
Abstract
The mechanism of the intestinal creatine absorption is not well understood. Previous studies have established the involvement of a CT1 carrier system in jejunal apical membrane. The current research was aimed at completing the picture of creatine absorption. To investigate the process supporting creatine exit from enterocyte, basolateral membrane vesicles isolated from rat jejunum were used. The presence of various symport and antiport mechanisms was searched and a NaCl-dependent electrogenic transport system for creatine was evidenced, which shares some functional and kinetic features with the apical CT1. However, Western blot and immunohistochemical experiments ruled out the presence of a CT1 transporter in the basolateral membrane. Further studies are required to identify the basolateral transport mechanism. However, in the in vivo conditions, the NaCl gradient is inwardly directed, therefore such a mechanism cannot energetically mediate the exit of creatine from the cell into the blood during the absorptive process, but rather it may drive creatine into the enterocyte. To shed more light on the creatine absorption process, a possible creatine movement through the paracellular pathway has been examined using the jejunal tract everted and incubated in vitro. A linear relationship between creatine transport and concentration was apparent both in the mucosa-to-serosa and serosa-to-mucosa directions and the difference between the two slopes suggests that paracellular creatine movement by solvent drag may account for transintestinal creatine absorption. As a matter of fact, when transepithelial water flux is reduced by means of a mucosal hypertonic solution, the opposite creatine fluxes tend to overlap. The findings of the present study suggest that paracellular creatine movement by solvent drag may account for transintestinal creatine absorption.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
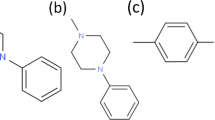
Creatine is an amino acid derivate supplemented with the diet as ergogenic and neuroprotective substrate. It is structurally similar to basic amino acids, has a molecular weight of 131 and a net positive charge at physiological pH (Fig. 1). Creatine is synthesized in liver, kidney and pancreas at rates of 1–2 g per day (Walker, 1979) and is transported via the blood to its target tissues, where it is taken up by the action of a highly specific Na+-Cl−-dependent transporter known as CT1 (Chen, Reith & Quick, 2004; Hediger et al., 2004). Intracellular creatine and phosphocreatine represent a high-energy buffering system that provides constant ATP supply (Walliman et al., 1992).

