
Abstract
Movement of fishes defines the distribution and abundance of populations and occurs on a range of spatial and temporal scales. To successfully parameterise assessments and design management strategies for exploited fish populations, knowledge and consideration of their movement patterns are essential. Since the efficacy of management approaches vary depending on the sedentary or mobile nature of the target species, presence, space use and depth utilisation were examined to understand the movement patterns of redthroat emperor Lethrinus miniatus. Sixty individuals were monitored for up to 12 months in an acoustic array comprising three coral reefs, and variation in space use and movement patterns was observed among 26 individuals. Half of the individuals were recorded only in proximity of one receiver along the reef edge, while the other half were detected at multiple receivers and used horizontal areas of approximately 4 km2. Periods of non-detection and lower detection frequency at night (χ 21 = 342.157, P < 0.001) indicated individuals may move away from the monitored reef edge to the adjacent sandy habitat, but most movements outside the array remain unknown. Long-distance movement was recorded for one individual, recaptured ~160 km from the release location. Generally, no trends in depth use were apparent, L. miniatus inhabited a variety of depths, which were not related to individual size or time of day, yet some effect of month was evident. Variation in movement among adult L. miniatus indicates that while some individuals undergo broader-scale movement, spatial closures that cover individual reefs (>4 km2) could provide protection from fishing for the proportion of the population that displayed high site fidelity and moderate-sized activity spaces (over a period of up to 12 months).
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Movement defines the distribution of species in time and space, and knowledge of movement patterns is useful for understanding population dynamics and potential effects of exploitation (Pittman and McAlpine 2001; Cadrin and Secor 2009). For exploited fish species, knowledge of whether a species is highly resident or mobile is important for stock assessments and the design of management regimes such as spatial or temporal fishery closures (e.g. Afonso et al. 2009) and effective placement of fish aggregation devices (e.g. Holland et al. 1990). Movement patterns occur at a range of scales, from small-scale habitat use (100s of m) to broad-scale movements (100s of km), and have been investigated using a variety of methods, including underwater surveys, tag-recapture, acoustic telemetry and otolith microchemistry (Kramer and Chapman 1999; Chapman and Kramer 2000; Palumbi 2004; Chateau and Wantiez 2009; Claisse et al. 2011; Currey et al. 2014). Movement information obtained using these methods has contributed to effective population management of exploited populations (Holland et al. 1993).
Passive acoustic telemetry enables the movement of multiple individuals fitted with transmitters to be monitored by an array of receivers and can provide movement data to be collected over multiple spatial and temporal scales (Heupel et al. 2006). The increased use of acoustic telemetry to study reef fish movement has provided information on a number of exploited species (e.g. Zeller 1997; Appeldoorn et al. 2009; Chateau and Wantiez 2009; Meyer et al. 2010; Marshell et al. 2011). Results indicate that some species have more active lifestyles than the sedentary behaviour often expected for adult reef fishes (Sale 1991), with daily movements including transitions to different habitats for resting, spawning and foraging (e.g. Holland et al. 1993, 1996; Appeldoorn et al. 2009; Claydon et al. 2012; Bunt and Kingsford 2014). Space use and trends in movement patterns, however, also vary among species and individuals (Quinn and Brodeur 1991; Chapman et al. 2012) suggesting that complex patterns exist and caution should be taken when generalising across and within species.
