
Abstract
Virus production in oxic surface sediments and virioplankton sorption to suspended particles was estimated across three stations in the Southern California region (33.4°N, 118.6°W). Viriobenthos production was estimated using a sterile sediment and filtered porewater dilution technique that targeted production from both attached bacteria and bacteria living free in the porewater, and attached bacteria alone. Potential virus production rates by bacteria free in the porewater ranged from 1.7 to 4.6 × 108 VLP cm−3 h−1, while attached bacteria had slower potential production rates of between 0.4 and 1.1 × 108 VLP cm−3 h−1, suggesting turnover rates of viruses in sediments (1–5 h) which are significantly higher than those of virioplankton (~24–48 h). Virioplankton adsorbed to small (<150 µm) suspended sediments at stations with high ambient suspended solid concentrations. Virioplankton scavenging rates combined with published sedimentation rates demonstrate that this mechanism of virus arrival could only account for 0.01% of daily benthic virus production. Calculated mortality rates of benthic bacteria (4–14% h−1) suggest viruses may play an important role in sediment carbon cycling.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Viruses have high abundances in both marine waters and sediments, ranging from 106 to 108 mL−1 for pelagic viruses (reviewed in [12, 47]) and 106–1010 cm−3 for benthic viruses (reviewed in [19]). Viruses have been shown to be responsible for a considerable fraction of pelagic microbial mortality. Typically 10–32% of bacteria [32, 33] and between 2% and 3% of certain phytoplankton species [41] are lysed per day as a consequence of virus infection. Consequently viruses have important effects on biogeochemistry [4] and potentially host diversity [36]. The abundance of benthic viruses and respective hosts has received recent attention since these environments are important biogeochemically [3] and they are a sink for suspended matter in the sea [6]. Viriobenthos (benthic virus) abundance has been described in freshwater (Lac Gilbert, Canada [26]), estuarine (Chesapeake Bay, USA; Brisbane River and Moreton Bay estuaries, Australia [9, 19]), neritic (Florida Bay and Saanich Peninsula [2, 31]), and abyssal plain (Ieraptera Trench, Meditteranean [7, 8]) sediments. The origin of these viruses is unknown since these studies did not identify hosts [26]. Viral abundance in sediment mirrors bacterial abundance and production [2, 9]; hence most probably infect co-occurring benthic bacteria and archaea [2]. However, viruses that affect protists in sediments have also been documented [20].
There are several possible origins of viruses in marine sediments. Viruses may arise as a consequence of infection of benthic microorganisms (autochthonous production). Viruses in the water column may adsorb to particles and sink to sediments (allochthonous production). Infected bacteria in the last phase of intracellular virus assembly may be consumed by metazoa, then excreted as feces which settle to sediments. This latter mechanism is not considered a major input to sediments since only 1–2% of viruses are at this stage at any one time [38]. Techniques used previously for determining benthic virus abundance cannot discriminate between viruses arising from these different mechanisms.
There is limited evidence consistent with the hypothesis that viruses within sediments may originate from overlying waters. Recent study around the Saanich Peninsula (Canada) found cyanophage specific to Synechococcus sp. and phycoviruses specific to Heterosigma akashiwo deep (20–30 cm) within sediments [24, 40], suggesting they originated in the water column, then later became buried since neither of these organisms are benthic. Additionally, observation of sediment and pelagic virus abundance along several estuarine and trophic gradients [9, 19] suggests viriobenthos is correlated primarily to productivity of overlying waters, which also could be due to enriched sediments supporting greater autochthonous production. Recent study of the Brisbane River and Moreton Bay estuaries (Australia) found strong correlations between benthic virus abundance and the concentration of suspended solids >0.7 µm in the water column, suggesting that sorbed particles that settle to the benthos may contribute to high sediment abundances [19].
Virus production in the water column has been studied using several techniques. The approaches use virus decay [18], incorporation of radioactively labeled thymidine [39], the frequency of visually infected cells [34, 39, 45], fluorescently labelled virus disappearance [29], and dilution techniques [46], reviewed in [10]. The last of these techniques removes viruses from seawater and retains infected bacteria, which subsequently lyse during incubation, and a time course increase in virus abundance is then observed by microscopy. Virus production in sediments can also be studied using an adaptation of this dilution technique by diluting sediment samples with sterile sediments or 0.02-µm-filtered porewater, a method used in this current study. The rationale behind this method is to shift the normal balance between production and loss by reducing the loss processes (bacterial or enzymatic viral decay).
The aim of this study is to determine the potential rate of oxic surface sediment benthic virus production and the relative contribution of pelagic viruses to sediment virus communities. The potential origins of viruses in sediments was investigated at three stations off the coast of Southern California representing a range of productivities and relationships between sediments and overlying waters. Dilution techniques are presented which were used to determine the production of viruses within sediments, while virus particle sorptive scavenging by suspended sediment particles is investigated using artificially enhanced suspended sediments in seawater.