Creatine structure.
Dietary creatine supplementation in humans and animals results in an at least 3- to 20-fold increase in its serum concentration (Horn et al., 1998; Wyss & Kaddurrah-Daouk, 2000; Ipsiroglu et al., 2001); thus creatine is likely to be absorbed at the intestinal level. The mechanism responsible for creatine movement across intestinal apical membranes has been elucidated in both jejunal (Tosco et al., 2004) and ileal (Peral-Rubio et al., 2002) tracts, where a CT1 transporter has been evidenced. To put forward the knowledge on the access of creatine to the blood stream, we explored the contribution of basolateral membrane to the overall process of creatine absorption, using basolateral membranes isolated from rat jejunum. Since none of the mechanisms tested to account for creatine exit from the cell was apparent, the whole intestinal tract was used to search for a potential involvement of paracellular creatine permeation. Findings reported herein conclusively show that creatine undergoes paracellular absorption across jejunal epithelium.
Materials and Methods
BASOLATERAL MEMBRANE ISOLATION
Two male albino rats (Wistar strain, Charles River Italiana) weighing 250–300 g (about two months old), fed on a rodent laboratory chow and tap water, were used for each experiment. The rats were deeply anaesthetized with an injection of 2,2,2-tribromoethanol (0.05 g (250 g body weight)−1I.P.) and then decapitated. Jejunal enterocytes were collected by scraping off the mucosal layer and diluted in 250 mM sucrose, 1 mM dithiothreitol, 0.2 mM phenylmethylsulfonylfluoride (PMSF), 0.01% (v/v) ethanol, 20 mM HEPES/Tris buffer, pH 7.5 (sol. 1). After a 10 min centrifugation at 2,500 × g, the supernatant was collected and centrifuged again at 19,500 × g for 20 min (Kontron, Centrikon mod. T 2070 ultracentrifuge). A double pellet was formed: the fluffy upper layer was resuspended in sol. 1, homogenized, supplemented with 11% Percoll, homogenized again and centrifuged for 30 min at 48,000 × g. Then the third 8 ml from the bottom were withdrawn by a slow-rate peristaltic pump (Haake-Buchler, Auto Densi-Flow IIC apparatus), diluted with sol. 1 and centrifuged at 48,000 × g for 20 min. The pellet was suspended in the appropriate buffer (see single experiments) and 7 mM MgCl2, which aggregates preferentially all membranes except brush border, was added. The suspension was stirred occasionally in an ice bath for 5 min and in the following centrifugation (3,000 × g for 10 min) the supernatant was discarded and the pellet collected. The pellet was washed in the same buffer (45,000 × g for 20 min) and subsequently used for analysis and for [14C]creatine uptake experiments.
To control the purity of the basolateral membrane fraction, as a rule total protein, γ-glutamyltransferase (γ-GT), a marker enzyme for brush border membranes, and (Na+, K+)-ATPase, a marker enzyme for basolateral membranes, were determined. Proteins were determined by the Coomassie Blue binding assay (Bradford, 1976). γ-GT was measured according to Persijn and Van der Slik (1976) by using a commercial kit (Sentinel Diagnostics, Milano, Italy). The (Na+, K+)-ATPase activity was estimated by the slightly modified method of Schoner et al. (1967), in which the resynthesis of the ATP split by the ATPase is coupled to NADH oxidation via pyruvate kinase-lactate dehydrogenase reaction. Membranes were pretreated with a hypotonic solution in order to open sealed vesicles; in the same sample the kinetics in the absence (= total ATPase) and in the presence of ouabain (= ouabain-insensitive ATPase) were recorded. The difference in the slope between the two straight lines represents a measure of the (Na+,K+)-ATPase activity.
BRUSH BORDER MEMBRANE ISOLATION
Brush border membrane vesicles were isolated from rat jejunum enterocytes by the Mg2+ precipitation method first described by Schmitz et al. (1973). Mucosal scrapings, homogenized in hypotonic solution (50 mM sorbitol and 2 mM Tris Cl, pH 7.1) and incubated with 10 mM MgCl2 for 15 min at 0°C, were centrifuged at 3,000 × g for 15 min. The supernatant was centrifuged at 27,000 × g for 30 min. In order to obtain the desired intravesicular ion composition, the pellet was resuspended in the appropriate buffer and centrifuged at 43,000 × g for 20 min. The final pellet was resuspended in the above solution and subsequently used in the transport studies.
UPTAKE EXPERIMENTS
Transport of 8 μM [14C]creatine into vesicles was measured at 28°C by a rapid microfiltration technique. A volume of membrane suspension (2–4 mg protein/ml), equilibrated with 0.2 mM EDTA, was mixed with the proper incubation solution (zero time). The composition of the resuspension buffers and incubation media are given in the legends of the figures. Samples were removed at selected times and diluted with 0.8 ml of ice-cold reaction-stopping solution (20 mM HEPES/Tris buffer pH 7.5, 0.08 mM PMSF, 0.004% (v/v) ethanol and NaCl at the appropriate concentration for the required osmolarity), filtered on wetted cellulose nitrate filters (0.45 μm pore size) and immediately rinsed with 5 ml of the stop solution. The radioactivity of the filters was counted by liquid scintillation spectrometry (Tri-Carb, Packard, mod. 1600 TR).
All experiments were performed in voltage-clamp conditions by using either valinomycin and equal or different intra- and extravesicular K+ concentrations or equal intra- and extravesicular NO −3 concentrations. The values of the superimposed membrane potentials can be calculated by means of the Nernst equation. In this study, membrane potential was clamped either to 0 (equal intra- and extravesicular K+ or NO −3 concentrations) or to −59 mV, inside negative (10 times higher intravesicular K+ concentration). The solutions used were pre-filtered through 0.22 μm pore size filters. Individual uptake experiments in triplicate, representative of more than three repetitions with qualitatively identical results, are presented throughout the paper. Uptake values are presented as means ± SEM.
SDS-PAGE AND IMMUNOBLOTTING OF PROTEINS
Apical and basolateral membrane proteins from the jejunum of four rats were obtained as described above, except for the presence of protease inhibitors (300 μM phenylmethylsulfonyl fluoride (PMSF), 100 μM L-l-chloro-3-[4-tosylamido]-4-phenyl-2-butanone (TPCK), 1.5 μM pepstatin, 1.5 μM leupeptin, 10 μg/ml aprotinin) in the solutions. The final insoluble pellet was suspended in 250 μl of phosphate buffer saline containing protease inhibitors, 1% nonidet P40 (NP40) and 0.5 % Triton X-100 and solubilized. The protein concentration was measured according to the Bradford method (Bradford, 1976) and equal amounts of protein (60 μg) were analyzed by SDS-Page electrophoresis. Each sample was dissolved in Laemmli sample buffer [final concentration 2% (w/v) sodium dodecyl sulphate (SDS), 50% (v/v) glycerol, l% (v/v) 2-mercaptoethanol, 50 mM Tris, pH 6.8] and heated at 65°C for 10 minutes.
