
Abstract
Marine fungi are known to originate from a wide variety of habitats within the marine environment. Marine sediment represents one environmental niche, with most fungi occurring in these sediments being facultative marine fungi with terrestrial origins. It has not been proven whether these fungi merely survive the harsh environmental conditions presented by the ocean sediment, as opposed to playing an active role in this ecological niche. During this study, marine sediment was collected from St. Helena Bay, on the west coast of the Western Cape, South Africa. Using dilution, enrichment, and repetitive culturing techniques, 59 fungal isolates were obtained from marine sediments and identified to at least genus level using morphological and molecular methods. Moreover, a series of tests were performed to characterize the physical and physicochemical attributes of the isolates. Results showed that the isolates not only survived but also had the potential to grow in the natural conditions present in this environment. Extracellular cellulase was produced by the filamentous fungal isolates indicating their probable role in detrital decay processes and therefore the carbon cycle on the ocean bed. Also, denitrification patterns were observed when isolates were grown in liquid media amended with NaNO2, NaNO3 , and (NH4)SO4, implicating that these fungi have the potential to play an active role in denitrification, co-denitrification, and ammonification phases of nitrogen cycles occurring in the marine sediments.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Marine fungi are frequently isolated from plant detritus and decomposing wood found in coastal waters [11, 12, 16, 20] but have also been found on sea shells [8, 19], in corals [8], sponges [30], coastal marine sediments [3], deep-sea sediments, deep-sea hydrothermal vents [2], and even anoxic marine sediments [20]. These fungi are the least studied of all marine organisms, forming a group defined by ecology and physiology, rather than taxonomic clusters [12]. Two distinct groups can be distinguished: facultative marine fungi which originate from the terrestrial or fresh water environment and adapt to marine conditions and obligate marine fungi that live and propagate exclusively in the marine aquatic habitat [16, 20]. Most species fall in the facultative group and belong to genera such as Penicillium, Aspergillus, Trichoderma, Wardomyces, Chrysosporium, and Chaetomium [15, 22, 24], as well as Cephalosporium, Cladosporium, and Alternaria [23, 24]. Less commonly reported species include those that represent the facultative marine genera Scytalidium, Verticillium, and Oidiodendron, as well as obligate marine fungi from the genus Dendriphiella [15].
Spores of facultative marine fungi enter pelagic sediment by being swept from land soils into the ocean or are carried on air currents from land areas, after which these spores gradually descend to the sea bed. Many fungi are known for their ability to withstand harsh environmental conditions and even to grow vegetatively under inhospitable circumstances, and the marine environment is no exception. However, it has not been proven whether these species merely survive and endure the prevailing conditions or play an active role in the ecosystem [16, 23, 29]. The marine environment is ultimately a unique habitat presenting marine fungi with a number of physical challenges: (1) salinity and pH of the water—marine fungi are known to grow and produce functional enzymes at pH 7–8; (2) low water potential, which is overcome by accumulating osmolytes such as glycerol, mannitol, polylol, and trehalose; (3) high sodium-ion concentration, which is tolerated by using a very efficient sodium efflux ATPase or storage in vacuoles; (4) low temperatures; and (5) high hydrostatic pressure. These unusual circumstances inevitably lead to organisms with novel genes and unique properties [13, 20].
Marine fungi from sediment had been reported to be involved in denitrification processes [3, 21]. A study by Cathrine and Raghukumar [3] showed that the majority of fungi isolated were common terrestrial fungi that adapted to marine conditions. Fungal denitrification takes place under suboxic conditions (300–900 μM O2), and excess oxygen (>900 μM O2) inhibits the process; in contrast, bacterial denitrification requires completely anoxic conditions. Fungal denitrifiers were presumed not to complete the denitrification process to dinitrogen (N2) but form nitrous oxide (N2O) instead, thereby contributing to an increase in harmful greenhouse gases. However, some authors found that many denitrifying fungi are capable of producing hybrid N2 or N2O molecules, by combining nitrogen (N) atoms from nitrite (NO −2 ) with other N-compounds such as azide or ammonium (NH +4 ), during denitrifying conditions. This process known as co-denitrification would be induced in the presence of NO −2 or nitrate (NO −3 ) [10, 21, 27]. Furthermore, in anoxic conditions, fungi produce NH +4 when reducing NO −3 through a process known as ammonia fermentations (ammonification). Fungi are therefore capable of performing a “multi-model type of respiration,” depending on the O2 levels in their environment [3, 9, 27].
