
Abstract
Acidification is one of the most common and serious problems inducing process failure in anaerobic digesters. The production of volatile fatty acids (VFAs) mainly triggers acidic shock. However, little is known about the bacteria involved in the processes of acidogenic metabolism, such as fermentation and reductive acetogenesis. Here, the metabolic responses of a methanogenic community to the acidification and resulting process deterioration were investigated using transcriptional profiling of both the 16S rRNA and formyltetrahydrofolate synthetase (FTHFS) genes. The 16S rRNA-based analyses demonstrated that the dynamic shift of bacterial populations was closely correlated with reactor performance, especially with VFA accumulation levels. The pH drop accompanied by an increase in VFAs stimulated the metabolic activation of an uncultured Chloroflexi subphylum I bacterium. The subphylum has been characterized as a fermentative carbohydrate degrader using culture- and molecular-based ecophysiological assays. At the beginning of VFA accumulation, FTHFS genes were expressed; the transcripts were derived from phylogenetically predicted homoacetogens, suggesting that reductive acetogenesis was operated by hitherto unidentified bacteria. When acetate concentrations were high, the FTHFS expression ceased and Thermoanaerobacterium aciditolerans proliferated selectively. This thermoacidophilic bacterium would play a decisive role in acetate production via fermentative metabolism. The results of this study reveal for the first time that an uncultured Chloroflexi, T. aciditolerans, and novel homoacetogens were metabolically associated with acidic shock and subsequent VFA accumulation in an anaerobic digester.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Mutual and syntrophic interactions of functionally distinct microorganisms are crucial for the methanogenic conversion of organic matter [36, 48]. In anaerobic digesters, the methanogenic process consists of hydrolysis, fermentation, syntrophic oxidation, and methanogenesis. Bacterial groups catalyze the first three metabolic steps, while the last one is mediated by the methanogenic archaea. Because these degradation steps are highly associated with each other, an imbalance between the bacterial and archaeal populations causes the deterioration of reactor performance [26, 55]. It has been reported that various environmental factors, e.g., pH, temperature, substrate, and dilution rate, greatly affect the activity and composition of digester microbiota [12, 24, 30, 45].
An important objective for microbial ecologists studying anaerobic digesters is to predict and prevent unexpected process failure based on knowledge of the microbial community structure and function. Up to now, some deterioration events have been investigated intensively in order to elucidate the process stability [5, 6, 42]. For instance, the bulking of methanogenic granules has been recognized as a serious problem. The diversity and distribution of the deterioration-triggering microbes have been demonstrated using molecular- and culture-based approaches [43, 51, 54]. However, in general, process failures occur accidentally, which makes it difficult to clarify the microorganisms involved.
Acidification is one of the most common problems causing process deterioration in anaerobic digesters [46]. The sudden drop in pH is attributed to the accumulation of volatile fatty acids (VFAs) and the resulting breakthrough in the buffering capacity of digestive sludge. The bacterial groups have been understood to be involved mainly in the production of VFAs. The main pathway for acidogenesis is fermentative metabolism by anaerobic organotrophs. In addition, the acetyl-CoA pathway for reductive acetogenesis is another possible way to generate acetate, one of the most abundant VFAs in anaerobic environments. Hori et al. reported that the acidification process can be reproduced by the control of reactor-operating conditions in a laboratory-scale experiment [14]. DNA-based analyses revealed that the dynamic shift of methanogens was related to the concentration of VFAs. On the other hand, the response of bacterial populations to acidic shock remains to be clarified.
RNA-based fingerprinting methods have been proven adequate for monitoring metabolically active microorganisms [10, 19, 40]. Transcription of the 16S rRNA gene is involved in the global protein synthesis of microbiota, whereas the mRNA expression of specific functional genes captures the substrate-metabolizing activity of the functional guilds. The formyltetrahydrofolate synthetase (FTHFS) gene has been used as an ecological biomarker for reductive acetogenesis [13, 23, 29]. Thus, transcriptional profiling of both the 16S rRNA and FTHFS genes would provide deeper insight into the active acidogenic bacteria in anaerobic digesters. The aim of this study was to investigate the metabolic responses of a microbial population to the acidification and subsequent process deterioration in a thermophilic anaerobic digester using the transcriptional profiling of the 16S rRNA and FTHFS genes.
Materials and Methods
Operation of the Anaerobic Digester
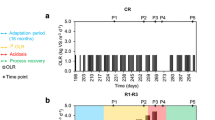
A seed culture was collected from a thermophilic anaerobic digester for treating garbage wastewater and was cultivated at 55°C in a 1.4 l agitation tank (Labo-controller MDL-8L; B. E. Marubishi, Tokyo, Japan) [15]. Synthetic wastewater containing glucose (1%, w/v) as a sole carbon and energy source was injected into the bioreactor at a dilution rate of 0.1 day−1. The composition of the wastewater was described in detail previously [14]. The minerals were supplemented once a day at the following final concentrations (mg l−1): FeSO4·7H2O, 5.53; CoCl2·6H2O, 0.48; ZnCl2, 0.67; CaCl2·2H2O, 0.59; CuCl2·2H2O, 0.16; MnCl2·4H2O, 2.02; H3BO4, 0.063; Na2MoO4·2H2O, 0.0045; and NiCl2·6H2O, 0.65. Prior to the initiation of the experiment, the reactor was operated stably with pH controlled at 7.1 by an automatic titration of 5 N NaOH. The pH control was stopped at the start of the experiment in order to induce a gradual decrease in pH. Subsequently, the pH naturally dropped because the buffering capacity of the wastewater broke down (Fig. 1). After the acidic shock, the pH value was maintained at 6.1 using the NaOH titration. A stepwise increase in pH was conducted to facilitate recovery from acidification. The reproducibility of induction of the acidification, deterioration, and recovery states was confirmed in the operation of parallel reactors. Gas production was measured daily on the basis of liquid displacement. The concentration of VFAs was determined at least once every 3 days using an HPLC (Alliance 2695; Waters, Tokyo, Japan) equipped with an OApak column (Tosoh, Tokyo, Japan) and a photodiode array (2996; Waters). The substrate carbon was recovered as the microbial cells (i.e., total suspended solids) and metabolites (e.g., VFAs and gases). Throughout the operation, the digestion slurries were sampled for molecular analyses.