Methods
Study Site Description
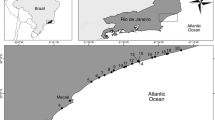
Three sites were chosen in the Southern California Bight region representing a range of water column production (Fig. l). The Los Angeles Harbor was sampled in November 2001 (autumn), the San Pedro Channel in June 2001 (summer) and the Big Fisherman’s Cove in February 2002 (winter). The Los Angeles harbor is a deep (>20 m) hypereutrophic artificially enclosed embayment, which has long water residence time, and several anthropogenic inputs from sewage treatment plants and urban/agricultural runoff via the channelised Los Angeles River, resulting in high productivity [23]. The specific location within the harbor (Cabrillo Beach; 33.7°N, 118.3°W) has a small area (~1 ha) of intertidal sand flats. In contrast, Big Fisherman’s Cove (Santa Catalina Island) is located within a restricted marine reserve and is meso- to oligotrophic (ambient water column NO −3 concentration ~1 µM). The sampling site was a subtidal sand flat located west of the Wrigley Marine Science Center dock (33.4°N, 118.4°W). The third site (33 33’N, 118 23’W), approximately halfway between Big Fisherman’s cove and Los Angeles Harbor is in approximately 900 m water in an occasionally suboxic basin [1]. The overlying water of the San Pedro Channel is oligotrophic and is subject to ephemeral advective upwelling [21].

Map of sampling locations in the Southern California region. (A) Los Angeles Harbor site, (B) San Pedro Channel site, and (C) Big Fisherman’s Cove site. Dotted lines indicate depth contours.
Sediment Core Collection
Sediments from Big Fisherman’s cove and Los Angeles Harbor were collected by wading out at low tide, and using a 5-mL syringe with the needle end removed as a sediment corer. Sediments for production experiments were placed initially into acid washed and seawater rinsed darkened polycarbonate bottles for transport (at ambient temperature) to the laboratory. Deep sediments from the San Pedro Basin were collected using a multicorer on board the R/V Point Sur (Moss Landing Marine Labs). After cores were retrieved, they were subsampled using syringe corers to desired depths.
Microscopy of Sediment Viruses
Viruses and bacteria in sediments were observed using SYBR Green I staining and epifluorescence miscroscopy as outlined in [19], which is a modification of water column virus staining [28]. Briefly, plugs of sediment (2 mL) were collected and discharged immediately into centrifuge tubes containing 8 mL of 0.02-µm-filtered 10 mM sodium pyrophosphate, 5 mM EDTA, and 2% formaldehyde in deionized water. Samples were then stored at 4°C until analysis (within 1 d). Before slide preparation, sediment samples were placed on a shaker table at 400 rpm for 10 min, then centrifuged at 1000 g for 5 min to remove fine sediments. Aliquots of eluate (typically 50–500 µL) were then filtered through 0.02 µm Al2O3 filters (Whatman Anodiscs). The filters were dried completely and stained with SYBR Green I (diluted 1:2500 from manufacturer supplied stock; Molecular Probes ; Eugene; OR). Filters were then dried completely again before mounting on a slide with 30 µL of antifade [0.1% p-phenylenediamine, 50% phosphate-buffered saline (120 mM NaCl, 10 mM NaH2PO4, pH 7.5], and 50% glycerol) as mountant. SYBR Gold and SYBR Green II were also tried for use in sediment virus microscopy, but their emission color is close to that of autofluorescent humic materials in sediments and they could not be used easily for sediment counts. Slides were examined on an Olympus BX-60 epifluorescence microscope under blue excitation at 100× objective. Viruses were distinguished from small sediment particles and colloidal mater by their color (humic materials autofluoresce orange-red, while viruses stained with SYBR Green I fluoresce green). More than 200 viruses and bacteria were counted for each slide, typically within 10 fields of view using an ocular grid. Virus abundances (M; cells cm−3) were then converted to volume of sediment using the equation

where mf is the count of microbes per field of view, θ is the number of fields per 25-mm Anodisc filter, S is the sediment volume (cm−3) extracted, E is the total eluate volume (cm−3); and F is the volume filtered (cm−3) for each slide.
Estimation of Virus Extraction Efficiency
To test the efficiency of virus and bacteria eluted in the previous section, sediment samples (15 mL) collected at the Big Fisherman’s Covesite in 50-mL sterile centrifuge tubes were shaken with 35 mL extraction buffer (10 mM pyrophosphate, 5 mM EDTA, 2% formaldehyde) for 10 min on a shaker table at 400 rpm. Afterwards the buffer was decanted completely into another 50-mL tube. A further 35 mL of extraction buffer was added to the sediment sample, which was reshaken for a further 10 min. This eluate was then decanted into another centrifuge tube. This process was repeated four times in total. All eluate samples were centrifuged at 1000 g for 5 min to remove large suspended material. The resulting supernatant was then processed for SYBR Green I epifluorescent microscopy as described above.
Virus Production
Experimental Design
The dilution method here is done in two different ways, targeting the two functional groups of bacteria in sediments, bacteria that are free within porewaters and unattached to sediment particles, and bacteria that are firmly attached to sediment grains [15]. Total potential virus production from all sediment bacteria is measured by diluting fresh sediments with sterilized sediment, then enumerating viruses produced over time in a homogenized slurry. Potential virus production by attached bacteria only is estimated by removing viruses and bacteria from porewater by filtration, then enumerating viruses produced (by the attached bacteria) over time in the filtered porewater after sediment rehydration with virus-free porewater.
