared in samples from EP. As for AOB, the effect of the sampling time point was lower compared to the land use intensity. The overall leave-one-out cross-validation rate was 0.96667 and almost every sample could classify using the LOOCV method in the right land use category. Major TRFs and their relative abundance for AOA and AOB are given in Table S1.
The described results were used to calculate the Shannon diversity index as well as evenness. As expected from the TRFLP profiles, the diversity of bacterial amoA genes was much lower compared to the archaeal amoA diversity (Table 2). In April, the Shannon index for the AOB was calculated as 0.35 compared to 1.5 for the archaeal counterpart. Similar results were obtained for July (AOB 0.41 vs AOA 1.65), indicating again a low influence of the sampling time point. The analysis of the evenness data confirmed these observations with lower evenness values for AOB compared to AOA and a low influence of the sampling time point.
Linking AOA and AOB Diversity to Soil Properties
The obtained TRLP profiles were used to link the diversity of AOA and AOB to major soil abiotic properties and potential nitrification rates (Fig. 1). The ordination explains 65.8 % of the variation in the AOA diversity data and 58 % for AOB. TRF 281, which has been described as major peak of the AOB profiles, could be clearly linked to PNR. Overall, however, due to the low variability of the AOB profiles no other links between AOB derived OTUs and measured soil properties could be made. In contrast amoA fingerprints of AOA changed with LUI and even differences between plots belonging to the same LUI category were visible, but to a lower extent (exept AEG3). The major TRFs (TRF 374B and TRF 222B) were slightly negatively correlated to the levels of dissolved organic carbon (DOC) in soil as well as to the ammonium concentrations measured. Only TRFs, which were detected in soils from EP (like TRF 38) responded positively to increased DOC contents. Surprisingly, most of the major TRFs were not or even negatively correlated to potential nitrification rates.

Correspondence analysis (CA) of TRFLP patterns of the amoA genes. Ordination was performed to maximize the amount of explained variance in the TRFLP patterns with focus scaling on interspecies distances using Hill’s scaling and log-transformed species data. Environmental variables are displayed for interpretation only, but have not been used for ordination. a archaeal amoA genes: The ordination explains on all axes 65.8 % of the variance in the TRFLP data, from that are 46.1 % shown on axis 1 (horizontal) and 25.1 % on axis 2 (vertical). b Bacterial amoA genes. The ordination explains on all axes 58.0 % of the variance in the TRFLP data, from that are 43.5 % shown on axis 1 (horizontal) and 14.5 % on axis 2 (vertical)
Conclusions
In contrast to our initial hypothesis, the AOA and AOB diversity is not related to the time point of sampling in soils from temperate grassland ecosystems, as the observed TRFLP pattern derived from amoA amplicons was very stable. This indicates that ammonia oxidizers, as part of the autotrophic microbiota in soil are not directly linked to the plant performance. Obviously, they are also not outcompeted by the heterotrophic microbiota in response to higher nutrient contents in soil during the vegetation period, as only a slight negative correlation to DOC was visible for some of the major AOA OTUs. This situation may change if plants are able to secrete nitrification inhibitors like it is well known for a large number of cereals [17]. However, also for some grasses (like Brachiaria humidicola) nitrification inhibitors have been described [18], but they were not present at our sites of investigation. In contrast, the land use intensity strongly influenced the diversity pattern of AOA, whereas the obtained diversity pattern for AOB was stable and did not indicate an influence, although an increase in the total gene copy numbers for AOB has been observed in response to the land use intensity [14]. However, also for AOA, the major OTUs did not change over time in relative abundance. The major OTUs for AOB and AOA, which have been assigned to Nitrosomonas respectively the Crenarchaeal soil cluster group I.1b and Nitrosopumilus, are in good agreement with published data on the dominant ammonia oxidizers in soil [19]. This data may also explain partly the high sensitivity of AOB towards stressors due to the very low diversity and the relatively high resilience of AOA in response to changing environments [20] as a result of a relatively high diversity. Finally, our data nicely illustrates that AOA might be an important driver of the ammonia oxidation but the formed hydroxylamine is not necessarily further oxidized into nitrite [21] as there was no positive correlation between the major TRFs from the AOA profiles and the potential nitrification rates found.