A 10% polyacrylamide minigel was run in a minigel apparatus (Hoefer SE 250-Amhersham) for 2 h at 120 V. Proteins were electrophoretically transferred to a polyvinylidene difluoride (PVDF) membrane (Miniprotean 3 - Biorad). After transfer, the membrane was stained with 0.1% Ponceau (Sigma, Italy) in 5% acetic acid to assess the quality of the transfer and equal loading of the lanes. After blocking with 5% non-fat dry milk in TBST buffer (50 mM Tris, 150 mM NaCl, 0.1% Tween, pH 8) for 2 h at room temperature, the proteins were probed overnight at 4°C with Anti N-terminal CT 75446 antibody (kindly supplied by Dr. Oliver Speer, Institute for Cell Biology, Zurich) diluted 1:1000 in 5% non-fat dry milk TBST buffer. The polyclonal antibodies against the N-terminal (NH2-MAKKSAENGIYSVSG-) and C-terminal (-PVSESSKVVVVESVM-COOH) of the rat and human CRT sequence (Nash et al. 1994) were prepared in rabbits as described by Guerrero-Ontiveros & Wallimann (1998). These sequences did not match the ones of any protein in the sequence data banks. The primary antibodies were detected with a donkey anti-rabbit IgG conjugated to horseradish peroxidase (Amersham N° NA 934) used at a 1:2000 dilution in 5% non-fat dry milk TBST buffer. Sites of antibody-antigen reaction were visualized by using Amersham ECL Plus, according to the manufacturer’s instruction, before exposure to X-ray film (Celbio, Italy).
IMMUNOHISTOCHEMISTRY
The jejunum was dissected from two adult rats under anesthesia, fixed by immersion in 4% paraformaldehyde in 0.1 M phosphate buffer (PB) pH 7.6 (5 h at 4°C), dehydrated in ethanol and embedded in paraffin. Sections (10 μm thick) were dewaxed in xylene, rehydrated by passing through a standard series of graded alcohols, and processed for the immunohistochemical detection of the CT1 transporter after antigen retrieval (microwave for 10 min in 0.01 M citrate buffer pH 6.0) and quenching of free aldehyde groups with 0.05 M NH4Cl in 0.01 M phosphate buffer saline (PBS) pH 7.4 (30 min). For the imnunoperoxidase procedure, sections were treated with 1% H2O2 (15 min) to inactivate endogenous peroxidase, preincubated in PBS containing 1% bovine serum albumin (BSA, Sigma) and 0.2% Triton X-100 (30 min), and then incubated with the anti N-terminal CT 75446 antibody diluted 1:500–1:1000 in PBS containing 0.1% BSA (48 h at 4°C in a humid chamber). After multiple rinses in PBS, sections were either a) incubated at room temperature in biotinylated goat anti-rabbit IgG (GAR, Vector) diluted 1:200 in PBS containing 0.1% BSA (75 min), rinsed in PBS and incubated in the avidin-biotin-peroxidase complex (ABC kit, Vector) diluted 1:100 in PBS, or b) incubated in the Envision kit (Dako). After rinsing in PBS and in 0.05 M Tris HC1 buffer (pH 7.6), sections were reacted in a fresh solution of 0.05% diaminobenzidine tetrahydrochloride (DAB, Sigma) and 0.002% H2O2 in Tris–HCl buffer and subsequently dehydrated and coverslipped. Sections were examined and photographed with a Zeiss III photomicroscope and collected by means of a high-resolution digital camera. For the immunofluorescence procedure, after the application of the primary antiserum, sections were incubated in GAR conjugated to Alexa 488 for 75 min and examined under a TCS NT confocal laser scanning microscope (Leica Lasertecknik GmbH, Heidelberg, Germany) equipped with a 75-mW Kripton/Argon mixed gas laser. Control slides were incubated in the same solutions without the primary antiserum and processed as above. This procedure always resulted in the absence of any staining.
EVERTED SAC PREPARATION
Under 2,2,2-tribromoethanol anaesthesia, rat upper jejunum (approximately 10 cm in length) was removed, everted, incubated and perfused for two subsequent periods of 20 min at 28°C. This temperature was chosen since at body temperature, at least in our hands, the transport activity of the in vitro intestine does not seem to be in a steady state (Faelli, Esposito & Capraro, 1976). The mucosal side of the intestine was incubated in 15 ml of Krebs-Henseleit-bicarbonate solution supplemented with 5.56 mM D-glucose and 8 mg/ml streptomycin and gassed with 95% O2 and 5% CO2. The serosal side of the intestine was first preincubated for 5 min with 0.5 ml of Krebs-Henseleit-bicarbonate solution supplemented with 20 mg% phenol red and after that, a first 20 min experimental period started. At the end, both serosal and mucosal compartments were thoroughly washed with the same solutions as above. After that, a further preincubation of 5 min and a second 20 min period of experiment were carried out. Either mucosal or serosal solution was supplemented with different concentrations of creatine and/or creatine transport inhibitors according to the experimental design. Samples of incubation fluids were withdrawn at the beginning and at the end of the two experimental periods, and assayed for phenol red (borax buffer pH 9.3, λ = 555 nm), creatine and creatinine (Sentinel Diagnostics, Milano, Italy. Creatinine was never detected.). At the end of the experiment, the intestine was blotted on filter paper and weighed. Transport of fluid into the serosal compartment (determined from the dilution of phenol red) was calculated as well as creatine transport towards mucosal and serosal sides. All transports were referred to wet weight of total intestine. Values reported in the text are means ± SE.
STATISTICS
Statistical analysis was done by Student’s t-test or by analysis of variance (ANOVA) followed by Newman-Keul’s Q-test.
Results
From the marker enzyme determinations, we calculated that the specific activities of (Na+, K+)-ATPase in the homogenate and in the basolateral membrane fraction were 138 ± 9 mU · mg protein−1 and 1904 ± 134 mU · mg protein−1, respectively (n = 8). The correspondent specific activities of γ-GT were 37 ± 8 and 21.5 ± 1 mU · mg protein−1 (n = 8). Thus, in the basolateral membrane fraction the (Na+, K+)-ATPase enrichment factor was 13.8 ± 1.8, while the enrichment of γ-GT was 0.58 ± 0.04.
To test the hypothesis that creatine transport across the basolateral membrane of jejunal enterocyte is mediated by antiport or symport mechanisms, studies were carried out using isolated membrane vesicles to check the effect of outwardly directed cation gradients and inwardly directed anion gradients on creatine uptake. Results reported in Figs. 2 and 3 show that the tested cations (H+, Na+, TMA+ and TEA+) and anions (HCO −3 , lactate and Cl−) are incapable of increasing creatine uptake value with respect to the control. These results exclude the possible involvement of members of various solute carrier families, e.g., Na+/H+ exchangers, cationic amino acid transporters, organic cation transporters, cation-Cl− cotransporters and bicarbonate transporters. It should be stressed that carbonic anhydrase activity is absent in rat jejunal basolateral membranes (Orsenigo, Tosco & Faelli, 1992) and thus the imposed HCO −3 gradient is adequate to test the possible coupling of HCO −3 to creatine transport. Moreover, data illustrated in Fig. 4 exclude the presence of symport mechanisms of creatine with Na+ and KCl, as well as the presence of a potential difference (PD)-sensitive conductive pathway.