Porewater Dilution (Attached Bacteria Potential Virus Production)
Porewater from sediments was collected by placing sediment samples (100 mL) over a GF/F filter (Whatman) in a 47-mm Gelman polycarbonate filtration manifold at room temperature and applying gentle vacuum (<5 mmHg) pressure until sediment was dry in consistency. Porewater content of sediments was determined by wet/dry comparison. Triplicate samples of sediment were first dried by decanting excess water, then rinsed over a 10-µm screen with deionized water to remove salts. Sediment samples were then dried at 60°C for 24 h before reweighing. The amount of porewater per unit volume of dry sediment varied (20–60%) between study sites. Porewater was then filtered through a 0.02-µm Anodisc filter (Whatman) to remove bacteria and viruses. Dry sediment was then reconstituted with filtered porewater (80–40% depending on predetermined wet/dry weight ratio) by gently introducing filtered porewater and agitation for ~2 min.
Sediment and Porewater Dilution (All Sediment Bacteria Potential Virus Production)
Approximately 500 g of sediment from each sampling location was washed with deionized water over a 10-µm Nitex screen, dried in a drying oven at 60°C, then autoclaved at 121°C for 30 min to kill live virus and bacteria. Virus-free porewater was prepared using the method described above and was used to rehydrate autoclaved sediments (~35% v/w of freshly collected and fully dried sediments was porewater). Rehydrated sterile sediment (80 mL) was then added to 20 mL freshly collected sediments and mixed thoroughly on a shaker table.
Triplicate incubations were prepared from each sampling site. Each 100-mL incubation was placed in a flat-bottomed acid-washed and seawater-rinsed polycarbonate container and incubated at ambient temperature (approx. 15°C) in the dark. Virus and bacterial abundances were measured from extracted (single wash) samples using SYBR Green I staining and epifluorescence microscopy as described earlier. Duplicate samples were withdrawn using a 3 mL cutoff syringe at regular intervals (approximately 1, 3, 6, 12, and 24 h). Virus production rate was calculated as the slope between mean virus abundances at each time point. Viruses produced by total bacteria (ν) was further calculated by multiplying slope by the dilution factor of hosts:

where dV is the slope of the regression of virus increase over time, b is the number of bacteria after dilution, and B is the initial undiluted number of bacteria.
Sorptive Scavenging
Collection of Sediments for Sorptive Scavenging Experiments
Sediments for sorptive scavenging experiments at all stations were collected from Big Fisherman’s Cove, Santa Catalina Island, from a sand patch immediately in front of the Wrigley Marine Science Center. Sediments were washed with deionized water over a 10-µm Nitex screen before being dried in a drying oven at 60°C overnight. Sediments were then size-fractioned using conventional dry sieve technique using standard sieves between 20 µm and 5 mm. Sediments were then autoclaved at 121°C for 1 h. Standard suspended matter solutions (10 g L−1) of fine sediments (i.e, <150 µm) were prepared in deionized water and were farther autoclaved for 30 min.
Effects of Sediment Size on Sorptive Scavenging
To determine the sorptive scavenging of different sized particles, sediments (ranging in size from 20 µm to 1 mm) were added to duplicate 50-mL seawater samples collected in 50-mL centrifuge tubes from the three sampling sites (0.02% sodium azide, final concentration, was added to prevent microbial growth) to a final concentration of 20 mg L−1. Duplicate seawater samples with suspended sediments and seawater samples with no addition were then shaken at 400 rpm at room temperature (20°C; water temperature 19°C) with tubes horizontal on a shaker table. After 24 h, incubations were fixed in 1% formaldehyde and centrifuged at 1000 g for 1 min; then the supernatant was processed for SYBR Green I staining and miscroscopy [28]. Sediment sizes assayed were 20, 75, 150, 250, and 500 µm.
Sediment Concentration Effects on Sorptive Scavenging
Aggregates and suspended particles in a similar environment to the San Pedro Channel have been studied previously using sediment traps moored near the sediment surface [37]. The mean size of sinking suspended solids in that study was found to be 1–63 µm in diameter, with a modal size of 26 µm. The 20-µm size fraction of sediments was therefore added to a range of final concentrations from 0–50 mg L−1. Incubations were then shaken for 24 h on a shaker table (at 400 rpm), before fixing with formaldehyde (2% final concentration) and epifluorescence microscopy as described above.
Determination of Suspended Sediment Concentration
Suspended sediment concentrations were determined by passing 2 L of site seawater through an anhydrous, preweighed GF/F filter (Whatman). Filters were rinsed with deionized water, then dried and reweighed. The difference in weight is the concentration of suspended solids >0.7 µm (nominal pore size of filters).
Results
Sediment Core Analysis
Viruses had maximum abundance near the sediment surface at the San Pedro Channel site, with decreasing abundance with sediment depth (Fig. 2). The abundance of bacteria and the ratio of viruses to bacteria follow virus distribution. Virus and bacteria abundance in sediments exceeded water column abundances on a per unit volume basis at all sites by between 1 and 2 orders of magnitude (Table 1). Viruses were lowest in the L.A. Harbor and similar at the other two locations, whereas bacteria were lowest in the San Pedro Channel and statistically indistinguishable at the other two locations. Note the very high VBR (98) at the surface of the Big Fisherman Cove site. Modal sediment grain size was highest the Big Fisherman’s Cove (250 µm), with the smallest in San Pedro Channel (10 µm).