References
Venter JC, Remington K, Heidelberg JF, Halpern AL, Rusch D, Eisen JA et al (2004) Environmental genome shotgun sequencing of the Sargasso Sea. Science 304:66–74. doi:10.1126/science.1093857
Treusch AH, Leininger S, Kletzin A, Schuster SC, Klenk HP, Schleper C (2005) Novel genes for nitrite reductase and Amo-related proteins indicate a role of uncultivated mesophilic crenarchaeota in nitrogen cycling. Environ Microbiol 7(12):1985–1995. doi:10.1111/j.1462.2920.2005.00906
Könneke M, Bernhard AE, de la Torre JR, Walker CB, Waterbury JB, Stahl DA (2005) Isolation of an autotrophic ammonia-oxidizing marine archaeon. Nature 437(7058):543–546. doi:10.1038/nature03911
Stahl DA, de la Torre JR (2012) Physiology and diversity of ammonia-oxidizing archaea. Annu Rev Microbiol 66:83–101. doi:10.1146/annurev-micro-092611-150128
Yao C, Lei HY, Yu Q, Li SP, Li HL, Chen K, Zhang XH (2013) Application of magnetic enhanced bio-effect on nitrification: a comparative study of magnetic and non-magnetic carriers. Water Sci Technol 67(6):1280–1287. doi:10.2166/wst.2013.697
Fernàndez-Guerra A, Casamayor EO (2012) Habitat-associated phylogenetic community patterns of microbial ammonia oxidizers. PloS ONE 7(10):e47330. doi:10.1371/journal.pone.0047330
Prosser JI, Bohannan BJ, Curtis TP, Ellis RJ, Firestone MK, Freckleton RP, Green JL et al (2007) The role of ecological theory in microbial ecology. Nat Rev Microbiol 5(5):384–392. doi:10.1038/nrmicro1643
Nicol GW, Schleper C (2006) Ammonia-oxidising Crenarchaeota: important players in the nitrogen cycle? Trends Microbiol 14(5):207–212. doi:10.1016/j.tim.2006.03.004
Hatzenpichler R, Lebedeva EV, Spieck E, Stoecker K, Richter A, Daims H, Wagner M (2008) A moderately thermophilic ammonia-oxidizing crenarchaeote from a hot spring. Proc Natl Acad Sci U S A 105(6):2134–2139. doi:10.1073/pnas.0708857105
de la Torre JR, Walker CB, Ingalls AE, Könneke M, Stahl DA (2008) Cultivation of a thermophilic ammonia oxidizing archaeon synthesizing crenarchaeol. Environ Microbiol 10(3):810–818. doi:10.1111/j.1462-2920.2007.01506
Tourna M, Freitag TE, Nicol GW, Prosser JI (2008) Growth, activity and temperature responses of ammonia-oxidizing archaea and bacteria in soil microcosms. Environ Microbiol 10(5):1357–1364. doi:10.1111/j.1462-2920.2007.01563
Ward BB (1987) Kinetic studies on ammonia and methane oxidation by Nitrosococcus oceanus. Arch Microbiol 147:126–133
Prosser JI (1989) Autotrophic nitrification in bacteria. Adv Microb Physiol 30:125–181
Meyer A, Focks A, Radl V, Keil D, Welzl G, Schöning I, Boch S, Marhan S, Kandeler E, Schloter M (2013) Different land use intensities in grassland ecosystems drive ecology of microbial communities involved in nitrogen turnover in soil PloS ONE (accepted)
Fischer M, Bossdorf O, Gockel S, Hänsel F, Hemp A, Hessenmöller D et al (2010) Implementing large-scale and long-term functional biodiversity research: The Biodiversity Exploratories. Basic Appl Ecol 11:473–485. doi:10.1016/j.baae.2010.07.009
Schauss K, Focks A, Leininger S, Kotzerke A, Heuer H, Thiele-Bruhn S, Sharma S et al (2009) Dynamics and functional relevance of ammonia-oxidizing archaea in two agricultural soils. Environ Microbiol 11(2):446–56. doi:10.1111/j.1462-2920.2008.01783
Subbarao GV, Rondon M, Ito O, Ishikawa T, Rao IM, Nakahara K, Lascano C (2007) Biological nitrification inhibitors (BNI)—is it a wide spread phenomenon? Plant Soil 295:5–18. doi:10.1007/s11104-006-9159-3
Subbarao GV, Wang H, Berry H, Ishikawa I, Nakahara K, Ito O (2006) A bioluminescence assay to detect nitrification inhibitors released from plant roots: a case study with Brachiaria humidicola. Plant Soil 288:101–112. doi:10.1007/s11104-006-9094-3
Prosser JI, Nicol GW (2012) Archaeal and bacterial ammonia-oxidisers in soil: the quest for niche specialisation and differentiation. Trends Microbiol 20(11):523–531. doi:10.1016/j.tim.2012.08.001
Ollivier J, Wanat N, Austruy A, Hitmi A, Joussein E, Welzl G, Munch JC, Schloter M (2012) Abundance and diversity of ammonia oxidizing prokaryotes in the root-rhizosphere complex of Miscanthus x giganteus grown in heavy metal contaminated soils. Microb Ecol 64:1038–1064. doi:10.1007/s00248-012-0078
Vajrala N, Martens-Habbena W, Sayavedra-Soto LA, Schauer A, Bottomley PJ, Stahl DA, Arp DJ (2012) Hydroxylamine as an intermediate in ammonia oxidation by globally abundant marine archaea. Proc Natl Acad Sci U S A 110(3):1006–1011. doi:10.1073/pnas.1214272110
Acknowledgments
We thank the managers of the three exploratories, Swen Renner, Sonja Gockel, Kerstin Wiesner, and Martin Gorke for their work in maintaining the plot and project infrastructure; Simone Pfeiffer and Christiane Fischer giving support through the central office, Michael Owonibi for managing the central data base, and Markus Fischer, Eduard Linsenmair, Dominik Hessenmöller, Jens Nieschulze, Daniel Prati, Ingo Schöning, François Buscot, Ernst-Detlef Schulze, Wolfgang W. Weisser and the late Elisabeth Kalko for their role in setting up the Biodiversity Exploratories project. The work has been (partly) funded by the DFG Priority Program 1374 “Infrastructure-Biodiversity-Exploratories” (SCHL446/7-1.). Field work permits were issued by the responsible state environmental offices of Baden-Württemberg, Thüringen, and Brandenburg (according to § 72 BbgNatSchG).
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
ESM 1
(DOC 137 kb)
Rights and permissions
About this article
Cite this article
Meyer, A., Focks, A., Radl, V. et al. Influence of Land Use Intensity on the Diversity of Ammonia Oxidizing Bacteria and Archaea in Soils from Grassland Ecosystems. Microb Ecol 67, 161–166 (2014). https://doi.org/10.1007/s00248-013-0310-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00248-013-0310-4