
Abstract
G protein-coupled receptors are at a central node of all cell communications. Investigating their molecular functioning is therefore crucial for both academic purposes and drug design. However, getting the receptors as isolated, stable and purified proteins for such studies still stumbles over their instability out of the membrane environment. Different membrane-mimicking environments have been developed so far to increase the stability of purified receptors. Among them are amphipols. These polymers not only preserve the native fold of receptors purified from membrane fractions but they also allow specific applications such as folding receptors purified from inclusion bodies back to their native state. Of importance, amphipol-trapped G protein-coupled receptors essentially maintain their pharmacological properties so that they are perfectly adapted to further investigate the molecular mechanisms underlying signaling processes. We review here how amphipols have been used to refold and stabilize detergent-solubilized purified receptors and what are the main subsequent molecular pharmacology analyses that were performed using this strategy.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
G protein-coupled membrane receptors (GPCRs) are major players in all cell communication processes (Lagerstrom and Schioth 2008). As such, they are important targets for drug-design (Lagerstrom and Schioth 2008). During the last 10 years, our understanding of GPCR-mediated signaling has evolved from a simplistic picture where a receptor activates a single G protein subtype to trigger its biological effect to a more sophisticated picture where a single receptor can activate multiple G-protein dependent and independent pathways to trigger a full range of biological responses (Luttrell and Kenakin 2011). A current model associates this pleiotropic signaling to the ability of GPCRs to explore complex conformational landscapes where discrete conformations would have particular properties at interacting with different intracellular signaling proteins (Kenakin 2011; Planchard et al. 2014). Understanding the way the signal is transmitted through these receptors as well as designing new generations of drugs with better specificity and reduced side effects therefore requires a detailed description of these conformational landscapes. Although spectacular progresses have been made in the structural analyses of GPCRs, a detailed description of the whole conformational landscape a receptor can adopt and how it is influenced by pharmacological distinct ligands, signaling proteins and other processes such as dimerization is still hampered by the difficulties in stabilizing these integral membrane proteins in their native conformation out of the membrane environment for biophysical analyses.
While currently used to stabilize the native fold of purified integral membrane proteins, detergents tend to be inactivating for these proteins. The reasons for such inactivating properties are not detailed here but they include the loss of the physical constraints provided by the membrane, the competition with the protein–protein interactions that stabilize the native fold of the protein, and trapping of essential cofactors such as specific lipids (Popot 2010). This prompted the development of different approaches aimed at stabilizing GPCRs in solution. One of them consists in increasing the stability of the receptor in detergents through multiple mutations (Schlinkmann and Pluckthun 2013; Tate and Schertler 2009). However, it can never be excluded that, to some extent, such modifications in the receptor sequence introduce some biases in the structural dynamics of GPCRs. An alternative therefore consists in using the wildtype protein trapped in original detergents/surfactants that have increased propensity in stabilizing the native fold of the purified receptor. The maltose-neopentyl glucose detergent series (MNGs), amphipols, and nanodisks are among the most extensively used compounds. The respective pros and cons of these membrane-mimicking environments have been already reviewed (Popot 2010). We will focus here on amphipols only.
As detailed elsewhere (see Popot in this issue), amphipols are amphipatic polymers with a high density of hydrophobic chains and highly hydrophilic groups that have been designed to keep individual integral membrane proteins soluble under the form of stable amphipol:protein complexes. Different classes of amphipols have been developed so far that differ in their physicochemical properties (see Zoonens and Popot in this issue). Because of their ability to maintain the native fold of integral membrane proteins in solution better than detergents, amphipols have also been used in the GPCR field to stabilize these receptors out of the membrane. The availability of stable purified receptors trapped in amphipols allowed a series of pharmacological and structural studies to be carried out with the aim of delineating the molecular mechanisms underlying GPCR-mediated signaling.