Emperors of the family Lethrinidae form an important component of tropical fisheries worldwide as commercial, recreational, subsistence and artisanal catch (Carpenter 2001). Lethrinids range in size from approximately 200–800 mm in length and display a diversity of life histories (Currey et al. 2013). One of the largest emperors, the redthroat emperor Lethrinus miniatus, is the most important emperor species in the commercial fisheries of Australia, Tonga, Japan and New Caledonia. In Australia, the 2011–2012 commercial harvest of L. miniatus was 237 t in Queensland (Fisheries Queensland 2013) and 62 t in Western Australia (Fletcher and Santoro 2013). This species is also common in the catch from Queensland charter (71 t) and recreational (90 t) fishers (Fisheries Queensland 2013). Yet, due to limited recaptures in conventional tag-recapture programs (e.g. B. Sawynock unpubl data; Egli et al. 2010), little information is available on the movement of emperors, and only a few studies have utilised acoustic telemetry to examine their movement. Nanami and Yamada (2009) investigated space use of Lethrinus harak by a snorkeler following individuals with a surface-towed hand-held GPS, and Meyer et al. (2010) obtained five days of acoustic telemetry data from Monotaxis grandoculis as part of a multi-species study. More recently, Taylor and Mills (2013) reported primarily small activity spaces for Lethrinus harak and Lethrinus obsoletus in Guam, suggesting small marine reserves may be effective for these species. Site fidelity and movement to nearby spawning areas have been documented for lethrinids, but movements associated with depth are unknown, only occurrence at depth has been recorded (Newman and Williams 1996; Fitzpatrick et al. 2012). Since management of emperor populations requires knowledge of their movements at different scales, longer-term information is needed. Presence, space use and depth utilisation of individuals can provide insight into the nature of emperors as sedentary or mobile, which is an important consideration for management strategies, e.g. if small spatial closures provide less protection to mobile rather than sedentary individuals. This study aimed to identify whether adult L. miniatus follow a sedentary or mobile lifestyle by using acoustic telemetry and tag-recapture information. Specifically, presence, activity space horizontally along the reef edge and vertically by depth and depth-use patterns of L. miniatus were analysed at three reefs in the southern Great Barrier Reef to better understand the patterns and scale of movement of this important exploited species.
Materials and methods
Fish sampling
Individual L. miniatus were collected by line fishing along the reef edges of Heron (23.4500°S, 151.9167°E) and One Tree (23.5083°S, 152.0917°E) Island reefs (Fig. 1) in the southern Great Barrier Reef (GBR), Australia. Fish were caught using rod and reel (13.6 kg line and 27.2 kg leader) with barbless 8/0 hooks baited with pilchard Sardinops sagax or squid Loligo spp. On capture, fish were vented and then anaesthetised with Aqui-S. A small incision was made longitudinally between the pectoral and ventral fins on the left side, a V13 transmitter with pressure (depth) sensor (V13P®, Vemco Ltd., Canada, 364 day battery life) was inserted into the body cavity and the incision closed with two simple interrupted stitches using absorbable sutures. Fork length (FL mm) was recorded and a dart tag (PDS; Hallprint©) fitted externally by insertion into the dorsal musculature for identification. The procedure took less than 8 min, and then, after recovery in fresh sea water, fish were released at the site of capture. A total of 60 adult individuals were fitted with transmitters in three deployments (20 each), in April 2011 and February and September 2012.

Location of Heron Island, Sykes and One Tree Island reefs in the Capricorn-Bunker group of the southern Great Barrier Reef, Queensland, Australia. Triangles indicate positions of acoustic receivers
Movement analysis
Long-term monitoring of the presence and movements of individuals was facilitated by 52 VR2W® acoustic receivers (Vemco Ltd., Canada) located around three reef platforms (Heron Island, Sykes and One Tree Island reefs; Fig. 1). This array was designed to track multiple species of mobile predators, with each receiver deployed on a star picket embedded in the substrate at 10–20 m depth (reef edge receivers) or <10 m (lagoon receivers). Acoustic receivers were downloaded twice per year. Data from receivers were analysed in the R environment (R Development Core Team: www.r-project.org). Detection range of receivers was examined through the use of six sentinel tags deployed at distances between 70 and 270 m from receivers located in different habitats (e.g. complex coral structure and sand) in the array. To account for variability in transmitter detection by time of day, hourly detections of sentinel tags over a period of months to years were analysed. Day was defined as 0600–1759 and night as 1800–0559 hours for all analyses, and the number of detections for each period compared using chi-squared tests. Mean detections of sentinel tags and L. miniatus per hour were plotted, and individual fish tags were standardised to the sentinel tag detections per hour according to Payne et al. (2010). Chi-squared tests were used to examine diel variation in mean number of standardised fish detections.