Reactor performance during the operation of the thermophilic methanogenic process: open diamond pH, black square gas production rate, black triangle acetate concentration, open circle propionate concentration, black diamond butyrate concentration, and square lactate concentration. Arrows indicate the sampling points for the 16S rRNA-based T-RFLP and clone library analyses (in boldface). The reproducibility of reactor performances was confirmed in the operation of parallel reactors. The representative operation as the whole consensus was chosen for presentation
Extraction of Total Nucleic Acids and the Subsequent RT-PCR or PCR
Nucleic acids were extracted from the culture samples using a direct lysis protocol involving bead beating [37]. Total RNA was prepared by digestion with DNase (RQ1; Promega, Tokyo, Japan), and total DNA was purified with RNase treatment (Type II-A; Sigma-Aldrich, Tokyo, Japan). RT-PCR and PCR were conducted for terminal restriction fragment-length polymorphism (T-RFLP) [28] and clone library analyses. For the 16S rRNA-based analysis, primer set Ba27f/Ba907r was used for bacteria, and a set of Ar109f/Ar912rt was used for archaea [33, 34]. The primer set for the FTHFS genes was described previously [29]. For T-RFLP, the 16S rRNA primers (Ba27f and Ar912rt) and the FTHFS-targeted forward primer were covalently labeled with 6-carboxyfluorescein (FAM). RT-PCR of the 16S rRNA was carried out using a Primescript one-step RT-PCR kit (Takara, Tokyo, Japan). A SuperScript III one-step RT-PCR with Platinum Taq (Invitrogen, Tokyo, Japan) was used for RT-PCR of the FTHFS-coding mRNA. The PCR amplification was performed with AmpliTaqGold (Applied Biosystems, Tokyo, Japan). The thermal conditions of the 16S rRNA-based PCR were as follows: pre-denaturation at 94°C for 10 min, and then 20 cycles of denaturation at 94°C for 30 s, annealing at 52°C for 45 s, and elongation at 72°C for 90 s; and a final extension step at 72°C for 5 min. During RT-PCR of the 16S rRNA, reverse transcription was performed at 48°C for 45 min, followed by the PCR program. The thermal cycle conditions of the FTHFS-based PCR were described previously [29]. The thermal conditions of the FTHFS RT-PCR were reported by Pester and Brune [38]. We conducted reverse transcription at 55°C for 45 min and 30 cycles in the standard PCR program, during which the annealing temperature was 55°C. A high temperature (55°C) for the reverse transcription and annealing steps was implemented to avoid the formation of nonspecific amplicons. The absence of DNA contamination was confirmed by RT-PCR without reverse transcriptase.
T-RFLP Analyses Based on the 16S rRNA and FTHFS Genes and Their Transcripts
The RT-PCR and PCR amplicons were purified using a Wizard SV Gel and PCR Clean-Up System (Promega). The bacterial and archaeal 16S rRNA fragments were digested with restriction enzymes MspI and TaqI (New England Biolabs, Tokyo, Japan), respectively. The FTHFS gene amplicons were digested simultaneously with TaqI and RsaI (New England Biolabs). One microliter of the restriction digests was added to 15 μl Hi-Di formamide (Applied Biosystems) containing 0.4 μl of a GeneScan 500 or 1,000 (Applied Biosystems) as the internal size standard. Prior to starting electrophoresis, the mixture was heated at 95°C for 3 min and chilled on ice. Terminal restriction fragments (T-RFs) were separated by capillary electrophoresis with a 3130xl genetic analyzer (Applied Biosystems). T-RFLP profiles were analyzed on the basis of the peak size and area using GeneMapper software, version 3.7 (Applied Biosystems). A T-RF with more than 1% abundance to the total peak area was treated as a distinct one [9]. T-RFLP was conducted in triplicate and the mean of the relative abundances of T-RFs was used for presentation.
Statistical Analyses of the 16S rRNA-Based T-RFLP Data
To investigate the bacterial transition during the reactor operation, the T-RFLP profiles based on the bacterial 16S rRNA genes and transcripts were applied to principal component analysis (PCA) using JMP software, version 5.1 (SAS Institute, Tokyo, Japan). The length and relative abundance of T-RFs were used for the statistical processing. After a covariance data matrix was calculated by using pairwise case deletion and varimax rotation methods, the resulting data were plotted in a two-dimensional map.