Amphipols as a Way to Stabilize Purified GPCRs
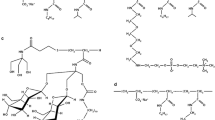
As stated above, a general trend of amphipols is that they stabilize integral membrane proteins better than any detergent. This prompted their use in the GPCR field to stabilize purified receptors in their native fold. In this context, two specific applications have been described so far that consist in either using amphipols to stabilize GPCRs after membrane solubilization with detergents or to fold the receptors back to their native state after purification under denaturing conditions (Fig. 1). Although many different amphipols have been developed so far (see Zoonens and Popot in this issue), we will consider only two of them here, the initially developed, negatively charged amphipol A8-35 and its non-ionic counterpart NaPol.

Strategies for trapping GPCRs in amphipols. Amphipol-trapped purified GPCRs can be obtained either after functional expression of the receptor at the membrane, detergent-mediated solubilization and detergent-to-amphipol exchange (a) or directly after functional refolding in amphipols of proteins recovered under denaturing conditions from bacterial inclusion bodies (b)
Stabilization of Detergent-Solubilized GPCRs
The last years have witnessed an explosion of the expression of GPCRs in eukaryotic systems such as insect sf9 and mammal cells for structural purposes (Andrell and Tate 2013). In these systems, the receptor is expressed in its native state at the plasma membrane. To get the receptor in its purified state, membranes have to be solubilized after cell lysis, usually with detergents. However, as is the case for many different membrane proteins, most of the purified GPCRs are unstable in such detergent environment (Le Bon et al. 2014). Based on their ability to stabilize integral membrane proteins, in particular α-helical proteins such as bacteriorhodopsin, amphipols have thus been used to stabilize purified GPCRs. The most documented case is that of the vasopressin V2 receptor (V2R). In this case, the receptor was expressed in sf9 cells and membranes solubilized with detergents (Rahmeh et al. 2012). Of importance, screening of multiple detergent mixtures was required to identify the complex mixture (Fos-Choline-12 or Fos-Choline-14/sodium cholate/DDM/cholesteryl hemisuccinate) that preserved the ligand-binding properties of the receptor. This detergent mixture was finally exchanged to the non-ionic amphipol NaPol using a biobeads-based classical protocol. Under such conditions, a stable receptor fraction was obtained that could be purified and subsequently used for pharmacological and structural characterizations (see below). Of importance, in accordance with the stabilization properties of amphipols, the vasopressin receptor in NaPols was stable over a period of time compatible with biophysical and pharmacological experiments. Since they significantly increased the V2R stability without affecting its pharmacological properties (see below), amphipols are therefore likely to be a promising alternative to detergents for receptor stabilization in strategies based on functional expression at the plasma membrane. Interestingly, trapping of the V2 receptor in NaPol allowed the separation of well-defined stable monomers and dimers. Dimerization is an important issue in the GPCR field. Although this process can be analyzed in other membrane-mimicking environments such as detergents (Arcemisbehere et al. 2010; Damian et al. 2006) or lipid disks (Mary et al. 2013), the data with the V2 vasopressin receptor indicate that amphipol trapping of GPCRs allows stabilization of complexes with well-defined stoichiometry that can be further used to compare the functional properties of these different assemblies.
Amphipols for GPCR Refolding
Expression of heterologous proteins in E. coli is frequently associated with accumulation of the recombinant protein in cytoplasmic aggregates, i.e., inclusion bodies (IBs). This occurs in most cases when GPCRs are produced in bacterial systems. Although commonly judged as a failure, getting GPCRs in IBs may present advantages to get high protein amounts (Baneres et al. 2011). This implies, however, that the receptors purified from IBs under denaturing conditions can be subsequently folded back to their native state in vitro. GPCR in vitro folding consists in exchanging the harsh detergent used for solubilizing the receptor from IBs with a milder surfactant (Baneres et al. 2011). Various environments that could stabilize the native fold of GPCRs in refolding experiments have been reported so far including detergents, lipid vesicles, and surfactants such as amphipols (Baneres et al. 2011). However, identifying the optimal conditions for refolding in detergents is time- and material-consuming since one has to screen for many different detergents and/or detergent mixtures to find the most appropriate one, whereas successful folding of GPCRs by transfer to lipid vesicles is limited to a very few examples [e.g., the chemokine receptor CXCR1 (Park et al. 2012a) or the NPY1 receptor (Schimmer et al. 2010)]. Moreover, even when refolding in detergent is successful, using the so obtained purified receptor stumbles again over the pitfall of detergent-associated instability.