Effect of outwardly directed H+, Na+, TMA+ and TEA+ gradients on creatine uptake. 25 μl membrane vesicles obtained in 90 mM mannitol and 50 mM MES/Tris buffer pH 5.5 (filled circles), or in 50 mM Na gluconate and 50 mM HEPES/Tris buffer pH 7.5 (triangles), or in 50 mM TMA-Cl and 50 mM HEPES/Tris buffer pH 7.5 (squares), or in 50 mM TEA-Cl and 50 mM HEPES/Tris buffer pH 7.5 (diamonds), or in 90 mM mannitol and 50 mM HEPES/Tris buffer pH 7.5 (empty circles) were incubated in 225 μl of 8 μM [14C] creatine, 90 mM mannitol and 50 mM HEPES/Tris buffer pH 7.5. All solutions contained 80 mM K gluconate, 0.2 mM PMSF and 0.01% (v/v) ethanol. Vesicles were preincubated with 25 μM valinomycin.

Effect of inwardly directed HCO −3 , lactate, Cl− gradients on creatine uptake. 25 μl membrane vesicles obtained in 90 mM mannitol were incubated in 225 μl of 50 mM NaHCO3 (filled circles), or 50 mM Na-lactate (triangles), or 50 mM TMA Cl (squares), or 90 mM mannitol (empty circles). All solutions contained 80 mM K gluconate, 50 mM HEPES/Tris buffer pH 7.5, 0.2 mM PMSF and 0.01% (v/v) ethanol. 8 μM [14C] creatine was in the incubating solutions. Vesicles were preincubated with 25 μM valinomycin.

Effects of Na+-gradient, KCl-gradient and PD on creatine uptake. 25 μl membrane vesicles obtained in 173 mM sorbitol and either 80 mM K gluconate (triangle, filled and empty circles) or 80 mM TMA NO3 (squares) were incubated in 225 μl of 80 mM K gluconate and 96 mM Na gluconate (triangles), or 80 mM TMA NO3 and 94 mM KCl (squares), or 317 mM mannitol (filled circles), or 80 mM K gluconate and 173 mM mannitol (empty circles). All solutions contained 50 mM HEPES/Tris buffer pH 7.5, 0.2 mM PMSF and 0.01% (v/v) ethanol. 8 μM [14C]creatine was in the incubating solutions. Except for the case represented by squares, vesicles were preincubated with 25 μM valinomycin.
Since evidence was given that the transport of organic cations across basolateral membranes can be linked to (Na+, K+)-pump activity (Laforenza, Gastaldi & Rindi, 1993), we measured creatine uptake in the presence of appropriately distributed ATP, Na+, K+ and Mg2+, in order to activate the (Na+, K+)-pump. The sidedness of the basolateral membrane preparation was previously determined (Orsenigo, Tosco & Faelli, 1994): the ratio of unsealed to sealed vesicles is 1:2; sealed vesicles are about 90% right-side-out and 10% inside-out oriented. These values have been confirmed in more recent studies. Since the site for catalytic hydrolysis of ATP is on the cytoplasmic side, results will be ascribed only to inside-out vesicles. As depicted in Fig. 5, the presence of ATP, Na+, K+ and Mg2+ strongly stimulates creatine uptake and the stimulation is enhanced by superimposing an inside-negative potential difference (PD); however, the addition of 5 mM strophanthidin (permeant analogue of ouabain) does not reverse the effect, making unlikely the actual involvement of the (Na+, K+)-pump in creatine uptake. Still in the presence of ATP, Mg2+ and PD, the removal of NaCl from the external medium reduces creatine uptake to the control level, thus excluding that a member of the ATP-binding cassette (ABC) transporter family could be responsible for creatine uptake stimulation.

Effect of (Na+, K+)-pump and ABC pump activation on creatine uptake. Vesicles were obtained in 80 mM K gluconate and 173 mM mannitol and an aliquot (squares) was preincubated with 5 mM strophanthidin for 20 min at room temperature. 25 μl membrane vesicles were incubated in 225 μl of 90 mM NaCl, 5 mM MgATP and 144 mM mannitol (filled circles, squares), or 90 mM NaCl, 5 mM MgATP and 80 mM K gluconate (triangles), or 5 mM MgATP and 310 mM mannitol (diamonds), or 80 mM K gluconate and 173 mM mannitol (empty circles). All solutions contained 50 mM HEPES/Tris buffer pH 7.5, 0.2 mM PMSF and 0.01% (v/v) ethanol. 8 μM [14C]creatine was in the incubating solutions. Vesicles were preincubated with 25 μM valinomycin.
The lack of strophanthidin inhibition shown in Fig. 5 induced us to better clarify the role of (Na+, K+)-pump in basolateral creatine movement. Data of Fig. 6 indicate that creatine uptake stimulation persists even if Mg2+ is removed from the incubation medium and ATP is substituted by its non-hydrolyzable analogue AMP-PNP. It has been reported that in some intestinal membrane vesicles the concentration of endogenous ATP can be as high as 90 μM (Van Dommelen, Hamer & DeJonge, 1986), however, in our experimental conditions 5.0 mM AMP-PNP should compete with possible endogenous ATP. Taken together, findings of Figs. 5 and 6 suggest that creatine uptake stimulation is not exerted by the (Na+, K+)-pump but rather by NaCl presence in the extravesicular solution.

Creatine uptake: effect of ATP and Mg2+ substitution in (Na+, K+)-pump–activating solutions. 25 μl membrane vesicles obtained in 80 mM K gluconate and 106 mM mannitol were incubated in 225 μl of 90 mM NaCl, 5 mM MgSO4, 5 mM Na2ATP and 66 mM mannitol (filled circles), or 90 mM NaCl, 5 mM MgSO4, 5 mM AMP-PNP and 60 mM mannitol (triangles), or 90 mM NaCl and 82 mM mannitol (squares), or 250 mM mannitol (empty circles). All solutions contained 50 mM HEPES/Tris buffer pH 7.5, 0.2 mM PMSF and 0.01% (v/v) ethanol. 8 μM [14C]creatine was in the incubating solutions. Vesicles were preincubated with 25 μM valinomycin.
To determine whether a carrier protein is involved in NaCl-dependent creatine transport, we examined the ability of 2 mM intravesicular creatine to transstimulate creatine uptake both in the presence and in the absence of NaCl in the incubating solutions. Figure 7 gives evidence that intravesicular creatine is able to energize the overshooting uptake of creatine only when NaCl is added, thus suggesting the presence of a carrier-mediated NaCl-dependent process. NaCl dependence is similar to that of the creatine transporter widely described in the literature (Chen, Reith & Quick, 2004; Hediger et al., 2004) and recently identified in jejunal brush border membranes (Tosco et al., 2004).