Cloning and Sequencing of the 16S rRNA and FTHFS Gene Transcript
To phylogenetically identify a whole range of metabolically active bacteria and archaea, the bacterial 16S rRNA from days 26, 33, 38, and 43 and the archaeal rRNA from day 43 were subjected to the RT-PCR, cloning, and sequencing. The transcription of the FTHFS genes was investigated when the concentrations of acetate in the culture supernatant were 2.1, 9.6, 21.4, and 31.1 mM; the RT-PCR product was not obtained at the concentration of 31.1 mM. Thus, the clone library analysis was conducted at the lower three concentrations. The PCR and RT-PCR conditions were the same as mentioned above. The PCR amplicons were purified using a QIAquick PCR purification kit (Qiagen, Tokyo, Japan), and then ligated into a plasmid vector pGEM-T Easy (Promega) according to the manufacturer’s instructions. Escherichia coli JM 109 supercompetent cells (Toyobo, Tokyo, Japan) were transformed with the plasmid. The DNA segments were extracted using a GenElute Plasmid Miniprep Kit (Sigma-Aldrich) and used as templates for sequencing reactions with a BigDye Terminator v3.1 Cycle Sequencing kit (Applied Biosystems). Sequencing was carried out using the 3130xl genetic analyzer (Applied Biosystems).
Phylogenetic Analysis of the Sequence Data
The absence of the chimeric structure was checked by phylogenetically analyzing the terminal stretches at 5′ and 3′ ends [31, 32]. The 16S rRNA clones with at least 97% sequence similarity were grouped in an operational taxonomic unit (OTU) [1, 47]. For the FTHFS analysis, clones with >93% amino acid sequence identity were treated as an identical phylotype and clones with at least 99.5% of the gene sequence similarity were defined as belonging to the same genotype [38]. A representative sequence from each OTU and genotype was deposited in the database. The cultured relative was determined by comparing the gene sequence with those in the DDBJ nucleotide sequence database using the BLAST program. For the 16S rRNA, the phylogenetic tree was constructed using ARB software (http://www.arb-home.de) [32]. The sequence data were aligned automatically and then corrected manually. Base lineage trees were constructed by neighbor-joining, maximum parsimony, and maximum likelihood methods using reference 16S rRNA gene sequences with >1,400 nucleotides. In order to reveal the phylogenetic position of partial 16S rRNA sequences, the aligned sequences were added to the trees by using the ARB parsimony tool. For the FTHFS gene, the deduced amino acid sequences were aligned using ClustalX software, version 2.00 [50]. After the alignment gaps were ruled out manually, phylogenetic trees were constructed by the neighbor-joining and maximum likelihood methods using Phylip software, version 3.68 [11]. In both gene cases, use of the different tree construction algorithms resulted in similar dendrogram topology. Bootstrap values were obtained from 1,000 replications.
Accession Numbers
The nucleotide sequence data obtained in this study have been deposited in the DDBJ nucleotide sequence database under accession numbers AB516974 to AB516992 for the 16S rRNA gene and AB517149 to AB517153 for the FTHFS gene.
Results
Monitoring of Reactor Performance in the Anaerobic Digester
Physicochemical parameters of reactor performance were monitored throughout the operation (Fig. 1). The reproducibility of the acidification and recovery processes was confirmed by the operation of parallel reactors. The pH decreased gradually from 7.1 to 6.6 during the first 23 days, and suddenly dropped to 6.1 at day 25. The concentration of acetate was <1.7 mmol l−1 for the first 20 days, and afterwards, it increased from 4.3 mmol l−1 at day 23 to 52.0 mmol l−1 at day 36. The propionate concentration increased from 0.3 mmol l−1 at the beginning of the operation to 1.6 mmol l−1 at day 23. After the acidic shock, propionate was accumulated again to the concentration of 4.6 mmol l−1 at day 31; the concentration of propionate remained at this level for the remainder of the experiment. The pH drop stimulated the butyrate and lactate accumulation; the concentration of these acids transiently increased and decreased within a range of 0 to 2.2 mmol l−1. The gas production rate fluctuated within 160 to 390 ml day−1 during the first 23 days and then decreased from 210 ml day−1 at day 25 to 70 ml day−1 at day 28. During the following 8 days, the gas production rate remained approximately 100 ml day−1. After the pH was increased to 6.3 at day 37, the accumulated VFAs (except propionate) were degraded; in particular, a rapid degradation of acetate was observed from days 37 to 43. The degradation of VFAs coincided with vigorous gas production and a spontaneous increase in the pH to the neutral range from day 39. More than 80% of the glucose carbon was recovered in microbial cells, VFAs, and gases (data not shown).