Distribution of viruses and bacteria and the ratio of viruses to bacteria at the San Pedro Channel site. Error bars indicate SE (n = 4).
Microbial Extraction Efficiency
Of the microorganisms that eluted from the sediments by the third successive wash after the initial elution, approximately 85% of viruses on average were removed in the first elution, whereas only 51% of bacteria were removed using this method (Fig. 3). Fewer than 2% of viruses were obtained in the second and third successive washes; however, the second and third successive washes yielded 30% of the initial extracted bacterial abundance, suggesting that further bacteria could be shaken off in four or more washes of the sediment. Therefore, because one wash of sediments was used to estimate bacterial and viral abundances here, the bacterial counts are probably underestimates. Successive washes are recommended in future studies.

Extraction efficiency of sediment viruses and bacteria with successive washes of pyrophosphate. Error bars indicate SE (n = 4).
Virus Production
The abundance of virus-like particles increased linearly over the course of the experiment. Potential virus production rates in the Los Angeles Harbor and Big Fisherman’s Cove were approximately 2.8 × 108 and 5.0 × 108 VLP cm−3 h−1, respectively, for both attached bacteria and bacteria free within porewaters, and attached-only bacteria had rates of approximately 1.1 × 108 and 0.4 × 108 VLP cm−3 h−1 (Fig. 4). Virus turnover time in sediments was therefore 1 and 5 h for the Los Angeles Harbor and Big Fisherman’s Cove, respectively, where turnover is calculated by virus abundance/production rate. Virus abundances in initial samples had fewer viruses than subsequent time points, despite the delay in processing (24 h). Virus increase in incubations was therefore due to production by hosts, rather than passive desorption from particles. Killed undiluted samples from the Big Fisherman’s Cove site that have been observed over 8–30 d have shown no increase in abundance, suggesting that desorption of viruses upon preservation does not contribute to extractable virus increases, at least not in undiluted samples. Virus C release from sediment bacteria is 2.2–10.9 mg C dm−3 d−1 for the Los Angeles Harbor and Big Fisherman’s Cove, respectively, when calculated as bacterial C−100 virus C times bacterial lysis rate. This assumes a burst size of 100 and that viruses have a mean capsid diameter of 50 nm. The bacterial C/cell is 24 fg cell−1, used because this estimate is from larger cells in eutrophic Long Island Sound water (and bacteria in sediments are in a similar “eutrophic” environment), and a viral C content of 0.02 fg VLP−1 which is based upon bacterial carbon-to-volume ratios [25].

Combined virus production (porewater + sediment associated) and sediment-associated virus production, respectively, in the Los Angeles Harbor (A, B), and in Big Fisherman’s Cove site (C, D). Error bars indicate SE (n = 4).
Sorptive Scavenging
Viruses were removed by suspended particles of different sizes and by the 20-µm size fraction of different concentrations (Fig. 5). Approximately 30–60% of viruses in L.A. Harbor water adsorbed to particles <150 µm in diameter; however, particles larger than 150 µm adsorbed no viruses. A similar pattern was observed at the Big Fisherman’s Cove site, where between 20% and 40% of viruses in seawater adsorbed to sediment particles <150 µm; however, there was no viral sorption to larger sediments.

Virus removal (as percentage lost from incubation with no addition) by 20-µm sediments and range of sediments from 20 µm to 1200 µm, respectively, from the Los Angeles Harbor (A, B), in the San Pedro Channel (C), and at the Big Fisherman’s Cove (D, E). Error bars indicate SE (n = 4).
Sorption of viruses to sediment particles also was dependent upon the concentration of suspended sediments. In the L.A. Harbor, up to 90% of viruses in incubations were removed by adding 50 mg L−1 suspended sediment (20 µm size fraction). In Big Fisherman’s Cove, up to 50% of ambient viruses were removed by suspended sediment concentrations above 20 mg L−1. At the San Pedro Channel site, viruses did not adsorb to suspended sediment particles. The ambient concentration of suspended solids at the time of sampling was approximately 35 mg L−1, 2 mg L−1, and 9 mg L−1, at L.A. Harbor, San Pedro Channel, and Big Fisherman’s Cove sites, respectively. Light microscopic analysis of aliquots of suspended solid slurry revealed that the number of particles in 100 µL of the 10 g L−1, 20 µm size fraction slurry was 435 (±20), indicating that each particle weighed on average approximately 23 ng.
Discussion
The results presented here suggest that viruses in sediments may be produced rapidly and may account for a significant percentage of sediment bacterial mortality. Virus sorption to sediment particles (and subsequent settlement) may be an important contribution to sediment virus communities in areas with high suspended solids concentrations; however, in low suspended matter environments, such as in the oligotrophic Southern California Bight, this mechanism of virus arrival appears to be minor.