Transstimulation of creatine uptake by intravesicular creatine. 15 μl membrane vesicles obtained in 85 mM NaCl and 2 mM mannitol (filled circles), or 85 mM NaCl and 2 mM creatine (triangles), or 160 mM mannitol (empty circles), or 158 mM mannitol and 2 mM creatine (stars) were incubated in 285 μl of 85 mM NaCl and 2 mM mannitol (filled circles and triangles), or 160 mM mannitol (empty circles and stars). All solutions contained 80 mM K gluconate, 50 mM HEPES/Tris buffer pH 7.5, 0.2 mM PMSF and 0.01% (v/v) ethanol. 8 μM [14C]creatine + 100 μM unlabelled creatine was in the incubating solutions. Vesicles were preincubated with 25 μM valinomycin.
In order to identify the functional properties of the carrier mediating creatine transport across the basolateral membrane, we tested: 1) its cation and anion specificity (Fig. 8); 2) its kinetic parameters (Fig. 9); 3) its stoichiometry with respect to Na+ (Fig. 10) and Cl−(Fig. 11). Preliminarly, the uptake of both 10 μM and 1 mM creatine was determined as a function of short incubation times, resulting in a linear response for at least 45 s (data not shown). Thus, a 40 s uptake was considered as an estimate of the initial uptake rate and experiments described above were performed with this incubation time. Results obtained from all these studies match the ones reported for apical creatine transporter. The only difference concerns the basolateral Jmax value (Fig. 9), that turns out about 4 times smaller than the one calculated for apical transporter (120 ± 14 pmoles mg protein−1 40 s−1, n = 5 and 521 ± 8.25 pmoles mg protein−1 40 s−1, n = 5, respectively), thus suggesting either a lower expression of the carrier protein in the basolateral membranes or a different sealed-to-unsealed vesicles ratio between the two membrane preparations. Moreover, results of Fig. 10 suggest that, as in brush border, at least 2 Na+ ions are required to transport one creatine molecule in the presence of Cl−; however, the low value of the Hill coefficient is indicative of a smaller cooperativity in the basolateral membranes.

Cation and anion specificity of creatine uptake. 10 μl membrane vesicles obtained in 100 mM K gluconate were incubated for 30 s in 40 μl of a buffer containing 100 mM of one of the salts indicated. Panel A shows creatine uptake in various cationic buffers, panel B shows creatine uptake in various anionic buffers. All solutions contained 220 mM mannitol, 50 mM HEPES/Tris buffer pH 7.5, 0.2 mM PMSF and 0.01% (v/v) ethanol. 10 μM [14C] creatine was in the incubating solutions. Vesicles were preincubated with 25 μM valinomycin.

Initial rate (40 s) of creatine uptake versus increasing concentrations of extravesicular creatine. 10 μl membrane vesicles obtained in 100 mM K gluconate were incubated for 40 s in 40 μl of 100 mM NaCl and increasing creatine concentrations (5–300 μM). All solutions contained 220 mM mannitol, 50 mM HEPES/Tris buffer pH 7.5, 0.2 mM PMSF and 0.01% (v/v) ethanol. [14C] creatine was in the incubating solutions. Vesicles were preincubated with 25 μM valinomycin. The small binding of creatine on the vesicle surface was subtracted and the Michaelian hyperbolic relationship obtained was linearized by Eadie-Hofstee transformation of the data, showing a good correlation coefficient (r = 0.98, inset). Calculated parameters were: Km = 25 ± 2.8 μM and Jmax = 120 ± 14 pmoles mg protein−1 40 s−1.

Dependence of creatine uptake on Na+ concentration.10 μl membrane vesicles obtained in 100 mM K gluconate and 565 mM mannitol were incubated for 40 s in 40 μl of NaCl at different concentrations (0–270 mM), which were made by substitution of NaCl with TMA-Cl. All solutions contained 50 mM HEPES/Tris buffer pH 7.5, 0.2 mM PMSF and 0.01% (v/v) ethanol. 10 μM [14C] creatine was in the incubating solutions. Vesicles were preincubated with 25 μM valinomycin. Since the rate of creatine uptake increases sigmoidally when the external concentration of Na+ is varied, results were analyzed by using the Hill plot, which was found to be linear (r = 0.99, inset). The Hill coefficient for Na+, determined from the slope of the Hill plot, is 1.35±0.07. Approximate values of kinetic parameters are: Km = 153 mM and Jmax = 69 pmoles mg protein−1 40 s−1.

Dependence of creatine uptake on Cl− concentration. 10 μl membrane vesicles obtained in 100 mM K gluconate and 440 mM mannitol were incubated for 40 s in 40 μl of NaCl at different concentrations (0–270 mM), which were made by substitution of NaCl with Na gluconate. All solutions contained 50 mM HEPES/Tris buffer pH 7.5, 0.2 mM PMSF and 0.01% (v/v) ethanol. 10 μM [14C] creatine was in the incubating solutions. Vesicles were preincubated with 25 μM valinomycin. Results were linearized by Eadie-Hofstee transformation of the data (r = 0.99, inset), giving Km = 21.8 ± 2.2 mM and Jmax = 222.5 ± 19 pmoles mg protein−1 40 s−1.
In view of the functional similarities between apical and basolateral creatine transport systems, the possible presence of a CT1 transporter was searched using both Western blot analysis and immunohistochemical techniques. Western blot experiments were performed on both jejunal apical and basolateral membrane proteins. In the brush border the previously reported 70 and 55 kDa bands were present, as shown in Fig. 12, whilst in the basolateral membrane fraction no immunoreactivity was observed. The immunohistochemical analysis (Fig. 13) corroborates the immunoblot data since the reaction product observed with both immunoperoxidase (Fig. 13A,B) and immunofluorecence (Fig. 13C) procedures is always confined to the enterocyte apical membranes and not detected at their basolateral membranes. Protein expression is most prominent at the villus tip and declines towards the crypts. However, the antibody used in both Western blot and immunohistochemical analysis was raised against the N-terminal sequence of CT1. Since there are examples of splice mutants of transporters that are truncated at their N-terminus and consequently trafficked to a different epithelial cell membrane domain (Adair-Kirk, 1999), we repeated Western blot experiments using an antibody raised against the CT1 C-terminal sequence. Once again no evidence was provided for CT1 presence at the basolateral level (data not shown).