Community Transition of Bacteria and Archaea Using the 16S rRNA-Based T-RFLP
The microbial community succession during the operation was examined using the RNA- and DNA-based T-RFLP (Fig. 2). A total of 16 T-RFs were detected in both the bacterial RNA and DNA profiles; however, the composition and abundance differed considerably from each other (Fig. 2a and c). The RNA analysis revealed a drastic change in T-RFs such as 148, 521, and 161 bp. On only the RNA fingerprint, T-RF of 521 bp appeared (relative abundance: 5.6–8.3%) from days 23 to 29, during which the pH dropped and acetate accumulated at concentrations of 4.1–26.0 mM. The PCA indicated a grouping of these days on the right side of the plot (Fig. 3a). Days 33–40, when acetate concentrations were >39.1 mM, were characterized by a proliferation of the 161-bp T-RF (41–66% of the total peak area). These days are closely located at the lower left on the PCA plot. After the degradation of accumulated acetate, T-RFs of 171 and 466 bp appeared, and those of 159 and 167 bp reappeared. Day 43 is mapped at the upper part of the PCA plot, which is distant from days 33–40 and rather close to the start of the operation. On the other hand, the DNA-based T-RFLP showed that T-RF of 289 bp was dominant, accounting for ∼88% of the total. Based on the PCA, days 0 and 8 are distinct from each other and any other days (Fig. 3b). Days 17 to 43 are located closely at the upper left on the plot; however, the population was slightly changed at day 38. The T-RFLP pattern at day 38 is rather similar to those at days 17, 23, and 26. This may have been caused by the exclusive predominance of the 171-bp T-RF and seems to have little relationship with reactor performance. The start and end points of the operation are far from each other on the plot. With respect to archaeal dynamics, no significant difference was observed between the RNA and DNA profiles (Fig 2b and d). During the operation, the 92-bp T-RF was most abundant (54–91% of the total). The abundance of the 187-bp T-RF decreased along with the acidification process. From days 29 to 40, when the pH was maintained at an acidic range (pH, 6.1–6.4), the 187-bp peak was not detected in the DNA profile, while a small portion of the peak remained in the RNA profile. Only on the RNA fingerprint, the 381-bp T-RF appeared at day 43.

Microbial community dynamics as determined by the RNA- and DNA-based T-RFLP. a Bacterial fingerprint from the 16S rRNA. b Archaeal fingerprint from the rRNA. c Bacterial fingerprint from the rRNA gene. d Archaeal fingerprint from the rRNA gene. The possible OTU corresponding to T-RF is presented in parentheses and boldface. The T-RFLP was conducted in triplicate, and the mean of the relative abundances of T-RFs was used for presentation

Principal component analysis (PCA) of the bacterial T-RFLP data. a PCA plot for the 16S rRNA. b PCA plot for the 16S rRNA gene. The successive time points (days) are connected by arrows and indicated by numbers
Phylogenetic Identification of the Acidic Deterioration-Associated Microbiota
To phylogenetically identify the metabolically active microorganisms during the reactor operation, clone libraries from the 16S rRNA were constructed. For bacteria (Table 1), (a) the Bd26 library (n = 47 clones) from day 26, (b) the Bd33 library (n = 47) from day 33, (c) the Bd38 library (n = 48) from day 38, and (d) the Bd43 library (n = 52) from day 43; for archaea, (e) the Ad43 library (n = 48) from day 43. A total of 14 OTUs were obtained from the bacterial libraries. In the Bd26 library, OTU B1 was detected as a main component (21% of the total). The expected T-RF size is 521 bp, which corresponds to the dominant T-RF from days 23 to 29 (Fig. 2a). The OTU had no cultured representative and formed a novel branch in the phylogenetic tree within the phylum Chloroflexi (Fig. 4a). In the Bd33 and Bd38 libraries, 52–59% of the clones belonged to Thermoanaerobacterium aciditolerans (AY350594; 98% similarity; Fig. 4a). The T-RF is expected to be 161 bp, which matches with the primary T-RF from days 33 to 40 (Fig. 2a). In addition, the Bd38 library included OTU B5, which was affiliated with Moorella thermoacetica (AY884087; 98% similarity). The T-RF is estimated to be 148 bp, which was found by the RNA T-RFLP at days 38 and 40 (Fig. 2a). The Bd43 library contained OTU B8. The predicted T-RF is 466 bp, corresponding to the major T-RF at day 43 (Fig. 2a). In the archaeal library, OTUs A1 and A2 were the dominant members (69% and 25% of the total, respectively). OTU A1 was identified as the hydrogenotrophic methanogen Methanothermobacter thermautotrophicus (DQ657903; 100% similarity). The expected T-RF is 92 bp, corresponding to the primary T-RF detected (Fig. 2b and d). OTU A2 was affiliated within an uncultured phylogenetic group, the so-called rice cluster III (Fig. 4b). They were closely related to the uncultured euryarchaeote clone 1C (GQ365371; 99% similarity). The expected T-RF is 381 bp, representing the distinctive appearance at the end of the reactor operation. Minor OTUs belonged to M. thermautotrophicus (AY196660; 95% similarity), Methanoculleus thermophilus (EF118904; 99% similarity), and Methanosarcina thermophila (M59140; 98% similarity); the respective T-RFs are estimated to be 393, 84, and 187 bp.