Amphipols appear as an extremely promising alternative to detergents and lipids for refolding membrane proteins. This was first demonstrated using model proteins such as OmpA, FomA, and bacteriorhodopsin (Pocanschi et al. 2006; Dahmane et al. 2009, 2013; Pocanschi et al. 2013) and further extended to GPCRs recovered from IBs (Dahmane et al. 2009; Bazzacco et al. 2012). Of importance, conditions initially established to refold bacteriorhodopsin were applied essentially without any major change to several GPCRs to achieve folding yields between 30 and 50 % (Bazzacco et al. 2012; Dahmane et al. 2009, 2012), suggesting that this method of refolding could be of general use, in contrast to that based on detergents. Interestingly, addition of lipids significantly increased the refolding yields for most of the receptors considered (Dahmane et al. 2009). One possibility is that lipids increase the folding yields by binding to sites that form when the transmembrane surface achieves its native state. By doing so, they would contribute to drive the folding toward the native state of the receptor. Moreover, as observed for most APol-trapped membrane proteins, GPCRs folded in amphipols are significantly more stable than those kept in lipid/detergent mixtures while the pharmacological properties of the receptor are maintained (Dahmane et al. 2009). Finally, the receptor:amphipol complexes obtained are homogeneous from a molecular point of view [essentially monomeric receptor under the conditions used (Catoire et al. 2010)] and of moderate size [overall correlation times τc in the 50 ns range for the BLT2:A8-35 complex, i.e., ~30–50 % longer than that of membrane proteins in complex with commonly used detergents (Catoire et al. 2011)], two features that are highly favorable for many biophysical applications. Combined to the paramount interest in bacterial expression systems for specific applications such as NMR, as illustrated by the NMR-based structure of the unmodified chemokine receptor CXCR1 (Park et al. 2012b), all these data suggest that amphipols could represent a solution of choice to recover GPCRs from IBs, and this will represent an important breakthrough for in vitro studies of purified GPCRs.
Pharmacological Properties of Amphipol-Stabilized GPCRs
Besides classical studies such as crystallography, purified receptors have been widely used to investigate the molecular mechanisms underlying ligand binding, receptor activation, and coupling to signaling partners such as G proteins and arrestins. For such analyses, it is of course essential for the receptor to maintain its functional properties when purified. When it comes to GPCRs, maintaining the functional properties essentially implies that ligand recognition and coupling to signaling proteins such as G-proteins and arrestins are preserved.
Ligand Binding
The effects of assembling a detergent-solubilized receptor into amphipols on its functional properties have been investigated with several receptors. For instance, the V2R obtained after solubilization of membrane fractions in detergents and subsequent trapping into the non-ionic amphipol NaPol (see above) recognizes its AVP natural agonist as well as synthetic antagonists or biased agonists ligands with an efficacy rank of order closely related to that of the same receptor in its native environment (Rahmeh et al. 2012). This means that trapping V2R in NaPol does not primarily affect the recognition of ligands, whatever their pharmacological class is. This is also the case for GPCRs obtained after in vitro refolding in amphipols of receptors expressed in E. coli inclusion bodies. Indeed, the leukotriene B4 receptors BLT1 and BLT2, the serotonin receptor 5-HT4(a), the canabinoid receptor CB1, and the ghrelin receptor GHS-R1a refolded in either A8-35 (BLT1R, BLT2R, 5-HT4(a)R, CB1R) or NaPol (GHS-R1a; Fig. 2a) all bind their ligands with affinities in the same range than native receptors (Dahmane et al. 2009; Bazzacco et al. 2012). The fact that binding of ligands is observed whether the amphipol is ionic or non-ionic could mean that the charges in A8-35 do not primarily affect ligand recognition. However, caution needs to be exerted as A8-35 can give rise to high background non-specific binding when considering ligands that themselves bear a charge, as is the case for instance with the highly positively charged ghrelin peptide. All these data nevertheless indicate that assembling a GPCR into amphipols essentially preserves its ligand-binding properties, establishing that these polymers preserve the native fold of the receptor.