Western blot analysis of jejunal apical (Ap) and basolateral (Bl) proteins performed with an anti-CT antibody.

Immunohistochemical detection of CT1 transporter. Immunoperoxidase (A, B) and immunofluorescence (C) labeling of jejunal sections show the presence of specific reaction product at the apical membranes of enterocytes (arrows). Original magnifications: A = 940 ×; B = 4.200 ×; C = 3.700 ×
Thus we can rule out CT1 expression in jejunal basolateral membranes. We can exclude any contribution of brush border transport systems in the results of uptake experiments since the marker enzyme analysis shows that the basolateral membrane preparation is very well purified; moreover our basolateral membranes were tested for their ability to accumulate D-glucose in the presence of an inward Na+ gradient (Fig. 14). The lack of effect is in agreement with the absence of a significant brush border contamination.

Control of brush border contamination by D-glucose uptake. 15 μl basolateral membrane vesicles obtained in 100 mM K gluconate and 70 mM mannitol were incubated in 225 μl of 100 mM NaCl, 64 mM mannitol and either 10 μM [14C]creatine (filled circles) or 0.2 mM D-glucose and trace amounts of [3H]D-glucose (filled squares), or 100 mM gluconate, 70 mM mannitol and either 10 μM [14C]creatine (empty circles) or 0.2 mM D-glucose and trace amounts of [3H]D-glucose (empty squares). All solutions contained 50 mM HEPES/Tris buffer pH 7.5, 0.2 mM PMSF and 0.01% (v/v) ethanol. Vesicles were preincubated with 25 μM valinomycin.
Besides Na+ and Cl−, in all experiments carried out so far, K+ was present in the intravesicular medium. It is known that inwardly directed Na+ and Cl− gradients coupled with outwardly directed K+ gradients serve as driving force for the transport of biogenic amines such as serotonine (5-HT, Rudnick & Clark, 1993). Since immunocytochemical observations suggested that 5-HT transporter (SERT) is expressed by enterocytes throughout the crypt-villus axis (Chen et al., 1998), to test its possible role in our results, we performed competition experiments using 70 μM and 700 μM unlabelled creatine, or 5-HT, or dopamine. 70 μM and 700 μM unlabelled creatine inhibit creatine uptake by 77% ± 3 and 99% ± 0.6, respectively (5 determinations), while the same concentrations of 5-HT exert 7% ± 1 and 22% ± 8 inhibition, respectively (5 determinations). Values obtained in the presence of dopamine are 8% ± 7 and 25% ± 4 (5 determinations). Thus it seems unlikely that the activation of 5-HT transporter could account for basolateral creatine transport.
To sum up, jejunal basolateral membranes exhibit a NaCl-dependent creatine transporter different from both CT1 and SERT. However, in the in vivo conditions, NaCl gradient is inwardly directed, thus it would drive creatine into the cell from the serosal side; therefore, a basolateral NaCl-dependent transporter could not account for creatine absorption towards the blood but rather for creatine entry into the enterocyte. No evidence is provided in this study for the presence of other creatine transport mechanisms in the same membranes. However, literature data suggest that creatine is absorbed at the intestinal level (Horn et al., 1998; Wyss & Kaddurrah-Daouk, 2000; Ipsiroglu et al., 2001). It is generally assumed for hydrophilic solutes, which cannot readily cross the cellular lipid membrane, that passive diffusion via the paracellular route would be the principal transport mechanism for intestinal absorption unless carrier-mediated transport systems are available. Thus, a possible creatine movement through the paracellular pathway has been examined using the jejunal tract everted and incubated in vitro.
Creatine transport from the mucosal to the serosal compartment was evaluated during 20 min incubation at 28°C in the presence of increasing creatine concentrations in the mucosal solution. Data reported in Fig. 15 show a linear relationship between transport and concentration (slope = 420 ± 31 μl g−1 20 min−1, r = 0.99); in the presence of 0.2 mM creatine, the addition of 1 or 2 mM β-guanidinopropionate (a structural analogue of creatine) or 0.5 mM 2-aminoethyl methanethiosulfonate hydrobromide (MTSEA, a methanethiosulfonate reagent) does not affect creatine transport (data not shown). Both substances are known to inhibit the apical transporter (Tosco et al., 2004). Thus, a passive paracellular pathway is likely to be involved in creatine movement. By reversing the experimental condition, i.e., adding creatine into the serosal compartment, a linear serosa-to-mucosa creatine movement is apparent (Fig. 15), but this transfer is significantly smaller than the mucosa-to-serosa one (slope = 228 ± 33 μl g−1 20 min−1, r = 0.94). The difference could be due to the transepithelial water flow, that is always present in our experimental conditions (0.58 ± 0.04 μl g−1 20 min−1, 13 determinations) and is known to affect the paracellular transport of drugs. If there is water absorption, paracellular mucosa-to-serosa permeation could be enhanced due to an additional solute flux associated with water flux (solvent drag effect). On the other hand, water absorption could reduce serosa-to-mucosa paracellular permeation by hindering diffusive flux. To test this hypothesis, water flux was reduced by means of a mucosal hypertonic solution (0.33 ± 0.03 μl g−1 20 min−1, 13 determinations); as a matter of fact, results reported in Fig. 15 indicate that under this experimental condition the opposite creatine fluxes tend to overlap (mucosa-to-serosa slope: 220 ± 33 μl g−1 20 min−1, r = 0.97; serosa-to-mucosa slope: 183 ± 30 μl g−1 20 min−1, r = 0.96).