Viriobenthos Production
Virioplankton turn over approximately every 24–48 h [e.g., 13, 22, 45]. In this study the viriobenthos turned over on significantly smaller time scales (1–5 h). The observation of higher potential virus production rates in sediments is consistent with reports of higher bacterial production rates in sediments compared with overlying waters [44], as well as reports of higher abundance in sediments than in the water column in most sediment types [8]. Assuming an average burst size of 50–100 VLP per infected bacterium (which is based upon estimated burst size by electron microscopy in the water column; reviewed in [47]), viruses in sediments may cause lysis of 4–14% of bacterial abundance h−1. This may represent a significant loss of bacterial biomass considering benthic bacterial growth rates have been found to be within the range of 0.02 to 3.6 d−1 [5, 17]. Grazing rates of sediment bacteria by protozoa have been shown to only account for a small proportion (1.4–36% of total abundance) of daily bacterial production within sediments [43], suggesting that other loss mechanisms including viral lysis account for the majority of bacteria lost in this system. The calculated virus C release (2.2–10.9 mg C dm−3 d−1) suggests that this mechanism may be an important carbon transfer within sediments, but however is smaller than allochthonous inputs of POC from overlying waters (40–100 mg C dm−3 d−1 [35]). Virus-mediated mortality of sediment bacteria may also account for a greater percentage of sediment bacterial mortality than in the water column, which is potentially only 10–20% of bacterial biomass d−1 [41]. Since an artifact of this method is to underestimate bacterial abundance (shown in extraction efficiency tests), viral mortality of bacteria could be considerably smaller than indicated here.
Interestingly, this experiment suggested that the majority of viruses produced in sediments arise from infected bacteria free in the porewater (1.7 × 108 and 4.6 × 108 VLP cm−3 h−1; 61 and 92% of total benthic virus production, respectively; for L.A. Harbor and Santa Catalina Island, respectively), despite reports that most bacteria within sediments are firmly attached to particles [15]. This observation may be real or possibly an artifact of our experiments. Bacteria free in porewaters may be more productive than attached bacteria, and consequently may produce viruses more rapidly. Since attached bacteria live within biofilms on sediment surfaces, it is conceivable that the rate of virus infection is less due to sorption of infective viruses (loss) onto sediment surfaces. However, we may have missed some attached virus production because of an inability to enumerate viruses firmly attached to sediment biofilms. Also, undetermined inhibitory or stimulatory effects of drying, then rewetting sediments in the porewater dilution method may have resulted in an artifact of altered potential virus production from attached bacteria.
Errors Associated with Virus Production Estimates
The method presented here is only a potential virus production rate since the sediments are significantly disturbed by the method of the experiment. For example, several biogeochemical gradients are broken down as a consequence of sample collection which may stimulate bacterial production. Sediments were collected from the surface 2 mm of sediments; however, the oxic/anoxic boundary is typically only at ~1 mm. Additionally, sediments which are autoclaved may release substantial amounts of nutrients which also may have stimulated bacterial production. Another limitation of the methods presented here is that they assume that decay does not occur over the course of the experiment. Viral decay is considered to be potentially caused by protease and nuclease activity. Since decay rate is proportional to active cells and enzymes, one would expect an 80% drop in decay rate since enzymes are diluted 1:5 in this experiment.
Suspended particles in surface seawater are likely coated in organic and abiotic matter (as a consequence of bacterial activity), and therefore may behave differently from autoclaved sediment particles. Size-fractioned particles were first placed into deionized water to avoid instant sorption upon contact with water. In addition, sodium azide was used to terminate bacterial activity instead of formaldehyde or other preservatives since it was thought that the latter may cause cross-linking with proteins which may interfere with virus adsorption onto sediment particles, and preservation of microbes has been shown to reduce microbial populations after short periods of time [16].
The method of virus extraction presented here was shown to remove viruses from sediments (85% of total extractable population are removed in first rinse with pyrophosphate); however, significantly fewer bacteria were removed (51% of total extractable population in first rinse). Sediment particles observed after staining were devoid of virus particles but still had attached bacteria. Variability in grain size and obscured cells presented reliable enumeration on sediment particles. Since this study was conducted primarily to investigate sediment virus communities, this underestimation is unlikely to have affected virus production rates, although calculations involving bacterial abundance (e.g., percentages lysed per day) were affected. Considering that the ratios of firmly attached bacteria to free-living bacteria could be 200:1 to 600:1 [15] for freshwater detrital sediments, and given that only one-half of the extractable bacteria were enumerated in the first wash, the number of bacteria lysed per day could be significantly smaller than given here (a fraction of a percent per day). It is interesting to note that similar methods used elsewhere [7] have yielded lower virus recovery efficiencies (~50% recovery on first wash).
Virioplankton Sorption
Viruses at both the Big Fisherman’s Cove and L.A. Harbor sites adsorbed to particles collected in Big Fisherman’s Cove, whereas those at the San Pedro Channel site did not adsorb to these same particles. The ambient concentration of suspended solids appears correlated to the magnitude of potential viral sorption (i.e., L.A. Harbor > Big Fisherman’s Cove > San Pedro Channel), This is probably due to relative virus/particle contact rates, since the probability of viruses encountering particles by passive diffusion is greatest when virus abundance in ambient samples is highest and is also proportional to particle load. Another possible reason for this observation is that since viruses in oligotrophic low suspended matter areas may be produced more slowly than those in high suspended matter areas (e.g., [19]), they may be adapted against sorption to abiotic particles because the probability of subsequent infection by progeny viruses is lower when some are lost on sediment particles.