Pharmacological properties of the ghrelin receptor GHS-R1a folded in NaPols. a ligand-binding properties. Ligand binding was measured with a fluorescent analog of a ghrelin peptide as described in Damian et al. (2012). b Signaling protein activation. Activation of purified Gαqβγ was monitored as the ability of the receptor to trigger GTPγS binding using BODIPY-GTPγS fluorescence changes (Damian et al. 2012) whereas arrestin recruitment was monitored using a bimane-labeled purified arrestin-2 (Damian et al. 2012)
Signaling Protein Recruitment
Agonist-mediated activation of GPCRs results in their increased ability to recruit and activate their cognate signaling partners such as G proteins and arrestins. Maintaining the native fold of a purified receptor in solution therefore implies that its ability to interact with these cytoplasmic proteins is preserved. This is not always the case, depending on the surfactant used. For instance, in many cases, purified GPCRs in detergent solutions lose their ability to activate G proteins so that further reconstitution of purified receptors into lipid environments such as liposomes, bicelles or nanodisks is usually required (Park et al. 2012a; Kaya et al. 2011). In contrast, the V2 receptor trapped into the non-ionic amphipol NaPol after expression in sf9 cell membranes activated Gαs and recruited arrestin-2 in an agonist-dependent manner (Rahmeh et al. 2012). Of importance, the selectivity of the different ligands toward either recruitment of arrestins or G protein activation was preserved after reconstitution in amphipols, indicating that the selectivity in receptor activation was maintained under such conditions. In the same way, the ghrelin receptor obtained after refolding in NaPols was able to activate purified Gαq and interacts with arrestin-2 in a ghrelin-dependent manner (Fig. 2b). Of importance, in the case of the ghrelin receptor, not only the agonist-dependent Gq activation was preserved but also its high agonist-independent basal activity (Fig. 2b). This means that trapping the receptor into NaPols does not block the receptor into an inactive conformation but instead preserves some of the conformational respiration that is likely associated to constitutive activity. All these data are direct evidence that GPCRs in amphipols maintain their ability to interact with their associated signaling proteins in an agonist-dependent manner, a feature that is of paramount importance for further pharmacological studies. Moreover, since G protein activation and arrestin recruitment are primarily dependent on the activation of a GPCR by its agonist, the observations reported above indirectly point that trapping GPCRs into amphipols does not prevent the receptor to undergo the conformational changes associated with this activation process (Deupi et al. 2012) (see below).
Molecular Pharmacology of Amphipol-Trapped GPCRs
One of the major challenges actually in the GPCR molecular pharmacology field consists in getting a detailed description of the receptor dynamics in the context of the selection of a transduction pathway. Although major progresses have been made in cell-based studies, in many cases such analyses still rely on the use of purified proteins on which biophysical methods can be directly applied. Amphipol-trapped receptors are adapted to these studies due to the fact that the polymeric compound stabilizes the receptor in solution as a protein:amphipol complex of moderate, homogeneous, size while maintaining its functional properties. Moreover, this synthetic polymer does not contribute, in most of the cases to the signal and, in cases it does, it can be accordingly modified (e.g., deuterated amphipol for NMR studies).
Luminescence resonance energy transfer (LRET) technology was used with the vasopressin V2 receptor trapped in NaPol and labeled with fluorescence donor and acceptor at different positions to directly demonstrate that ligands with distinct pharmacological profiles (full agonists, biased agonists) stabilized distinct conformations of the receptor (Rahmeh et al. 2012). In the same way, distinct fluorescence resonance energy transfer (FRET) signals were observed with the ghrelin receptor refolded in NaPol and labeled with fluorescent probes in the presence of ligands from different classes (Fig. 3). This indicates that in this case also pharmacologically distinct ligands stabilized different conformations of the ghrelin receptor assembled in amphipols. Of importance, the changes in FRET efficiency were of comparable amplitude whether the receptor was trapped in NaPols or assembled into lipid disks (Fig. 3). This is an important observation as it means that amphipols, despites the fact that they do not directly mimic the highly complex and anisotropic lipid bilayer, nevertheless preserve the activation-associated conformational changes in a similar way than an environment mimicking more closely the membrane. In particular, amphipols do not preclude per se the conformational rearrangements required for GPCR functioning, even when large-scale rearrangements such as those associated with agonist-mediated activation are considered (Deupi et al. 2012). Associated to the fact that sample preparation of GPCRs associated to amphipols is straightforward compared to other solubilizing media such as bicelles or nanodisks, these observations make amphipol-trapped receptor well-adapted model systems for molecular pharmacology studies.