Individual fish were considered present on a given day when more than two detections were recorded per day by a given receiver. Detection data were examined to remove any spurious data that were the result of false detections. Unlikely fish positions were identified from visual inspection of detections with time and depth (scatterplots) for each individual and considered positions to be spurious when (1) less than two detections were made per receiver on any given day, (2) detections occurred after the programmed life of the transmitter, (3) represented fast, erratic movements (i.e. consistent with fish consumed by predator) or (4) detections lacked vertical movement aside from a consistent tidal signature (i.e. individual perished). Residency index was calculated as the number of days an individual was detected in the array divided by the period of detection (from deployment release date). Horizontal space use was calculated as measures of activity space (horizontal kernel utilisation distributions; hKUD) (Heupel et al. 2004) using the adehabitat package in R (Calenge 2006). Individuals detected for >5 days by two or more receivers were used in activity space analyses. Average positions (Simpfendorfer et al. 2002) were calculated for each individual at 2-h intervals and the core use areas (50 % hKUD) and extent of activity space (95 % hKUD) reported. Linear regressions tested whether days detected, residency or activity space were related to fish size (FL). Data were screened for normality and homogeneity of variances, and variables were log10 transformed when required.
Vertical activity space was estimated by vertical kernel utilisation distributions (vKUD) (Heupel and Simpfendorfer 2014) for individuals that were detected for >5 days using the ks package in R (Duong 2007). Rather than latitude and longitude, average positions (at 2-h intervals) were calculated in two-dimensional space of mean depth and distance along the reef. Receivers located along the reef edge of Heron Island Reef were represented as a linear system in this analysis (e.g. Simpfendorfer et al. 2008; Heupel and Simpfendorfer 2014), with linear positions of receivers along the reef edge calculated as the distance from the northwest point of the Heron Island Reef in a clockwise direction. Only reef edge receivers were included since L. miniatus rarely enters the lagoon (deep access points are limited; L. Currey unpubl data) and their presence was not detected by lagoon receivers. Positions of detections were calculated by adding the distance between each of the receivers from the receiver on which a detection occurred to the northwest point (in an anticlockwise direction around the reef) (Heupel and Simpfendorfer 2014). Core use areas (50 % vKUD) and activity space extent (95 % vKUD) by depth (m) and reef distance (km) were plotted for each individual. Relationships between activity space and fish size (FL) were tested using linear regressions.
Mixed effects models (restricted maximum likelihood estimation) were used to determine whether differences in depth use existed between month, day, hour and FL. Models were analysed using the nlme package in R (Pinheiro et al. 2013) with an information theoretic model selection process. Mean depth (m) was calculated for each individual, for each month (1–12), in each hour (0–23). Mean depth was treated as the response variable, and month, day, hour and FL were modelled as fixed factors. Individual was treated as a random effects term to account for the lack of temporal and spatial independence among individuals and reduce the effect of individuals with more detections. Models were compared using the Akaike information criterion corrected for small sample bias (AICc) to determine the best fitting model. Fifteen models were compared using a stepwise approach, from the most complex fully saturated model to the simplest model. The best-approximating model was considered as the model with the lowest ∆AICc and the fewest terms. Each model was compared to the null model to evaluate the relative strength of fit of each model to the data.
Results
Detection range of the receivers in the array was estimated as 270–340 m from sentinel tag detections and the known position of a non-detected recaptured individual (ID 4031). Receivers demonstrated with twice the detection ability during the day (chi-squared test, χ 21 = 2164.95, P < 0.001). This diel pattern was also reflected in detections of tagged L. miniatus. Standardising fish detections to sentinel tag detections per hour controlled for any difference in receiver detection ability and the frequency of detections recorded remained significantly higher during daytime hours (chi-squared test, χ 21 = 342.157, P < 0.001; Fig. 2). Greatest detections occurred during crepuscular hours (Fig. 2).