Phylogenetic tree showing the relationship of the 16S rRNA phylotypes (i.e., OTUs B1 and B2 [A] and OTU A2 [B]). The base lineage tree was constructed using the maximum parsimony method. Bootstrap values were obtained from 1,000 replications, and >90%, 70 to 89%, and <69% are shown with black, gray, and open circles, respectively. The scale bar represents 10% sequence divergence. The GenBank accession numbers of the reference sequences are given. The subphylum-level groups in the phylum Chloroflexi are numbered in brackets according to a previous report [18]
Expression Profile of the FTHFS Genes by the T-RFLP and Clone Library Analyses
Under acidic conditions (pH 6.1–6.4), acetate was formed as the main microbial metabolite, and other VFAs, such as propionate and/or butyrate, were also found (Fig. 1). The FTHFS genes were transcribed at acetate concentrations (mM) of 2.1, 9.6, and 21.4, while expression was not detected at the concentration of 31.1 mM. T-RFLP was conducted to characterize the FTHFS-expressing bacteria. The T-RFLP patterns of DNA were quite simple; only one predominant T-RF of 329 bp was observed. In the mRNA T-RFLP, the 329-bp T-RF was also a main constituent. Additionally, the 138- and 102-bp T-RFs were detected at the acetate concentration of 2.1 mM. The clone library analysis phylogenetically identified these T-RFs (Table 2). A total of two phylotypes were obtained in the libraries. Phylotype A, corresponding to the 329-bp T-RF, had no cultured representative, and the sequence identity of amino acids was 73.0% to the homoacetogen Thermoanaerobacter kivui (formerly called Acetogenium kivui) [25]. The other phylotype B (T-RF of 138 bp) was also novel, and the sequence identity of amino acids was 79.0% to the homoacetogen M. thermoacetica [8]. The phylotypes formed separate branches that clustered with sequences from known homoacetogens (Fig. 5). Recently, Henderson et al. proposed a “Homoacetogen similarity” (HS) score on the basis of conserved FTHFS residues that are diagnostic for homoacetogenic bacteria [13]. An FTHFS sequence with high HS scores may have originated from homoacetogen. The HS score from phylotypes A and B was calculated to be >75%, higher than the score (<60%) from bacterial groups containing non-homoacetogenic isolates [13]. The clones representing the 102-bp T-RF were detected during the construction of the mRNA-based library; however, these were identified to be chimerical and thus, ruled out for further examinations.

Phylogenetic tree showing the relationships of the FTHFS phylotypes A and B (presented in boldface). Gray zone indicates non-homoacetogens. The deduced amino acid sequences were used to construct the tree using the neighbor-joining method. Bootstrap values were obtained from 1,000 replications, and >90%, 70 to 89%, and 50% to 69% are shown in black, gray, and open circles, respectively. The bar represents 10% sequence divergence. The GenBank accession numbers of the reference sequences are shown
Discussion
In this study, we investigated metabolically active microbiota during the acidification processes in a thermophilic anaerobic digester. The reproducible induction of acidic reactor conditions formed the foundation of this study. The transcription profiling of the 16S rRNA gene revealed a distinctive appearance of putatively fermentative bacteria (i.e., an uncultured Chloroflexi subphylum I bacterium, T. aciditolerans, and M. thermoacetica) along with the accumulation of VFAs. Furthermore, the expression analysis of the FTHFS genes demonstrated that novel and phylogenetically presumed homoacetogens may have been involved in reductive acetogenesis during the acidification process.
A novel Chloroflexi subphylum I bacterium (OTU B1, T-RF of 521 bp) was metabolically active from days 23 to 29 (Figs. 2a and 4a), during which the pH dropped and acetate accumulated at concentrations of 4.1–26.0 mmol l−1. This OTU was, to some extent, related to the filamentous Anaerolinea thermophila (AB046413; 82% sequence similarity), which proliferated during the granule balking in UASB-type anaerobic digesters [44]. The uncultured subphylum I bacteria have also been found as the dominant members in the submerged membrane reactor (e.g., environmental clones CMBR-2 and CMBR-3 in Fig. 4a) [35]. Owing to physiological assays on a wide range of subphylum I bacteria [35, 44, 52, 53], the phylogenetic group might be capable of fermenting carbohydrates under anaerobic conditions. At present, it is unknown why these appeared distinctively at the acidic shock. However, it is noted that the phenomenon can be potentially indicative of initiating the sudden decrease of pH. The FTHFS genes were transcribed at acetate concentrations of 2.1–21.4 mmol l−1 (Table 2). Higher accumulation of acetate might interfere with FTHFS expression. The main transcript, phylotype A, was presumed to be a novel and homoacetogenic bacterium because of its phylogenetic placement (Fig. 5) and high HS score (>75%). Under acidic conditions, the concentration of dissolved H2 might be high [4, 41]. Acetogens could utilize H2/CO2 for reductive acetogenesis even if hydrogenotrophic methanogens coexisted [56]. The acetyl-CoA pathway has been known to be operational in acetate assimilation [7], and thereby, the FTHFS expression can reflect the outgrowth of such bacteria. However, the temporal expression of specific FTHFS genes (i.e., phylotypes A and B) rules out the possibility that the expression was accounted for by assimilative metabolism. These findings strongly suggest that reductive acetogenesis by the specific bacterial lineages contributed to acetate production at the beginning of the process deterioration. The precise and quantitative evaluation of reductive acetogenesis can be done by the isotope probing technique and/or cultivation to directly link the microbial phylogeny and physiology.
T. aciditolerans (OTU B2, T-RF of 161 bp) was activated selectively at high concentrations of acetate (>39.1 mmol l−1) from days 33 to 40 (Figs. 2a and 4a). This bacterium is able to ferment a variety of mono- and disaccharides including glucose under acidic and thermophilic conditions (i.e., within a pH range of 3.2–7.1 and a temperature range of 45–68°C) [22]. The primary products of glucose fermentation are acetate and lactate, which were found during the deteriorative phase. Therefore, fermentative bacterium OTU B2 was expected to have a direct role in VFA production in the acidic digester (pH, 6.1–6.4). At this time, M. thermoacetica (OTU B5, T-RF of 148 bp) was also metabolically active. The OTU might engage in fermentative metabolism rather than reductive acetogenesis [8] because the FTHFS expression was not observed with the high acetate accumulation. With respect to archaeal dynamics, the aceticlastic methanogen Methanosarcina sp. (OTU A5, the 187-bp T-RF) disappeared from the acidic digester (Fig. 2d), which is consistent with the fact that, thus far, all detected acidophilic methanogens have been hydrogenotrophic [2, 3, 21].