Ligand induced changes in the conformation of the ghrelin receptor. The ghrelin receptor was produced and assembled either in amphipols or in lipid disks as described (Damian et al. 2012). The receptor was then labeled with a unique fluorescence donor (Cy3) and acceptor (Cy5) at unique reactive cysteine and methionine residues located in the extracellular part of the first and fifth transmembrane helices, respectively. Labeling was carried out using the orthogonal strategy previously described (Lang et al. 2005). FRET efficiency was measured in the absence of ligands, in the presence of a full-agonist (MK0677), of a Gq-biased agonist (JMV3002), of a neutral antagonist (JMV3011) or of an inverse agonist (SPA) (Mary et al. 2012)
Amphipol-trapped GPCRs have not been used in fluorescence spectroscopy studies only. Indeed, their ability to stabilize in its native fold a receptor produced in E. coli opened the door to the use of more specific biophysical methods such as solution-state NMR. Application of NMR to amphipol-trapped receptors will not be detailed here since it is reported in an accompanying paper (Planchard et al. 2014). Briefly, stabilization of the purified LTB4 receptor BLT2 in A8-35 allowed the determination of the structure of two different agonists, LTB4 and 12-HHT, bound to the purified receptor (Catoire et al. 2010, 2011). Of importance, not only the NMR spectra of the receptor-bound ligands were obtained but also that of the perdeuterated, 15N uniformly labeled receptor (Catoire et al. 2010). Under such conditions, specific differences in the 2D 1H,15N CRINEPT spectrum of the receptor could be observed depending whether LTB4 was present or not (Catoire et al. 2010) indicating that, as reported above for the ghrelin receptor, agonist can indeed trigger changes in the conformation of the LTB4 receptor even when it is assembled in amphipols and that amphipol-trapped receptors can thus be readily used to analyze these changes with NMR-based approaches. All these results indicate that amphipol-trapped GPCRs are probably totally amenable to the use of solution-state NMR techniques to delineate the dynamics of receptors under a variety of conditions.
Conclusion
Analyzing the structural features of GPCRs as well as the relationship between their conformational dynamics and their signaling properties is still of paramount importance from both an academic perspective and for drug design purposes. Although spectacular progresses have been accomplished during the last years in crystallographic analyses, development of complementary biophysical methods is still required to get a detailed picture of the molecular determinants that govern GPCR signaling. Such biophysical studies still stumble over the instability of the receptor protein out of its membrane environment. In this context, associated to other membrane-mimicking media such as lipid nanodisks, amphipols offer unique perspectives. Indeed, these compounds have the potential to stabilize GPCRs in vitro after expression at the membrane of eukaryotic cells as well as to provide a unique medium in which receptors recovered from bacterial inclusion bodies can be refolded. The amphipol-trapped receptors essentially maintain their functional properties in terms of ligand binding and recruitment of signaling proteins. From a fundamental point of view, this means that, despite these polymers bear poor resemblance with a lipid environment, association with amphipols does not prevent neither the conformational rearrangement of the receptor associated with its activation nor the specific protein:ligand and protein:protein contacts occurring upon ligand recognition and coupling to signaling proteins. From a practical point of view, this means that purified GPCRs in amphipols can be used to further investigate the conformational features associated with signaling, as recently illustrated with the V2 vasopressin, leukotriene, and ghrelin receptors using fluorescence spectroscopy and solution-state NMR. Of importance, the synthetic versatility of amphipols provides us with a toolbox of compounds that can be accordingly modified for specific biophysical applications, e.g., deuterated amphipols for NMR studies (Giusti et al. 2014) or functionalized amphipols form immobilization (Della Pia et al. 2014). Trapping in amphipols therefore allows homogeneous preparations of stable receptor in particles of moderate size to be obtained. This opens the way to the application of many different biophysical methods, most of which are detailed in different articles of this special issue.