The observation of high rates of adsorption at the L.A. Harbor site initially suggests that arrival of viruses to sediments on particles may be important in contributing to sediment virus populations. However, combined with sedimentation rate data it is clear that this is still a very minor contribution to overall sediment populations. The sedimentation rate within shallow areas of the L.A. Harbor is approximately 35 mg cm−2 y−1 [14], which, based on the estimated weight of a single sediment particle (23 ng), is approximately 1.52 × 106 sediment particles arriving to sediments cm−2 y−1. Since the ambient concentration of suspended solids in the L.A. Harbor is approximately 30 mg L−1, approximately 60% of the virus standing stock is potentially removed by suspended sediment per day, which is approximately 400 VLP per sediment particle. This is consistent with decay rate measurements conducted in this region in previous studies [29]. Therefore, assuming that 400 VLP per particle is the maxmimum number of viruses that can adsorb to particles, the potential arrival of viruses on sediment particles is only approximately 7 × 104 VLP cm−2 h−1. The approximate turnover time by this mechanism alone is 74,500 h, which is very slow compared to potential autochthonous viriobenthos production in sediments. Since there were poor sorption rates of viruses to suspended particles at the San Pedro Channel site, it is not reasonable to expect this mechanism of virus arrival to be significant in this environment. It is interesting to note that previous observations of virus decay at the San Pedro Channel site using a fluorescently labeled virus approach have indicated a difference between production and decay (e.g., in July 1998, production was 5.1% d−1 and decay was 2.2% d−1) [27]. Virioplankton sorption to abiotic particles, which was not measured in this study, may account in part for this observation.
Grazing of Lytic Cells
Another possible mechanism by which viruses may arrive to sediments, which was not investigated in this study, is by sorption to particles and then uptake by filter-feeders and excretion, uptake by zooplankton of phytoplankton or bacteria in the final part of the lytic cycle, or adsorption of viruses to sinking detritus or cellular debris. Since viruses in the San Pedro Channel did not in our experiment appear to sorb significantly to sediment particles, it is likely that viral contribution to sediments via metazoan filter feeding is small. However, in areas of high suspended sediment concentrations, such as in estuaries, filter feeding organisms may increase sedimentation by up to 40% compared to areas away from infaunal benthic filter feeders [30]. Therefore, in estuaries this mechanism of virus arrival may be a significant input to sediments. Once viruses arrive in sediments via sorption mechanisms, they may become entrained and degraded by enzymes. There is limited evidence to suggest that virus-sized particles in the absence of interaction with labile enzymes do not become resuspended even after long periods of time [11].
Understanding of mechanisms by which viruses decay and are produced in sediments and the water column is essential to describing fully biogeochemical relationships between organisms in the ocean. The biogeochemical importance of viruses in the open ocean has been described previously [12, 42]; however, the high potential production rates of viruses in sediments measured here suggests that their role in this compartment may be larger than in overlying waters. In addition, wind-induced resuspension of sediments may bring large numbers of benthic viruses into the water column. This study raises interesting questions as to the dynamics of viruses in sinking aggregates and the specificity of viruses in sediments and how they might affect water column populations.
References
WM Berelson (1991) ArticleTitleThe flushing of 2 deep-sea basins, Southern California Borderland. Limnol Oceanogr 36 1150–1166 Occurrence Handle1:CAS:528:DyaK38XhtFejsbg%3D
DF Bird SK Juniper M RicciardiRigault P Martineu YT Prairie SE Calvert (2001) ArticleTitleSubsurface viruses and bacteria in Holocene/Late Pleistocene sediments of Saanich Inlet, BC: ODP Holes 1033B and 1034B, Leg 169S. Mar Geol 174 227–239 Occurrence Handle10.1016/S0025-3227(00)00152-3 Occurrence Handle1:CAS:528:DC%2BD3MXjtFegtrc%3D
TH Blackburn ND Blackburn (1993) ArticleTitleCoupling of cycles and global significance of sediment diagenesis. Mar Geol 113 101–110 Occurrence Handle10.1016/0025-3227(93)90152-L Occurrence Handle1:CAS:528:DyaK2cXhvFKhu70%3D
G Bratbak M Heldal TF Thingstad B Reimann OH Haslund (1992) ArticleTitleIncorporation of viruses into the budget of microbial C-transfer. A first approach. Mar Ecol Prog Ser 83 273–280