St. Helena Bay is a shallow, semi-enclosed bay east of Cape Columbine (Western Cape, South Africa), representing one of the main upwelling centers in the Southern Benguela upwelling system. It is a major controlling factor of the biogeochemical characteristics of the bay, and sediments in the bay come from either atmospheric input or are carried there by the Berg River. Fine-grained offshore sediment from as far north as the Orange River’s mouth is also known to be transported to the bay in a subsurface southward current. The Berg River and its tributaries flow through areas dominated by agriculture, wineries, canneries, and textile milling. The upper reaches of the river flow mainly through the Table Mountain Group sandstone, granitic hills, and clay soils consisting of weathered granitic material. Downstream, sandstone had been eroded to expose Malmesbury Group shale that forms the underlying bed rock for the remaining part of the river channel to the mouth. Many aquatic ecosystems are supported in St. Helena Bay and the area is known to be rich in marine life [18]. The aims of our study were to determine the diversity of culturable fungal strains from marine sediment collected from St. Helena Bay on the west coast of Southern Africa and to typify these isolates using a number of physiological characteristics in an effort to understand their possible role in the ecology of this unique habitat.
Methods
Sampling
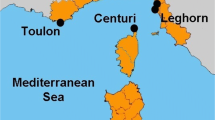
Sediment samples were collected on 14 February 2009 from five sampling stations in St. Helena Bay (and Great Berg River estuary), Western Cape, South Africa. The stations included St 11 (S32 40 21.7, E18 01 25.3, depth 28.0 m), St 24 (S32 35 59.3, E18 07 27.1, depth 32.0 m), St 227 (S32 30 31.6, E18 05 39.3, depth 15.0 m), St 43a (S32 29 32.3, E18 06 57.6, depth 61.0 m), and St 228 (S32 44 38.5, E18 00 45.9, depth 8.0 m) (Fig. 1). Sampling was done by coring (60 × 300-mm cores), using an Ocean Instruments MC-200 multi-corer operated from the CSIR semirigid vessel, “Wave Rider.” Due to strong wind and rough sea conditions, only one coring procedure could be performed at each sampling site. Subsequent to their collection, cores were stored at 4°C.

Map showing the location of the sediment core samples collected in 2009 from St. Helena Bay in the Great Berg River estuary along the Western Cape of South Africa
Isolation
In the laboratory, the cores were removed from the acrylic core barrels by sliding them into a sterile glass bowl. Sterilized spatulas were used to scoop ca. 1 cm3 sediment subsamples from the top 5 cm of each core; each of these subsamples was suspended in 100 ml sterile sea water. Different isolation methods were subsequently employed to ensure optimal culturing conditions and to accommodate possible low spore numbers. For each subsample, a dilution series (10−1 to 10−6) was prepared in triplicate, using sterile sea water. MFA medium (2 % D(+) Glucose (Saarchem, Merck Chemicals, South Africa), 1 % soluble starch, 2 % tryptone soy broth (Biolab, Merck, South Africa), 0.3 % meat extract powder (Biolab, Merck, South Africa), 0.5 % yeast extract powder (Biolab, Merck, South Africa), 1.5 % bacteriological agar (Biolab, Merck, South Africa), 0.2 g/l chloramphenicol (Sigma-Aldrich, Germany), and 1 l sea water) was prepared as a culture medium for the dilutions. Other isolation media included malt extract agar (MEA; Biolab, Merck, South Africa) and oatmeal agar (OA; 3 % oat flakes simmered (30 min) in distilled water, clarified by filtration through a stainless steel sieve, mixed with 1.5 % bacteriological agar (Biolab, Merck, South Africa)), supplemented with distilled water to 1 l and autoclaved (121°C for 15 min). In addition to this method, a second method involved the filtering of a spatula-scooped subsample suspended in 100 ml sterile sea water, through a sterile Sartorius Filter system, fitted with a cellulose acetate filter (0.2 μm; Sartorius Stedim Biotech, Germany); filters were subsequently placed in Petri dishes containing MFA, MEA, and OA. A third isolation method was also employed, wherein 1 ml of the sediment suspensions of all subsamples was enriched in 100 ml MF broth (MFA without agarose) in an Erlenmeyer flask incubated on a shaker at 28°C for 24 h in the dark. Isolations of the enriched microorganisms were done by transferring 100 μl liquid culture from each Erlenmeyer flask to a culture plate using a pipette (done in triplicate). A sterilized glass rod was used to spread the suspension across the entire surface of each culture plate. All plates were incubated at 25°C in the dark. The plates were examined daily for developing fungal colonies which appeared after 5 to 7 days. Colonies were counted on day 7 and transferred to fresh plates (repetitive culturing). Stock cultures of all fungal isolates were kept on MEA at 8°C, and all subsequent tests were performed using these stock cultures.