Bi-directional fluxes of creatine at 28°C in everted sac preparations. Creatine flux was examined in the mucosa to serosa (m-s) and serosa to mucosa (s-m) directions. 20 min incubation experiments were performed, with creatine concentrations ranging from 0.5 to 15 mM. Each value represents the mean ± SEM of 10 experiments. Left: control condition. Right: 400 mM mannitol added in the mucosal solution.
Discussion
The aim of this work was to characterize the mechanism involved in the uptake of creatine by rat jejunal basolateral membranes, since up to now nothing is known concerning the basolateral step of creatine absorption.
To focus on the mechanism that would support creatine exit from enterocyte, isolated basolateral membranes were used. Actually, for a substrate that undergoes some degree of intracellular metabolism, as is the case with creatine, such studies are best performed using purified membrane vesicle preparations, which are devoid of intracellular components.
There are two principal routes by which intracellular creatine may cross the basolateral membrane: it may undergo passive diffusion or may be facilitated via specific carrier-mediated pathways. However, its rather high molecular weight (131.13 Da) and its positive charge at physiological pH make the former hypothesis unlikely. A plethora of transmembrane transport proteins is present in the intestine; whereas most are highly specialized, mediating facilitated transport of essential nutrients, some others are polyspecific transporters due to their broad substrate specificity. Data reported in Fig. 1 exclude the presence of an exchange mechanism of creatine for Na+, H+, TMA+ and TEA+ in the basolateral membrane, thus ruling out the involvement of antiports that might be mediated by various transporters, including organic cation transporters (OCT). The OCT belong to a superfamily that includes multidrug-resistance proteins, proton antiporters and facilitative diffusion systems that mediate the transport of a variety of structurally diverse organic cations. OCTs are found in intestinal serosal and brush border membranes (Wright & Dantzler, 2003). A possible role of OCT in creatine transport is ruled out also by results depicted in Fig. 4 (filled and empty circles), that exclude the presence of an electrogenic uniport of creatine, as well as the presence of a conductive pathway. Alternatively, creatine transport across the basolateral membrane might be coupled with the flux of anions that exit the cell down their electrochemical gradient, namely, HCO −3 (which is absorbed in the jejunal tract of the intestine; Parsons, 1956), lactate (which is produced by the glycolytic metabolism of jejunal enterocyte; Cremin & Fleming, 1997) and Cl− (which is introduced into the cell by a basolateral Cl−/HCO −3 exchanger; Orsenigo, Tosco & Faelli, 1991). However, as illustrated in Fig. 3, also this hypothesis can be excluded.
A possible involvement of primary-active, ATP-dependent transporters, including (Na+, K+)-pump and members of the ATP-binding cassette (ABC) transporter family, in creatine transfer across the basolateral membrane is ruled out by data of Figs. 4 and 5. On the contrary, the data strongly evidence that creatine uptake is stimulated by the presence of NaCl in the incubation medium and is further increased by a superimposed membrane potential. The overshoot phenomenon suggests the presence of an electrogenic NaCl-dependent carrier-mediated system, whose existence is further confirmed by the finding of a significant transstimulation by unlabelled creatine (Fig. 7). This observation is somewhat puzzling, since such a mechanism could energetically mediate the exit of creatine from the cell into the blood during the absorptive process only if the thermodynamics of apical and basolateral transporters were different so that a sufficiently high intracellular creatine concentration could be generated for basolateral efflux. Kinetic results reported in this study, as well as the electrogenicity of basolateral transport, indicate that this is not the case, but rather that the basolateral transporter may drive creatine into the enterocyte.
Thus, results obtained in the present research and in a previous one (Tosco et al., 2004) suggest that jejunal enterocytes can take in creatine from both luminal and serosal sides. We can assume that the mechanisms supporting apical and basolateral creatine intake may supply creatine for enterocyte motility. Actually, high levels of creatine are found in the intestine (Wyss & Kaddurrah-Daouk, 2000), where cytosolic creatine kinase isoenzymes have been observed to associate with the cellular cytoskeleton and have been localized distinctly in the terminal web region of the brush border (Walliman & Hemmer, 1994). Further studies are required to verify this hypothesis and to identify the basolateral NaCl-dependent transport mechanism for creatine, which does not correspond to CT1, as shown in the present study.
Besides that, creatine is absorbed in small intestine. As a matter of fact, dietary creatine supplementation in humans and animals results in at least 3- to 20-fold increase in the serum concentration of creatine (Wyss & Kaddurrah-Daouk, 2000). In particular, it has been demonstrated that creatine feeding raises serum creatine levels in rat (Horn et al., 1998, Ipsiroglu et al., 2001). Results of the in vitro experiments suggest that creatine absorption occurs via a paracellular pathway: actually, creatine transepithelial transport is unaffected when inhibitors of apical creatine transporter are added. Moreover, previous studies gave no evidence for a diffusive creatine permeation across the apical membrane (Tosco et al., 2004). Intestinal absorption of small paracellular marker molecules has been shown to increase in direct proportion to the net water absorption, suggesting that transepithelial movement could be mediated by solvent drag in the fluid absorbed through the paracellular pathway. Results of Fig. 15 indicate that paracellular creatine movement by solvent drag may account for transintestinal creatine absorption. Paracellular movement has been suggested to be an important component for intestinal absorption of many hydrophilic drugs and peptides (Karlsson et al., 1999): this study demonstrates that this pathway is shared also by creatine.