LM Cammen (1991) ArticleTitleAnnual bacterial production in relation to benthic microalgal produiction and sediment oxygen uptake in an intertidal sandflat and an intertidal mudflat. Mar Ecol Prog Ser 71 13–25
R Collier J Dymond S Honjo S Manganini R Francois R Dunbar (2000) ArticleTitleThe vertical flux of biogenic and lithogenic material in the Ross Sea: Moored sediment trap observations 1996–1998. Deep Sea Res Part II—Top Studies Oceanogr 47 3491–3520 Occurrence Handle10.1016/S0967-0645(00)00076-X Occurrence Handle1:CAS:528:DC%2BD3MXjsFOhtw%3D%3D
R Danovaro A DellÁnno A Trucco M Serresi S Vanucci (2001) ArticleTitleDetermination of virus abundance in marine sediments. Appl Environ Microbiol 67 1384–1387 Occurrence Handle10.1128/AEM.67.3.1384-1387.2001 Occurrence Handle1:CAS:528:DC%2BD3MXhslSjtrc%3D Occurrence Handle11229937
R Danovaro M Serresi (2000) ArticleTitleViral density and virus-to-bacterium ratio in deep-sea sediments of the eastern Mediterranean. App Environ Microbiol 66 1857–1861 Occurrence Handle10.1128/AEM.66.5.1857-1861.2000 Occurrence Handle1:CAS:528:DC%2BD3cXjtV2lt70%3D
LA Drake KH Choi AGE Haskell FC Dobbs (1998) ArticleTitleVertical profiles of virus-like particles and bacteria in the water column and sediments of Chesapeake Bay, USA. Aqua Microb Ecol 16 17–25
UR Fischer B Velimirov (2000) ArticleTitleHigh control of bacterial production by viruses in a eutrophic oxbow lake. Aquat Microb Ecol 27 1–12
JA Flood NJ Ashbolt (1999) ArticleTitleVirus-sized particles can be entrapped and concentrated one hundred fold within wetland biofilms. Adv Environ Res 3 403–411
JA Fuhrman (1999) ArticleTitleMarine viruses and their biogeochemical and ecological effects. Nature 399 541–548 Occurrence Handle1:CAS:528:DyaK1MXjvFeiurk%3D Occurrence Handle10376593
JA Fuhrman RT Noble (1995) ArticleTitleViruses and protists cause similar bacterial mortality in coastal seawater. Limnol Oceanogr 40 1236–1242
DS Gorsline (1992) ArticleTitleThe geological setting of Santa Monica and San Pedro Basins, California Continental Borderland. Prog Oceanogr 30 1–36 Occurrence Handle10.1016/0079-6611(92)90008-N
C Greibler B Mindl D Slezak (2001) ArticleTitleCombining DAPI and SYBR Green II for the enumeration of total bacterial numbers in aquatic sediments. Int Rev Hydrobiol 86 453–465 Occurrence Handle10.1002/1522-2632(200107)86:4/5<453::AID-IROH453>3.3.CO;2-C
K Gundersen G Bratbak M Heldal (1996) ArticleTitleFactors influencing the loss of bacteria in preserved seawater samples. Mar Ecol Prog Ser 137 305–310
JA Hansen DM Alongi (1991) ArticleTitleBacterial productivity and benthic standing stocks in a tropical coastal embayment. Mar Ecol Prog Ser 68 301–310
M Heldal G Bratbak (1991) ArticleTitleProduction and decay of viruses in aquatic environments. Mar Ecol Prog Ser 72 205–212
I Hewson JM O’Neil JA Fuhrman WC Dennison (2001) ArticleTitleVirus-like particle distribution and abundance in sediments and overlying waters along eutrophication gradients in two subtropical estuaries. Limnol Oceanogr 47 1734–1746
I Hewson JM O’Neil CA Heil G Bratbak WC Dennison (2001) ArticleTitleEffects of concentrated natural viral communities on photosynthesis and community composition of co-occuring benthic microalgae and phytoplankton. Aquat Microb Ecol 25 1–10
BM Hickey (1992) ArticleTitleCirculation over the Santa Monica San Pedro Basin and shelf. Prog Oceanogr 30 37–115 Occurrence Handle10.1016/0079-6611(92)90009-O
SC Jiang JH Paul (1994) ArticleTitleSeasonal and diel abundance of viruses and occurrence of lysogeny/bacteriocinogeny in the marine environment. Mar Ecol Prog Ser 104 163–172
GS Kieppel (1992) ArticleTitleEnvironmental regulation of feeding and egg production by Acartia tonsa off Southern California. Mar Biol 112 57–65
JE Lawrence AM Chan CA Suttle (2002) ArticleTitleViruses causing lysis of the toxic bloom-forming alga Heterosigma akashiwo (Raphidophyceae) are widespread in coastal sediments of British Columbia, Canada. Limnol Oceanogr 47 545–550
S Lee JA Fuhrman (1987) ArticleTitleRelationships between biovolume and biomass of naturally derived marine bacterioplankton. Appl Environ Microbiol 53 1298–1303 Occurrence Handle1:CAS:528:DyaL2sXksVGrtr4%3D
P Maranger DF Bird (1996) ArticleTitleHigh concentrations of viruses in the sediments of Lake Gilbert, Quebec. Microb Ecol 31 141–151
RT Noble JA Fuhrman (2000) ArticleTitleRapid virus production and removal measured with fluorescently labeled viruses as tracers. Appl Environ Microbiol 66 3790–3797
RT Noble JA Fuhrman (1998) ArticleTitleUse of SYBR Green I rapid epifluorescence counts of marine viruses and bacteria. Aquat Microb Ecol 14 113–118