Identification
Fungal isolates were examined and identified to genus level using light microscopy. Classical morphological features of the fungal isolates were used for these preliminary identifications of the fungal isolates [6]. Genomic DNA was then extracted using the ZR Fungal/Bacterial DNA Kit™ (Zymo Research Corp., USA) according to the manufacturer’s instructions. The internal transcribed spacer regions (ITS1 and ITS2) of the ribosomal DNA operon (including the 5.8S gene) of the filamentous fungi were subsequently amplified using the primer set ITS1 (5′-TCCGTAGGTGAACCTGCGG-3′) and ITS4 (5′-TCCTCCGCTTATTGATATGC-3′) [32]. For the yeast spp., the D1/D2 (600–650 bp) region of the large subunit (26S) rDNA was amplified by the polymerase chain reaction (PCR) employing the universal forward primer F63 (5′-GCATATCAATAAGCGGAGGAAAAG-3′) and reverse primer LR3 (5′-GGTCCGTGTTTCAAGACGG-3′). The PCR used for the D1/D2 region was performed in a 50-μl reaction mixture, containing 10 ng DNA, 2.5 mM MgCl2, 0.25 mM deoxynucleotide triphosphates, 0.1 μM of each primer, and 0.5 U Fermentas Taq. Amplification was in a GeneAmp PCR system (Applied Biosystems Inc, USA) with the following cycling parameters for 30 cycles: initial denaturation for 3 min at 94°C, denaturation for 1 min at 94°C, followed by annealing for 1 min at 61°C, elongation for 2 min at 72°C, followed by a final elongation of 7 min at 72°C. The amplified product was separated on an agarose gel (0.8 %, w/v) and visualized by ethidium bromide staining. The cycling parameters for the PCR employed to amplify the ITS-regions of the filamentous fungi [32] were the same as the PCR for the D1/D2 region, except for the annealing temperature which was set at 56°C. PCR products were purified by means of a High Pure PCR Product Purification Kit (Roche Molecular Biochemicals (Pty) Ltd., S.A.) and sequenced using a Hitachi 3730xl DNA Analyzer (Applied Biosystems, USA). The fungal sequences were compared to reported sequences using a BLAST search on the NCBI database (http://www.ncbi.nlm.nih.gov/blast/), as well as the database of the Centraalbureau for Schimmelcultures (http://www.cbs.knaw.nl). The results obtained for each isolate were compared to morphological data to determine the most likely identity. In addition, only sequence matches above 98.5 % were regarded as meaningful for identification to species level. In cases where sequence matches were above 99 % for two or more species in a genus that were different in morphology and colony characteristics, only the genus name with an allocated strain number were used in subsequent tests.
Physiological Tests
Physiological growth tests were conducted on filamentous fungal isolates by studying radial growth (measured on an x- and y-axis, across the bottom surface of each plate) on differential or growth media in Petri dishes for each selected environmental parameter. For all these tests, growth of the fungal colonies was measured by taking the average of two colony diameter measurements for each plate. Three Petri dishes were used at each test criterion (e.g., temperature) and the experiments were conducted in the dark and repeated once. The average colony diameters from the two experiments were then calculated. For the yeast species, similar physiological tests were performed but in liquid media and growth was measured spectrophotometrically at 600 nm, using a SmartSpec™Plus spectrophotometer (Biorad Laboratories Ltd, South Africa).