Active bacterial and archaeal populations were established as a result of the recovery from acidification. Uncultured bacteria (OTUs B11 and B8) may be possible candidates of VFA-oxidizing syntroph [39, 49]; however, they both showed low sequence similarities of the 16S rRNA to those from cultured representatives (Table 1). Further research is needed to determine whether or not these bacteria are the VFA-oxidizing syntroph. Furthermore, a novel uncultured archaeon (OTU A2, T-RF of 381 bp) became active just after acetate degradation (Figs. 2b and 4b). Its physiological propensities have not been identified yet. However, in another reactor perturbed by organic loading, the related sequence (Clone D204_2 in Fig. 4b) was obtained after the decomposition of acetate (data not shown). And the relative clone (euryarchaeote clone 1C in Fig. 4b) was found in acetate-degrading enrichment [27]. It is tempting to think that this organism is related to the anaerobic metabolism of acetate. An enrichment of the rice cluster III organisms (clones LL25A1 and LL37A19 in Fig. 4b) has been investigated by cultivation and molecular approaches [20]. Those authors suggested that the uncultured archaeon grew heterotrophically on peptides; nevertheless, this is one possibility.
The results of this study demonstrate that the dynamic shifts of metabolically active microbiota were closely correlated with the reactor performance, especially with the acidic shock and acetate accumulation levels. The transcriptional profiling of the 16S rRNA and FTHFS genes showed that an uncultured Chloroflexi bacterium, T. aciditolerans, and novel homoacetogens were responsible for acidogenic metabolism (i.e., fermentation and reductive acetogenesis) during the acidification process. Following the present study, the key acidogenic bacteria will be directly identified by stable isotope probing [16, 17]. Clarification of the ecophysiological features of deterioration-associated microorganisms should contribute to our understanding of the mechanism underlying the process stability.
References
Boone DR, Whitman WB, Rouviere P (1993) Diversity and taxonomy of methanogens. In: Ferry JG (ed) methanogens. Chapman and Hall, inc., New York, pp 35–80
Brauer SL, Cadillo-Quiroz H, Yashiro E, Yavitt JB, Zinder SH (2006) Isolation of a novel acidiphilic methanogen from an acidic peat bog. Nature 442:192–194
Cadillo-Quiroz H, Yashiro E, Yavitt JB, Zinder SH (2008) Characterization of the archaeal community in a minerotrophic fen and terminal restriction fragment length polymorphism-directed isolation of a novel hydrogenotrophic methanogen. Appl Environ Microbiol 74:2059–2068
Cord-Ruwisch R, Mercz TI, Hoh C-Y, Strong GE (1997) Dissolved hydrogen concentration as an on-line control parameter for the automated operation and optimization of anaerobic digesters. Biotechnol Bioeng 56:626–634
Cresson R, Dabert P, Bernet N (2009) Microbiology and performance of a methanogenic biofilm reactor during the start-up period. J Appl Microbiol 106:863–876
Delbes C, Moletta R, Godon J (2001) Bacterial and archaeal 16S rDNA and 16S rRNA dynamics during an acetate crisis in an anaerobic digestor ecosystem. FEMS Microbiol Ecol 35:19–26
Drake HL (1994) Acetogenesis. Chapman and Hall, New York
Drake HL, Daniel SL (2004) Physiology of the thermophilic acetogen Moorella thermoacetica. Res Microbiol 155:869–883
Dunbar J, Ticknor LO, Kuske CR (2001) Phylogenetic specificity and reproducibility and new method for analysis of terminal restriction fragment profiles of 16S rRNA genes from bacterial communities. Appl Environ Microbiol 67:190–197
Eichler S, Christen R, Holtje C, Westphal P, Botel J, Brettar I, Mehling A, Hofle MG (2006) Composition and dynamics of bacterial communities of a drinking water supply system as assessed by RNA- and DNA-based 16S rRNA gene fingerprinting. Appl Environ Microbiol 72:1858–1872
Felsenstein J (2005) PHYLIP (phylogeny inference package). Department of Genome Sciences, University of Washington, Seattle
Goberna M, Insam H, Franke-Whittle IH (2009) Effect of biowaste sludge maturation on the diversity of thermophilic bacteria and archaea in an anaerobic reactor. Appl Environ Microbiol 75:2566–2572
Henderson G, Naylor GE, Leahy SC, Janssen PH (2010) Presence of novel, potentially homoacetogenic bacteria in the rumen as determined by analysis of formyltetrahydrofolate synthetase sequences from ruminants. Appl Environ Microbiol 76:2058–2066
Hori T, Haruta S, Ueno Y, Ishii M, Igarashi Y (2006) Dynamic transition of a methanogenic population in response to the concentration of volatile fatty acids in a thermophilic anaerobic digester. Appl Environ Microbiol 72:1623–1630