Combined to the recent progresses in expression systems for GPCRs, amphipols, in association with other stabilizing environments such as lipid disks or MNGs, have therefore the potential to open the way to integrated strategies associating up-to-date biophysical methods that should ultimately provide a detailed molecular description of how GPCRs can trigger their signal. The pro’s and con’s of these different membrane-mimicking media have been extensively discussed (Popot 2010) and will not be reviewed here. One can nevertheless mention that nanodisks have proved themselves useful when a strict control of the oligomeric state of the receptor is required (Mary et al. 2013) or when analyzing the impact of the lipid nature on the receptor function (Inagaki et al. 2012). On the other side, as developed in the present review, amphipols provide a way to refold and stabilize GPCRs using straightforward protocols. Amphipols are also particularly useful for instance when modification or functionalization of the stabilizing media is required for applications such as NMR and immobilization (Le Bon et al. 2014; Giusti et al. 2014). Finally, the possibility of trapping GPCRs in amphipols opens the way to more direct practical applications such as the immobilization of receptors on surfaces through functionalized amphipols for screening purposes (Della Pia et al. 2014 in preparation) or immunization with amphipol-trapped proteins. The latter application should have a general interest if we consider the increasing demand on GPCR-directed antibodies, in particular nanobodies, for both structural (Steyaert and Kobilka 2011) and signaling (Irannejad et al. 2013) studies.
Abbreviations
- 5HT4(a):
-
5-Hydroxytryptamine receptor 4
- 12-HHT:
-
12S-Hydroxyheptadeca-5Z, 8E,10E-trienoic acid
- A8-35:
-
Polyacrylate-based amphipol A8-35
- AVP:
-
Arginine-vasopressin
- BLT1:
-
Leukotriene B4 receptor 1
- BLT2:
-
Leukotriene B4 receptor 2
- CB1:
-
Cannabinoid receptor 1
- CRINEPT:
-
Cross-correlated relaxation-enhanced polarization transfer
- CXCR1:
-
C-X-C chemokine receptor 1
- DDM:
-
Dodecyl-β-D-maltopyranoside
- FRET:
-
Fluorescence resonance energy transfer
- FomA:
-
Outer membrane protein A from Fusobacterium nucleatum
- GHS-R1a:
-
Ghrelin receptor type 1
- GPCR:
-
G protein-coupled receptor
- IB:
-
Inclusion bodies
- LRET:
-
Luminescence resonance energy transfer
- LTB4 :
-
Leukotriene B4
- MNG:
-
Maltose-neopentylglucose
- OmpA:
-
Outer membrane protein A from Escherichia coli
- NaPol:
-
Non-ionic amphipol
- NMR:
-
Nuclear magnetic resonance
- Sf9 :
-
Spodoptera frugiperda
- V2R:
-
Vasopressin receptor 2
References
Andrell J, Tate CG (2013) Overexpression of membrane proteins in mammalian cells for structural studies. Mol Membr Biol 30(1):52–63
Arcemisbehere L, Sen T, Boudier L, Balestre MN, Gaibelet G, Detouillon E, Orcel H, Mendre C, Rahmeh R, Granier S, Vives C, Fieschi F, Damian M, Durroux T, Baneres JL, Mouillac B (2010) Leukotriene BLT2 receptor monomers activate the G(i2) GTP-binding protein more efficiently than dimers. J Biol Chem 285(9):6337–6347
Baneres JL, Popot JL, Mouillac B (2011) New advances in production and functional folding of G-protein-coupled receptors. Trends Biotechnol 29(7):314–322
Bazzacco P, Billon-Denis E, Sharma KS, Catoire LJ, Mary S, Le Bon C, Point E, Baneres JL, Durand G, Zito F, Pucci B, Popot JL (2012) Nonionic homopolymeric amphipols: application to membrane protein folding, cell-free synthesis, and solution nuclear magnetic resonance. Biochemistry 51(7):1416–1430