Physical Physiology
Isolates were cultured on MFA medium from stock cultures, and disks of fungal growth, 5 mm in diameter, were cut from culture plates with the back end of a sterilized Pasteur pipette and used to inoculate plates. Yeast strains were grown in test tubes containing 10 ml YPD broth (2 % D (+) glucose (Saarchem, Merck Chemicals, South Africa), 2 % peptone (Biolab, Merck, South Africa), and 1 % yeast extract (Biolab, Merck, South Africa) in distilled H2O). The effect of temperature was determined at 4, 15, 26, 37, and 45°C. Subsequent experiments were incubated at the respective optimal growth temperatures determined in this experiment. Growth at different concentrations of salinity was determined by modifying concentrations of sea salts (Instant ocean synthetic sea salt, Aquarium systems, France) in the MFA medium to 0, 1.5, 3.0, 4.5, and 6.0 %. The effect of pH was determined by altering the pH of MFA medium to 5, 6, 7, 8, 9, and 10 respectively, with 10 M NaOH or concentrated HCl. The ability of isolates to grow under anaerobic conditions was studied by incubating, inoculated MFA plates and test tubes containing YPD broth, in anaerobic chambers containing an Anaerocult A (Merck, Germany) set and Anaerotest (Merck, Germany) strip.
Physiochemistry
Three replicates for each isolate were used for all experiments described in this section. Ability to grow or secrete a specific enzyme or substance was documented as positive (+), while inability to grow or absence of the latter as negative (−). Protease activity of fungal isolates was determined by growing them on a solid medium in Petri dishes containing skimmed milk and a pH indicator (0.1 % yeast extract, 1 % skim milk, 2 % bacteriological agar, and 0.01 % bromocresol purple indicator (pH 5.2, Riedel De Haën AG Zeelze, Germany)). Subsequent to the incubation (24 h), plates were examined for the formation of clearing zones. Purple zones were assumed to indicate the secretion of ammonia, which raises the ambient pH and allows extracellular protease production and activity [25]. Cellulase production by the isolates was tested on carboxymethylcellulose medium (0.67 % yeast nitrogen base (Becton, Dickinson and Company Sparks, USA), 0.5 % carboxymethylcellulose (Sigma-Aldrich, Germany), and 2 % bacteriological agar) plates. After 7 days, the plates were stained using 0.1 % Congo red (B & M Scientific, South Africa) for 10 min, followed by de-staining with 1 M NaCl (KIMIX, South Africa) for 5 min. Clearing zones surrounding the fungal colony indicated cellulase activity. Hemolytic activity was determined by inoculating plates of Columbia agar base containing 5 % whole sheep blood. Plates were incubated at 26°C for 7 days and small yellow zones around the colonies indicated hemolysis.
Anaerobic Nitrogen Utilization
To determine nitrogen utilization under anaerobic conditions, Czapek Dox agar (CDA) medium was modified to contain the following in 1 l distilled water: 5.2 g KH2PO4, 6.4 g K2HPO4, 1.0 g MgSO4·7H2O, 1.0 g KCl, 0.02 g FeSO4, 0.02 g ZnSO4·7H2O, 0.008 g CuSO4·5H2O, 16.7 g glucose, 1.32 g (NH4)2SO4, 0.85 g NaNO3, and 0.28 g NaNO2. Test tubes, each containing 10 ml of the medium, were inoculated with a small amount of spores or hyphae and transferred with an inoculation loop from fungal colonies grown on the CDA medium. Negative controls, without any fungal growth, were included, along with positive controls inoculated with Botrytis cinerea spores. Regarding the positive controls, Shoun and coworkers [21] showed that B. cinerea is capable of evolving N2O, nitric oxide (NO), and/or N2 when incubated anaerobically with NO −2 as N source. The test tubes were incubated in anaerobic jars, as described above, for 3 weeks at 26°C. The NH4 and NO −3 concentrations were determined colorimetrically with the method described by Kartal et al. [14], and the NO −2 concentration was also verified colorimetrically with the method described by Egli et al. [7] using a BioRad SmartSpec Plus spectrophotometer. Each isolate was prepared in triplicate, and the average absorbance of the isolate was used to determine the ratio of absorbance to the average absorbance of the negative controls. The experiment was performed three times and the average ratio values of these experiments were calculated.