Hori T, Haruta S, Ueno Y, Ishii M, Igarashi Y (2006) Direct comparison of single-strand conformation polymorphism (SSCP) and denaturing gradient gel electrophoresis (DGGE) to characterize a microbial community on the basis of 16S rRNA gene fragments. J Microbiol Methods 66:165–169
Hori T, Noll M, Igarashi Y, Friedrich MW, Conrad R (2007) Identification of acetate-assimilating microorganisms under methanogenic conditions in anoxic rice field soil by comparative stable isotope probing of RNA. Appl Environ Microbiol 73:101–109
Hori T, Muller A, Igarashi Y, Conrad R, Friedrich MW (2010) Identification of iron-reducing microorganisms in anoxic rice paddy soil by 13C-acetate probing. ISME J 4:267–278
Hugenholtz P, Stackebrandt E (2004) Reclassification of Sphaerobacter thermophilus from the subclass Sphaerobacteridae in the phylum Actinobacteria to the class Thermomicrobia (emended description) in the phylum Chloroflexi (emended description). Int J Syst Evol Microbiol 54:2049–2051
Juottonen H, Tuittila ES, Juutinen S, Fritze H, Yrjala K (2008) Seasonality of rDNA- and rRNA-derived archaeal communities and methanogenic potential in a boreal mire. ISME J 2:1157–1168
Kemnitz D, Kolb S, Conrad R (2005) Phenotypic characterization of Rice Cluster III archaea without prior isolation by applying quantitative polymerase chain reaction to an enrichment culture. Environ Microbiol 7:553–565
Kotsyurbenko OR, Friedrich MW, Simankova MV, Nozhevnikova AN, Golyshin PN, Timmis KN, Conrad R (2007) Shift from acetoclastic to H2-dependent methanogenesis in a west Siberian peat bog at low pH values and isolation of an acidophilic Methanobacterium strain. Appl Environ Microbiol 73:2344–2348
Kublanov IV, Prokofeva MI, Kostrikina NA, Kolganova TV, Tourova TP, Wiegel J, Bonch-Osmolovskaya EA (2007) Thermoanaerobacterium aciditolerans sp. nov., a moderate thermoacidophile from a Kamchatka hot spring. Int J Syst Evol Microbiol 57:260–264
Leaphart AB, Friez MJ, Lovell CR (2003) Formyltetrahydrofolate synthetase sequences from salt marsh plant roots reveal a diversity of acetogenic bacteria and other bacterial functional groups. Appl Environ Microbiol 69:693–696
Lee C, Kim J, Hwang K, O’Flaherty V, Hwang S (2009) Quantitative analysis of methanogenic community dynamics in three anaerobic batch digesters treating different wastewaters. Water Res 43:157–165
Leigh JA, Mayer F, Wolfe RS (1981) Acetogenium kivui, a new thermophilic hydrogen-oxidizing acetogenic bacterium. Arch Microbiol 129:275–280
Lettinga G (1995) Anaerobic digestion and wastewater treatment systems. Antonie Leeuwenhoek 67:3–28
Lins P, Malin C, Wagner AO, Illmer P (2010) Reduction of accumulated volatile fatty acids by an acetate-degrading enrichment culture. FEMS Microbiol Ecol 71:469–478
Liu WT, Marsh TL, Cheng H, Forney LJ (1997) Characterization of microbial diversity by determining terminal restriction fragment length polymorphisms of genes encoding 16S rRNA. Appl Environ Microbiol 63:4516–4522
Lovell CR, Leaphart AB (2005) Community-level analysis: key genes of CO2-reductive acetogenesis. Methods Enzymol 397:454–469
Lu F, Shao LM, Bru V, Godon JJ, He PJ (2009) Synergetic effect of pH and biochemical components on bacterial diversity during mesophilic anaerobic fermentation of biomass-origin waste. J Appl Microbiol 106:580–591
Ludwig W, Strunk O, Klugbauer S, Klugbauer N, Weizenegger M, Neumaier J, Bachleitner M, Schleifer KH (1998) Bacterial phylogeny based on comparative sequence analysis. Electrophoresis 19:554–568
Ludwig W, Strunk O, Westram R, Richter L, Meier H, Yadhukumar BA, Lai T, Steppi S, Jobb G, Forster W, Brettske I, Gerber S, Ginhart AW, Gross O, Grumann S, Hermann S, Jost R, Konig A, Liss T, Lussmann R, May M, Nonhoff B, Reichel B, Strehlow R, Stamatakis A, Stuckmann N, Vilbig A, Lenke M, Ludwig T, Bode A, Schleifer KH (2004) ARB: a software environment for sequence data. Nucleic Acids Res 32:1363–1371
Lueders T, Friedrich MW (2002) Effects of amendment with ferrihydrite and gypsum on the structure and activity of methanogenic populations in rice field soil. Appl Environ Microbiol 68:2484–2494
Lueders T, Wagner B, Claus P, Friedrich MW (2004) Stable isotope probing of rRNA and DNA reveals a dynamic methylotroph community and trophic interactions with fungi and protozoa in oxic rice field soil. Environ Microbiol 6:60–72
Miura Y, Watanabe Y, Okabe S (2007) Significance of Chloroflexi in performance of submerged membrane bioreactors (MBR) treating municipal wastewater. Environ Sci Technol 41:7787–7794
Narihiro T, Sekiguchi Y (2007) Microbial communities in anaerobic digestion processes for waste and wastewater treatment: a microbiological update. Curr Opin Biotechnol 18:273–278
Noll M, Matthies D, Frenzel P, Derakshani M, Liesack W (2005) Succession of bacterial community structure and diversity in a paddy soil oxygen gradient. Environ Microbiol 7:382–395