References
Adair-Kirk T.L. 1999. Intracellular trafficking of variant chicken kidney AE1 exchangers: role of alternative NH(2) termini in polarized sorting and Golgi recycling. J.Cell Biol. 147:1237–1248
Bradford M.N. 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of proten-dye-binding. Anal. Biochem. 72:248–254
Chen J.X., Pan H., Rothman T.P., Wade P.R., Gershon M.D. 1998. Guinea-pig 5-HT transporter: cloning, expression, distribution and function in intestinal sensory reception. Am. J. Physiol, 275:G433–G448
Chen N.H., Reith M.E., Quick M.W. 2004. Synaptic uptake and beyond: the sodium- and chloride-dependent neurotransmitter transporter family SLC6. Pfluegers Arch. 447:519–531
Cremin J.D., Fleming S.E. 1997. Glycolysis is a source of pyruvate for transamination of glutamine amino nitrogen in jejunal epithelial cells. Am. J. Physiol. 272:G575–G588
Faelli A., Esposito G., Capraro V. 1976. Energy-rich phosphates and transintestinal transport in rat intestine incubated in vitro at different temperatures. Biochim-BiophyActa 455:759–766
Guerrero-Ontiveros M.L.,Wallimann T. 1998. Creatine supplementation in health and disease. Effects of chronic creatine ingestion in vivo: Down-regulation of the expression of creatine transporter isoforms in skeletal muscle. Mol. Cell. Biochem. 184:427–437
Hediger M., Romero M.F., Peng J.B., Rolfs A., Takanaga H., Bruford E.A. 2004. The ABCs of solute carriers: physiological, pathological and therapeutic implications of human membrane transport proteins. Pfluegers Arch. 447:465–468
Horn M., Frantz S., Remkes H., Laser A., Urban B., Mettenleiter A., Schnackerz K., Neubauer S. 1998. Effects of chronic dietary creatine feeding on cardiac energy metabolism and on creatine content in heart, skeletal muscle, brain, liver and kidney. J. Mol. Cell. Cardiol. 30:277–284
Ipsiroglu O.S., Stromberger C., Has J., Hoger H., Mühl A., Stökler-Ipsiroglu S. 2001. Changes of tissue creatine concentrations upon oral supplementation of creatine-monohydrate in various animal species. Life Sci. 69:1805–1815
Karlsson J., Ungell A.L., Gråsjö, Artursson P. 1999. Paracellular drug transport across intestinal epithelia: influence of charge and induced water flux. Eur. J. Pharm. Sci. 9:47–56
Laforenza U., Gastaldi G., Rindi G. 1993. Thiamine outflow from the enterocyte: a study using basolateral membrane vesicles from rat small intestine. J. Physiol. 468:401–412
Nash S.R., Giros B., Kingsmore S.F., Rochelle J.M., Suter S.T., Gregor P., Seldin M. F., Caron M.G. 1994. Cloning, pharmacological characterization, and genomic localization of the human creatine transporter. Receptors Channels 2:165–174
Orsenigo M.N., Tosco M. Faelli A. 1991. Cl/HCO3 exchange in the basolateral membrane domain of ratjejunal enterocyte. J. Membrane Biol. 124:13–19
Orsenigo M.N., Tosco M., Faelli A. 1992. Basolateral Cl/HCO3 exchange in rat jejunum: evidence from H14CO3 uptake in membrane vesicles. Biochim. Biophys. Acta 1108:140–144
Orsenigo M.N., Tosco M., Faelli A. 1994. Rat jejunal basolateral membrane Cl/HCO3 exchanger is modulated by a Na-sensitive modifier site. J. Membrane Biol. 138:47–53
Parsons D.S. 1956. The absorption of bicarbonate-saline solutions by the small intestine and colon of the white rat. Quart. J. Exp. Physiol. 41:410–420
Peral-Rubio M.J., Garcia-Delgado M., Calonge M.L., Duran J.M., De La Horra M.C., Wallimann T., Speer O. Ilundain A.A. 2002. Human, rat and chicken small intestinal Na+-Cl−-creatine transporter: functional, molecular characterization and localization. J. Physiol. 545:133–144
Persinjn J.P., Van der Slik W. 1976. A new method for the determination of y-glutamyltransferase in serum. J. Clin. Chem. Clin. Biochem. 14:421–427.
Rudnick G., Clark J. 1993. From synapse to vesicle: the reuptake and storage of biogenic amine neurotransmitters. Biochim. Biophys. Acta 1144:249–263
Schmitz J., Preiser H., Maestracci D., Ghosh B.K., Cerda J.J., Crane R.K. 1973. Purification of the human intestinal brush border membrane. Biochim. Biophys. Acta 323:98–112
Schoner W., Von Ilberg C., Kramer R., Seubert W. 1967. On the mechanism of Na+ and K+ stimulated hydrolysis of adenosine triphosphate. Purification and properties of Na+ and K+ activated ATPase from ox brain. Europ. J. Biochem. 1:334–343
Tosco M., Faelli A., Sironi C., Gastaldi G., Orsenigo M.N. 2004. A creatine transporter is operative at brush border level of rat jejunal enterocyte. J. Membrane Biol. 202:85–95
Van Dommelen F.S., Earner C.M., DeJonge H.R. 1986. Efficient entrapment of large and small compounds during vesiculation of intestinal microvilli. Biochem. J. 236:771–778
Walker J. 1979. Creatine: biosynthesis, regulation and function. Adv. Enzym. 50: 177–242
WaJlimann T., Wyss M., Brdiczka D., Nicollay K., Eppenberger H.M. 1992. Significance of intracellular compartmentation, structure and function of creatine kinase isoenzymes for cellular energy homeostasis: the phosphocreatine circuit. Biochem. J. 281:21–40
Walliman T., Hemmer W. 1994. Creatine kinase in non-muscle tissues and cells. Mol. Cell Biol. 133–134: 193–220
Wright S.H., Dantzler H. 2003. Molecular and cellular physiology of renal organic cation and anion transport. Physiol. Rev. 84:987–1049
Wyss M., Kaddurrah-Daouk R. 2000. Creatine and creatinine metabolism. Physiol. Rev. 80:1107–1213
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Orsenigo, M., Faelli, A., De Biasi, S. et al. Jejunal Creatine Absorption: What is the Role of the Basolateral Membrane?. J Membrane Biol 207, 183–195 (2005). https://doi.org/10.1007/s00232-005-0813-0
Received:
Revised:
Issue Date:
DOI: https://doi.org/10.1007/s00232-005-0813-0