Results
Identities of Isolates
Only 10−1 and 10−2 dilutions of the diluted marine sediment yielded colonies. The 59 isolates included 25 different species of Acremonium, Aspergillus, Penicillium, Trichoderma, Paraconiothyrium, Eurotium, Rhodotorula, and Yarrowia (Tables 1 and 2). Using morphological as well as ITS data, many isolates could be identified to species level, and only sequence matches above 98.5 % were regarded as meaningful. Species of Aspergillus, Acremonium, Penicillium, and Trichoderma were also present, although in some cases, the methods and characteristics used were not specific enough to distinguish closely related species. In these cases, the sequence matching analysis was above 99 % for a number of different species within the genus. Sampling station 43a yielded 11 species, St 227 yielded 9 species, St 228 yielded 4 species, St 24 yielded only 1 species, and St 11, no isolates (Table 1 and Fig. 2).

Graph showing percentage mud, organic carbon and nitrogen, as well as carbon–nitrogen ratio and the number of species isolated from available sampling stations (St 11 = sampling station 11, St 24 = sampling station 24, St 43a = sampling station 43a, St 227 = sampling station 227, St 228 = sampling station 228)
Fungal Physiology
Physical Physiology
The optimum temperature for most of the fungi was 26°C. Seven isolates (numbers 7, 12-14, 15, 16, and 17) had an optimum temperature of 37°C (Table 1). All isolates were also able to grow at 15°C and some as low as 4°C. Most isolates preferred a 1.5–3 % salt concentration, although a number of isolates were more halotolerant, showing enhanced growth at 6 % salt, and only one isolate preferred 0 % salt (Table 1). The pH range for growth of the isolates was between pH 5 and 10 (Table 1). Most isolates were able to grow anaerobically, with only numbers 15, 16, 25, 26, and the yeast spp. as exceptions (Table 1).
Physiochemistry
All of the mycelial fungi were able to produce cellulase under aerobic conditions, although the two yeast species were unable to utilize cellulose as carbon source (Table 1). Aerobic ammonia production varied among isolates (Table 1). Hemolytic activity was observed in 13 of the species, as opposed to the 12 displaying no hemolytic ability (Table 1).
Anaerobic Nitrogen Utilization
Of the 33 isolates tested, 15 showed a nitrogen utilization pattern with a simultaneous decrease in NO −2 and a slight decrease in NH +4 , with elevated NO −3 concentrations. A different denitrification pattern was observed in the other isolates, where NO −3 was utilized and NO −2 produced or both NO −2 and NO −3 utilized (Table 5). The positive control showed a significant decrease in NO −2 and an increase in NH +4 , consistent with literature [21].
Discussion
Most previous studies on marine fungi focused mainly on fungal diversity [2, 20, 22, 23] and the presence of these fungi cannot testify to their participation in physiological processes, ecological role, or adaptations. This study was a pioneering effort to explore the physiology and possible ecological role of marine fungi in biogeochemical processes along the west coast of South Africa. These biogeochemical processes along the west coast, including St. Helena Bay, are primarily regulated by a wind-driven ocean upwelling occurring in the region [18]. This upwelling of cold nutrient-rich water drives the biological pump and hydrodynamics along the coast, determining the fate of sediment, organic matter, and trace elements, ultimately affecting the ecosystem of the region. During previous studies in this region, the physicochemical composition of sediment samples taken from the seabed at different sampling stations was used to construct plots of the sediment composition of St. Helena Bay [4, 5]. The sediments at the sampling stations were found to differ regarding percentage mud and the C/N ratio [5]. Sampling station 43a was characterized by a high percentage of mud (80 %) with a low C/N ratio (4.0), as opposed to samples from St 24, which had a much lower mud percentage (47 %) with a higher C/N ratio (10.0) (Tables 3 and 4). Samples from St 11 had a mud percentage of 55 % with a C/N ratio of 7.93 and were therefore more similar to samples from St 24 than to samples from St 43a, in percentage mud and C/N ratio. Stations 227 and 228 were not sampled during the 1999 [4] and 2007 [5] surveys. However, according to the constructed plots portraying sediment composition in the bay [4, 5], the physicochemical composition of the sediment at St 227 is similar to that of St 43a.