RT Noble JA Fuhrman (1997) ArticleTitleVirus decay and its causes in coastal waters. Appl Environ Microbiol 63 77–83 Occurrence Handle1:CAS:528:DyaK2sXhs1Ojsw%3D%3D
A Norkko JE Hewitt SF Thrush GA Funnell (2001) ArticleTitleBenthic–pelagic coupling and suspension-feeding bivalves: linking site-specific sediment flux and biodeposition to benthic community structure. Limnol Oceanogr 46 2067–2072
JH Paul JB Rose SC Jiang CA Kellogg L Dickson (1993) ArticleTitleDistribution of viral abundance in the reef environment of Key Largo, Florida. Appl Environ Microbiol 59 718–724 Occurrence Handle1:STN:280:ByyB2cznt1M%3D Occurrence Handle8480998
LM Proctor JA Fuhrman (1992) ArticleTitleMortality of marine bacteria response to enrichments of the virus size fraction from seawater. Mar Ecol Prog Ser 87 283–293
LM Proctor JA Fuhrman (1990) ArticleTitleViral mortality of marine bacteria and cyanobacteria. Nature 343 60–61 Occurrence Handle10.1038/343060a0
LM Proctor A Okubo JA Fuhrman (1993) ArticleTitleCalibrating estimates of phage-induced mortality in marine bacteria: Ultrastructural studies of marine bacteriophage development from one-step growth experiments. Microb Ecol 25 161–182
EE Roden JH Tuttle WR Boyton WM Kemp (1995) ArticleTitleCarbon cycling in mesohaline Chesapeake Bay sediments. 1. POC deposition rates and mineralization pathways. J Mar Res 53 799–819 Occurrence Handle1:CAS:528:DyaK2MXps1Cjt7s%3D
K Simek J Pernthaler MG Weinbauer K Hornák JR Dolan J Nedoma M Masín R Amann (2001) ArticleTitleChanges in bacterial community composition and dynamics and viral mortality rates associated with enhanced flagellate grazing in a mesoeutrophic reservoir. Appl Environ Microbiol 67 2723–2733 Occurrence Handle10.1128/AEM.67.6.2723-2733.2001 Occurrence Handle1:CAS:528:DC%2BD3MXkt1Cit7c%3D Occurrence Handle11375187
RW Sternberg I Berhane AS Ogston (1999) ArticleTitleMeasurement of size and settling velocity of suspended aggregates on the northern California continental shelf. Mar Geol 154 43–53 Occurrence Handle10.1016/S0025-3227(98)00102-9
GF Steward DC Smith F Azam (1996) ArticleTitleAbundance and production of bacteria and viruses in the Bering and Chukchi Sea. Mar Ecol Prog Ser 131 287–300
GF Steward J Wikner WP Cochlan DC Smith F Azam (1992) ArticleTitleEstimation of virus production in the sea, II: Field results. Mar Microb Food Webs 6 79–90
CA Suttle (2000) Cyanophages and their role in the ecology of cyanobacteria, p. 563–589. M Potts (Eds) The Ecology of Cyanobacteria, vol. 1. Kluwer Academic Amsterdam
CA Suttle (1994) ArticleTitleThe significance of viruses to mortality in aquatic microbial communities. Microb Ecol 28 237–243
TF Thingstad M Heldal G Bratbak I Dundas (1993) ArticleTitleAre viruses important partners in pelagic food webs? Trends Evolutionary Ecol 8 209–213 Occurrence Handle10.1016/0169-5347(93)90101-T
FC vanDuyl RPM Bak AJ Kop G Nieuwland EM Berghuis A Kok (1992) ArticleTitleMesocosm experiments mimicking seasonal developments of microbial variables in North Sea sediments. Hydrobiologia 235 267–281
FC vanDuyl B deWinder AJ Kop U Wollenzein (1999) ArticleTitleTidal coupling between carbohydrate concentrations and bacterial activities in diatom-inhabited intertidal mudflats. Mar Ecol Prog Ser 191 19–32
MG Weinbauer P Peduzzi (1995) ArticleTitleSignificance of viruses versus heterotrophic nanoflagellates for controlling bacterial abundance in the northern Adriatic Sea. J Plankton Res 17 1851–1856
SW Wilhelm SM Bridgen CA Suttle (2002) ArticleTitleA dilution technique for the direct measurement of viral production: A comparison in stratified and tidally mixed coastal waters. Microb Ecol 43 168–173 Occurrence Handle10.1007/s00248-001-1021-9 Occurrence Handle1:CAS:528:DC%2BD38XjslGht74%3D Occurrence Handle11984638
KE Wommack RR Colwell (2000) ArticleTitleVirioplankton: Viruses in aquatic ecosystems. Microbiol Mol Biol Rev 64 69–114 Occurrence Handle10.1128/MMBR.64.1.69-114.2000 Occurrence Handle1:STN:280:DC%2BD3c7ntF2jsw%3D%3D Occurrence Handle10704475
Acknowledgements
The authors thank the crew of the R/V Point Sur, the staff of the Wrigley Marine Science Center on Santa Catalina Island, D. Hammond, W. Berelson, D. Capone, D. Kiefer, D. Caron, M. Schwalbach, J. Steele, and X. Liang for their assistance and input. I.H. was supported by a USC Seagrant Traineeship while conducting this research. This work was supported by NSF Grant OCE9906989 awarded to J.F.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Hewson, I., Fuhrman, J. Viriobenthos Production and Virioplankton Sorptive Scavenging by Suspended Sediment Particles in Coastal and Pelagic Waters . Microb Ecol 46, 337–347 (2003). https://doi.org/10.1007/s00248-002-1041-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00248-002-1041-0