Pester M, Brune A (2006) Expression profiles of fhs (FTHFS) genes support the hypothesis that spirochaetes dominate reductive acetogenesis in the hindgut of lower termites. Environ Microbiol 8:1261–1270
Qiu YL, Sekiguchi Y, Hanada S, Imachi H, Tseng IC, Cheng SS, Ohashi A, Harada H, Kamagata Y (2006) Pelotomaculum terephthalicum sp. nov. and Pelotomaculum isophthalicum sp. nov.: two anaerobic bacteria that degrade phthalate isomers in syntrophic association with hydrogenotrophic methanogens. Arch Microbiol 185:172–182
Roy CS, Talbot G, Topp E, Beaulieu C, Palin MF, Masse DI (2009) Bacterial community dynamics in an anaerobic plug-flow type bioreactor treating swine manure. Water Res 43:21–32
Schink B (1997) Energetics of syntrophic cooperation in methanogenic degradation. Microbiol Mol Biol Rev 61:262–280
Schoen MA, Sperl D, Gadermaier M, Goberna M, Franke-Whittle I, Insam H, Ablinger J, Wett B (2009) Population dynamics at digester overload conditions. Bioresour Technol 100:5648–5655
Sekiguchi Y, Takahashi H, Kamagata Y, Ohashi A, Harada H (2001) In situ detection, isolation, and physiological properties of a thin filamentous microorganism abundant in methanogenic granular sludges: a novel isolate affiliated with a clone cluster, the green non-sulfur bacteria, subdivision I. Appl Environ Microbiol 67:5740–5749
Sekiguchi Y, Yamada T, Hanada S, Ohashi A, Harada H, Kamagata Y (2003) Anaerolinea thermophila gen. nov., sp. nov. and Caldilinea aerophila gen. nov., sp. nov., novel filamentous thermophiles that represent a previously uncultured lineage of the domain Bacteria at the subphylum level. Int J Syst Evol Microbiol 53:1843–1851
Shigematsu T, Tang Y, Kobayashi T, Kawaguchi H, Morimura S, Kida K (2004) Effect of dilution rate on metabolic pathway shift between aceticlastic and nonaceticlastic methanogenesis in chemostat cultivation. Appl Environ Microbiol 70:4048–4052
Speece RE (1996) Anaerobic biotechnology for industrial wastewaters. Archae, Nashville
Stackebrandt E, Goebel BM (1994) A place for DNA–DNA reassociation and 16S rRNA sequence analysis in the present species definition in bacteriology. Int J Syst Bacteriol 44:846–849
Talbot G, Topp E, Palin MF, Masse DI (2008) Evaluation of molecular methods used for establishing the interactions and functions of microorganisms in anaerobic bioreactors. Water Res 42:513–537
Thevenieau F, Fardeau ML, Ollivier B, Joulian C, Baena S (2007) Desulfomicrobium thermophilum sp. nov., a novel thermophilic sulphate-reducing bacterium isolated from a terrestrial hot spring in Colombia. Extremophiles 11:295–303
Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, Higgins DG (1997) The CLUSTAL_X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res 25:4876–4882
Yamada T, Sekiguchi Y, Imachi H, Kamagata Y, Ohashi A, Harada H (2005) Diversity, localization, and physiological properties of filamentous microbes belonging to Chloroflexi subphylum I in mesophilic and thermophilic methanogenic sludge granules. Appl Environ Microbiol 71:7493–7503
Yamada T, Sekiguchi Y, Hanada S, Imachi H, Ohashi A, Harada H, Kamagata Y (2006) Anaerolinea thermolimosa sp. nov., Levilinea saccharolytica gen. nov., sp. nov. and Leptolinea tardivitalis gen. nov., sp. nov., novel filamentous anaerobes, and description of the new classes Anaerolineae classis nov. and Caldilineae classis nov. in the bacterial phylum Chloroflexi. Int J Syst Evol Microbiol 56:1331–1340
Yamada T, Imachi H, Ohashi A, Harada H, Hanada S, Kamagata Y, Sekiguchi Y (2007) Bellilinea caldifistulae gen. nov., sp. nov. and Longilinea arvoryzae gen. nov., sp. nov., strictly anaerobic, filamentous bacteria of the phylum Chloroflexi isolated from methanogenic propionate-degrading consortia. Int J Syst Evol Microbiol 57:2299–2306
Yamada T, Yamauchi T, Shiraishi K, Hugenholtz P, Ohashi A, Harada H, Kamagata Y, Nakamura K, Sekiguchi Y (2007) Characterization of filamentous bacteria, belonging to candidate phylum KSB3, that are associated with bulking in methanogenic granular sludges. ISME J 1:246–255
Zehnder AJB (1978) Ecology of methane formation. In: Mitchell R (ed) Water pollution microbiology, vol 2. Wiley, London, pp 349–376
Zehnder AJB, Stumm W (1988) Geochemistry and biochemistry of anaerobic habitats. In: Zehnder AJB (ed) Biology of anaerobic microorganisms. Wiley Interscience, New York, pp 1–38
Author information
Authors and Affiliations
Corresponding author
Additional information
Masateru Akuzawa and Tomoyuki Hori contributed equally to this work.
Rights and permissions
About this article
Cite this article
Akuzawa, M., Hori, T., Haruta, S. et al. Distinctive Responses of Metabolically Active Microbiota to Acidification in a Thermophilic Anaerobic Digester. Microb Ecol 61, 595–605 (2011). https://doi.org/10.1007/s00248-010-9788-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00248-010-9788-1