Our study showed a greater fungal diversity at St 43a (11 species) versus stations 24 (one species) and 11 (zero isolates); we attribute this phenomenon to differences in the physicochemical composition of the sediment at the different sampling stations (Tables 3 and 4). The west coast of Southern Africa is characterized by water temperatures between 10 and 14°C, with even lower temperatures (2–3°C) in the deep-sea [18]. Water salinity along the coast would be 3.5 % and pH between 7.5 and 8.4. Our findings showed that a number of fungal isolates were able to grow at low temperatures (as low as 4°C), in the pH range of sea water and even at salinity levels higher than that of the bay (Table 1). Interestingly, most isolates were able to grow anaerobically, suggesting that they have adapted to anaerobic conditions presented by the oxygen-deficient environment within the sediment. These results indicated that the majority of fungal isolates not only survive but have the potential to grow at conditions presented by the ocean on the west coast of South Africa.
Sampling station 43a yielded the highest number of isolates, although this station represented the deepest sampling station in this study. We hypothesize that the high mud fraction, high levels of organic matter, and low C/N ratio of St 43a may provide an environment supporting growth of a larger number of microbes. It must be noted, however, that the potential nutrients in the sediment might not necessarily be bioavailable but fixed in microbial biomass. Moreover, all of the taxa isolated from St 43a (Acremonium, Aspergillus, Penicillium, and Eurotium) are well-known terrestrial fungi, supporting contentions that spores of these fungi had been transported from land areas into the deep sea [22, 29]. The sampling station closest to St 43a, i.e., St 227 also yielded a variety of other known terrestrial fungi (Aspergillus and Penicillium spp.), as well as the basidiomycetous yeast, Rhodotorula mucilaginosa (Tables 1 and 2). The latter species has a worldwide distribution in terrestrial, aquatic, and marine environments and has even been isolated from the Mariana Trench (11,500 m deep) in the Pacific Ocean [26]. Sampling station 228, situated in the shallow water of the harbor area (Fig. 1) yielded only four species, although it is of value to note that at least one of these species is known to be associated with higher anthropogenic activity. Yarrowia lipolytica had been shown to occur in hydrocarbons, kerosene, aviation fuel, contaminated gas or oil fuel, n-paraffin [28], and even human cornea, skin, and finger nails [1]. This species is well known for its unique physiological properties of industrial value [17, 20], including the utilization of n-paraffin, and therefore, most likely plays a role in the degradation of crude oil on the harbor bed [20]. Interestingly, it has previously been found that areas with intense anthropogenic loads were dominated by hemolytically active fungal strains. Genera containing the latter included Chaetomium, Penicillium, Aspergillus, and Trichoderma [15]. Our study also revealed the presence of Trichoderma and hemolytically active Aspergillus candidus isolates in sediment from the harbor area (Tables 1 and 2). However, hemolytically active fungi were not exclusively found in the harbor sediment but also in sediment from all our sampling stations. The ecological role of fungi with this characteristic is largely unknown, although it is tempting to speculate that it may play a role in the decay of dead animals; a role of which relatively little is known [11].
It is widely accepted that marine fungi are able to degrade plant material [11]. It was therefore not surprising to find that with the exception of the two yeast species, all the fungal isolates were able to produce extracellular cellulase, indicating their potential involvement in detrital processes and the crucial reminerilization processes occurring in detritus formation, as well as concomitant changes in the C/N ratio [31]. Aside from their role in the microbial regulation of the C/N ratio, marine fungi have also been documented to aid in the release of secondary metabolites from dead plant material. These metabolites include compounds such as omega-3 fatty acids, which are essential for the growth and development of crustaceans [20].