Catoire LJ, Damian M, Giusti F, Martin A, van Heijenoort C, Popot JL, Guittet E, Baneres JL (2010) Structure of a GPCR ligand in its receptor-bound state: leukotriene B4 adopts a highly constrained conformation when associated to human BLT2. J Am Chem Soc 132(26):9049–9057
Catoire LJ, Damian M, Baaden M, Guittet E, Baneres JL (2011) Electrostatically-driven fast association and perdeuteration allow detection of transferred cross-relaxation for G protein-coupled receptor ligands with equilibrium dissociation constants in the high-to-low nanomolar range. J Biomol NMR 50(3):191–195
Dahmane T, Damian M, Mary S, Popot JL, Baneres JL (2009) Amphipol-assisted in vitro folding of G protein-coupled receptors. Biochemistry 48(27):6516–6521
Dahmane T, Rappaport F, Popot JL (2013) Amphipol-assisted folding of bacteriorhodopsin in the presence or absence of lipids: functional consequences. Eur Biophys J 42(2–3):85–101
Damian M, Martin A, Mesnier D, Pin JP, Baneres JL (2006) Asymmetric conformational changes in a GPCR dimer controlled by G-proteins. EMBO J 25(24):5693–5702
Damian M, Marie J, Leyris JP, Fehrentz JA, Verdie P, Martinez J, Baneres JL, Mary S (2012) High constitutive activity is an intrinsic feature of ghrelin receptor protein: a study with a functional monomeric GHS-R1a receptor reconstituted in lipid discs. J Biol Chem 287(6):3630–3641
Della Pia EA, Holm JV, Lloret N, Le Bon C, Popot JL, Zoonens M, Nygard J, Martinez KL (2014) A step closer to membrane protein multiplexed nanoarrays using biotin-doped polypyrrole. ACS Nano 8(2):1844–1853
Deupi X, Standfuss J, Schertler G (2012) Conserved activation pathways in G-protein-coupled receptors. Biochem Soc Trans 40(2):383–388
Giusti F, Rieger J, Catoire LJ, Qian S, Calabrese AN, Watkinson TG, Casiraghi M, Radford SE, Ashcroft AE, Popot JL (2014) Synthesis, characterization and applications of a perdeuterated amphipol. J Membr Biol 36(7):1–6
Inagaki S, Ghirlando R, White JF, Gvozdenovic-Jeremic J, Northup JK, Grisshammer R (2012) Modulation of the interaction between neurotensin receptor NTS1 and Gq protein by lipid. J Mol Biol 417(1–2):95–111
Irannejad R, Tomshine JC, Tomshine JR, Chevalier M, Mahoney JP, Steyaert J, Rasmussen SG, Sunahara RK, El-Samad H, Huang B, von Zastrow M (2013) Conformational biosensors reveal GPCR signalling from endosomes. Nature 495(7442):534–538
Kaya AI, Thaker TM, Preininger AM, Iverson TM, Hamm HE (2011) Coupling efficiency of rhodopsin and transducin in bicelles. Biochemistry 50(15):3193–3203
Kenakin T (2011) Functional selectivity and biased receptor signaling. J Pharmacol Exp Therap 336(2):296–302
Lagerstrom MC, Schioth HB (2008) Structural diversity of G protein-coupled receptors and significance for drug discovery. Nat Rev Drug Discovery 7(4):339–357
Lang S, Spratt DE, Guillemette JG, Palmer M (2005) Dual-targeted labeling of proteins using cysteine and selenomethionine residues. Anal Biochem 342(2):271–279
Le Bon C, Popot JL, Giusti F (2014) Labeling and functionalizing amphipols for biological applications. J Memb Biol 288(37):26419–26429
Luttrell LM, Kenakin TP (2011) Refining efficacy: allosterism and bias in G protein-coupled receptor signaling. Methods Mol Biol 756:3–35