In this study, the highest number of isolates originated from the sampling site with the highest percentage of organic nitrogen. Consequently, the question arose as to whether these fungi are able to participate in the nitrogen cycle within the sediment. We therefore designed an experiment in which liquid cultures of the fungal isolates were challenged with inorganic nitrogen under anaerobic conditions. We found that in these liquid cultures amended with NaNO2, NaNO3, and (NH4)SO4, approximately 50 % of isolates showed nitrogen utilization patterns characterized by a decrease in NO −2 levels, a slight but simultaneous decrease in NH +4 levels, as well as elevated NO −3 concentrations (Table 5). These results suggest the occurrence of co-denitrification [21]. Another denitrification pattern was observed in cultures of most other isolates utilizing NO −3 and producing NO −2 or utilizing both NO −2 and NO −3 ; this pattern is known to occur in fungi [10]. These data therefore suggest that these fungal species indeed have the potential to be involved in different parts of the very complex nitrogen cycle, even in this challenging environmental niche, and confirm the observations of Shoun et al. [21] concerning denitrifying abilities that occur in a variety of fungal taxa. The positive control included in the experiment (Table 5) showed a significant decrease in NO −2 and an increase in NH +4 , consistent with literature [21], suggesting the occurrence of ammonification. This study therefore supports the sentiment of Raghukumar [20] that fungi may potentially play a role in the nitrogen cycle of the marine environment where hypoxic and anoxic conditions prevail.
This study showed that culturable fungi, originating from sea sediment in St. Helena Bay, have the potential to survive and grow in the physicochemical conditions presented by this marine environment and have the potential to contribute to carbon and nitrogen cycles. We therefore believe that this study contributes to knowledge of this unique environmental niche, since this is the first study on fungi and their role in the sediments of the nutrient-rich Southern Benguela system.
References
Barnett JA, Payne RW, Yarrow D (2000) Yeasts: Characteristics and identification, 3rd edn. Cambridge University Press, Cambridge
Burgaud G, Le Calvez T, Arzur D, Vandenkoornhuyse P, Barbier G (2009) Diversity of culturable filamentous fungi from deep-sea hydrothermal vents. Environ Microbiol 11:1588–1600
Cathrine SJ, Raghukumar C (2009) Anaerobic denitrification in fungi from the coastal marine sediment off Goa, India. Mycol Res 113:100–109
CSIR Report ENV-S-C 2002-050/A (2002) St. Helena Bay Quality Water Trust: the biochemical status of surface sediments in St. Helena Bay in 2001
CSIR Report ENV-S-C (2007) St. Helena Bay Quality Water Trust: the biochemical status of surface sediments in St. Helena Bay
De Hoog GS, Guarro J, Gené J, Figueras MJ (2000) Atlas of clinical fungi, 2nd edn. Centraalbureau for Scimmelcultures, Utrecht
Egli K, Fanger U, Alvarez PJJ, Siegrist H, Van der Meer JH, Zehnder AJB (2001) Enrichment and characterization of an anammox bacterium from a rotating biological contactor treating ammonium-rich leachate. Arch Microbiol 175:198–207
Golubic S, Radtke G, Le Campion-Alsumard T (2005) Endolithic fungi in marine ecosystems. Trends Microbiol 13:229–235
Greben HA, Joubert L-M, Tjatji MP, Whites HE, Botha A (2007) Biological nitrate removal from synthetic wastewater using a fungal consortium in one stage bioreactors. Water SA 33:285–290
Hayatsu M, Tago K, Saito M (2008) Various players in the nitrogen cycle: diversity and functions of the microorganisms involved in nitrification and denitrification. Soil Sci Plant Nutr 54:33–45