Mary S, Damian M, Louet M, Floquet N, Fehrentz JA, Marie J, Martinez J, Baneres JL (2012) Ligands and signaling proteins govern the conformational landscape explored by a G protein-coupled receptor. Proc Natl Acad Sci USA 109(21):8304–8309
Mary S, Fehrentz JA, Damian M, Gaibelet G, Orcel H, Verdie P, Mouillac B, Martinez J, Marie J, Baneres JL (2013) Heterodimerization with its splice variant blocks the ghrelin receptor 1a in a non-signaling conformation: a study with a purified heterodimer assembled into lipid discs. J Biol Chem 288(34):24656–24665
Park SH, Casagrande F, Chu M, Maier K, Kiefer H, Opella SJ (2012a) Optimization of purification and refolding of the human chemokine receptor CXCR1 improves the stability of proteoliposomes for structure determination. Biochim Biophys Acta 3:584–591
Park SH, Das BB, Casagrande F, Tian Y, Nothnagel HJ, Chu M, Kiefer H, Maier K, De Angelis AA, Marassi FM, Opella SJ (2012b) Structure of the chemokine receptor CXCR1 in phospholipid bilayers. Nature 491(7426):779–783
Planchard N, Point E, Dahmane T, Giusti F, Renault M, Le Bon C, Durand G, Milon A, Guittet E, Zoonens M, Popot JL, Catoire LJ (2014) The use of amphipols for solution NMR studies of membrane proteins: advantages and constraints as compared to other solubilizing media. J Memb Biol 63(9):11–18
Pocanschi CL, Dahmane T, Gohon Y, Rappaport F, Apell HJ, Kleinschmidt JH, Popot JL (2006) Amphipathic polymers: tools to fold integral membrane proteins to their active form. Biochemistry 45(47):13954–13961
Pocanschi CL, Popot JL, Kleinschmidt JH (2013) Folding and stability of outer membrane protein A (OmpA) from Escherichia coli in an amphipathic polymer, amphipol A8-35. Eur Biophy J 42(2–3):103–118
Popot JL (2010) Amphipols, nanodiscs, and fluorinated surfactants: three nonconventional approaches to studying membrane proteins in aqueous solutions. Annu Rev Biochem 79:737–775
Rahmeh R, Damian M, Cottet M, Orcel H, Mendre C, Durroux T, Sharma KS, Durand G, Pucci B, Trinquet E, Zwier JM, Deupi X, Bron P, Baneres JL, Mouillac B, Granier S (2012) Structural insights into biased G protein-coupled receptor signaling revealed by fluorescence spectroscopy. Proc Natl Acad Sci USA 109(17):6733–6738
Schimmer S, Lindner D, Schmidt P, Beck-Sickinger AG, Huster D, Rudolph R (2010) Functional characterization of the in vitro folded human y(1) receptor in lipid environment. Protein Pept Lett 17(5):605–609
Schlinkmann KM, Pluckthun A (2013) Directed evolution of G-protein-coupled receptors for high functional expression and detergent stability. Methods Enzymol 520:67–97
Steyaert J, Kobilka BK (2011) Nanobody stabilization of G protein-coupled receptor conformational states. Curr Opin Struct Biol 21(4):567–572
Tate CG, Schertler GF (2009) Engineering G protein-coupled receptors to facilitate their structure determination. Curr Opin Struct Biol 19(4):386–395
Acknowledgments
We are particularly indebted to J.-L. Popot (IBPC, Paris) for his invaluable contribution to all this work. We also wish to thank L. Catoire and M. Zoonens (IBPC, Paris) for helpful discussions. This work was supported by CNRS and National Agency of Research Grants ANR 06-BLAN-0087, ANR-10-BLAN-1208 and PCV08_323163.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Mary, S., Damian, M., Rahmeh, R. et al. Amphipols in G Protein-Coupled Receptor Pharmacology: What Are They Good For?. J Membrane Biol 247, 853–860 (2014). https://doi.org/10.1007/s00232-014-9665-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00232-014-9665-9