Hyde HD, Jones EBG, Leaño E, Pointing SB, Poonyth AD, Vrijmoed LLP (1998) Role of fungi in marine ecosystems. Biodivers Conserv 7:1147–1161
Hyde KD, Sarma VV, Jones EBG (2000) Morphology and taxonomy of higher marine fungi. In: Hyde KD, Pointing SM (eds) Marine mycology—a practical approach. Fungal Diversity Research, Hong Kong, pp 172–204
Jones EBG (2000) Marine fungi: some factors influencing biodiversity. Fungal Divers 4:53–73
Kartal B, Koleva M, Arsov R, Van der Star W, Jetten MSM, Strous M (2006) Adaptation of a freshwater anammox population to high salinity wastewater. J Biotechnol 126:546–553
Khudyakova YV, Pivkin MV, Kuznetsova TA, Svetashev VI (2000) Fungi in sediments of the sea of Japan and their biologically active metabolites. Microbiology 69:608–611
Kohlmeyer J, Kohlmeyer E (1979) Marine mycology. The higher fungi. Academic, New York
Kurtzman CP, Fell JW (2000) The yeasts. A taxonomic study, 4th edn. Elsevier Science BV, Amsterdam
Monteiro PMS, Roychoudhury AN (2005) Spatial characteristics of sediment trace elements in an eastern boundary upwelling retention area (St. Helena Bay, South Africa): a hydrodynamic–biological pump hypothesis. Estuar Coast Shelf S 65:123–134
Raghukumar C (1992) Endolithic fungi from deep sea calcareous substrata: isolation and laboratory studies. In: Desai BN (ed) Oceanography of the Indian Ocean. Oxford and IBH, New Delhi, pp 3–9
Raghukumar C (2008) Marine fungal biotechnology: an ecological perspective. Reviews Crit New Technol Fungal Divers 31:19–35
Shoun H, Kim D-H, Uchiyama H, Sugiyama J (1992) Denitrification by fungi. FEMS Microbiol Lett 94:277–282
Singh P, Raghukumar C, Verma P, Shouche Y (2011) Fungal community analysis in the deep-sea sediments of the central Indian basin by culture-independent approach. Microb Ecol 61:507–517
Sparrow FK Jr (1937) The occurrence of saprophytic fungi in marine muds. Biology Bull 73:242–248
Steele CW (1967) Fungus populations in marine waters and coastal sands of the Hawaiian, Line, and Phoenix Islands. Pac Sci 21:317–331
St Leger RJ, Nelson JO, Screen SE (1999) The entomopathogenic fungus Metarhizium anisopliae alters ambient pH, allowing extracellular protease production and activity. Microbiology 145:2691–2699
Takami H (1999) Isolation and characterization of microorganisms from deep-sea mud. In: Horikoshi K, Tsujii K (eds) Extremophiles in deep-sea environments. Springer, Tokyo, pp 3–26
Takaya N (2002) Review. Dissimilatory nitrate reduction metabolisms and their control in fungi. J Bio Sci Bioeng 94:506–510
Tatsunori F, Akira I, Kazuhito M, Koki H (1994) Isolation of a marine yeast that degrades hydrocarbon in the presence of organic solvent. Bio Sci Biotech Bioch 58:1784–1788
Vrijmoed LLP (2000) Isolation and culture of higher filamentous fungi. In: Hyde KD, Pointing SM (eds) Marine mycology—a practical approach. Fungal Diversity Research, Hong Kong, pp 1–20
Wang G, Li Q, Zhu P (2008) Phylogenetic diversity of culturable fungi associated with the Hawaiian sponges Suberites zeteki and Gelliodes fibrosa. A Van Leeuw 93:163–174
Weston NB, Joye SB (2005) Temperature-driven decoupling of the key phases of organic matter degradation in marine sediments. PNAS 102:17036–17040
White TJ, Bruns T, Lee S, Taylor DJ (1990) Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In: Innes MA, Gelfand DH, Sninsky JJ, White TJ (eds) PCR protocols. Academic, San Diego, pp 315–322
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Mouton, M., Postma, F., Wilsenach, J. et al. Diversity and Characterization of Culturable Fungi from Marine Sediment Collected from St. Helena Bay, South Africa. Microb Ecol 64, 311–319 (2012). https://doi.org/10.1007/s00248-012-0035-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00248-012-0035-9