Mean number of detections per hour for L. miniatus before (solid line) and after standardising (bold solid line) to sentinel tag detections (dashed line). Hours of night are represented by grey boxes
Presence, residency and site fidelity
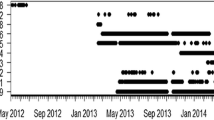
Tagged L. miniatus ranged in size from 323 to 493 mm FL and 48 of the 60 fish (80 %) were detected over 1–332 days (Fig. 3). Two individuals (ID 4026 and ID 4029) displayed active movement post-release, but after 6 and 7 days, respectively, movements were consistent with the tidal signature, suggesting these individuals perished or shed the transmitter. Although transmitter shedding is considered unlikely in this species, it cannot be ruled out as an explanation for movement cessation. Detections after this period for these individuals and spurious detections for other individuals were removed prior to analysis. After removal of these detections, 26 fish (52 %) were detected for 5 days or more (mean ± SE = 86 ± 21 days). A weak positive relationship was observed between FL and the number of (log10 transformed) days detected (linear regression, r 2 = 0.13, F 1,24 = 4.74, P = 0.040).

Presence plot of L. miniatus at One Tree Island Reef (ID 4018) and Heron Island Reef (all other individuals) in the Capricorn-Bunker region per day. Each point represents one day detected (with >2 detections)
Residency index varied among individuals from 0.01 to 1 (mean ± SE = 0.31 ± 0.07, n = 26; Table 1) and was not related to FL (linear regression, r 2 = 0.04, F 1,24 = 1.01, P = 0.326). Over 77 % of individuals were detected for less than 50 % of the monitoring period, demonstrated by a residency index < 0.5 (Table 1). Individuals were only detected on the receivers located around the reef edge, with no detections on lagoon receivers. The number of receivers visited ranged from 1 to 6 (mean = 2.19) with half of the individuals (n = 13) detected only at one receiver over an average of 108 days, indicating high site fidelity. A further four individuals (15 %) were detected at two receivers only. Failure to detect L. miniatus on acoustically monitored reefs nearby (i.e. Sykes and One Tree Island reefs) indicated a lack of movement between these reefs.
Horizontal activity space and recaptures
Activity space use was calculated for 13 individuals that were detected by two or more receivers. Both the core use (50 % hKUD) and extent of activity space (95 % hKUD) among individuals were small in area, with mean ± SE values of 0.36 ± 0.04 km2 (50 % hKUD) and 1.93 ± 0.27 km2 (95 % hKUD; Table 1). The maximum amount of horizontal space used was <4 km2 (Table 1). No relationship was observed between FL and 50 % hKUD (log10 transformed) (polynomial regression: r 2 = 0.16, F 2,9 = 0.84, P = 0.464), or between FL and 95 % hKUD (polynomial regression: r 2 = 0.03, F 1,10 = 0.35, P = 0.569).
Two individuals were recaptured during the study and provided contrasting results. One individual (ID 4031) was caught approximately 84 m from the tagging release site after 132 days at liberty but was not detected within the receiver array (approximately 340 m from nearest receiver). Transmitter malfunction is a possible explanation for non-detection, yet it is also possible that this individual utilised an activity space between receiver detection ranges, resulting in lack of detection. This behaviour would be consistent with the 17 individuals that showed restricted (e.g. presence at a single receiver) horizontal movement. The second (and largest overall) individual (ID 6707) was recaptured by a fisher on Abraham Reef, approximately 160 km (linear distance) from the release site (Fig. 4). This individual was at liberty for 495 days and had moved across the sandy Capricorn Channel (80–120 m in depth). Prior to departure, this individual was detected on the north side of Heron Island Reef for 264 days (out of 350 days) between three receivers (21, NW, 20) and displayed high residency (Fig. 4a). On the last day of detection (25/01/2013), this individual was detected at receiver 21, then 2 h 27 min later to the east at receiver NE (detected for 5 min), and finally 37 min later at receiver 22, where it was only detected for 7 min before detections ceased (Fig. 4a). Recapture at Abraham Reef (Fig. 4b) occurred 145 days after departure. Thus, evidence for both long-distance and limited movement was gained from recapture data.

Movement of individual ID 6707 at Heron Island Reef (a) indicating release point (cross symbol), movement over 11 months (solid arrows) between receivers (triangles; 20, NW, 21) and movement over the last day of detections (dashed arrows between receivers 21, NE and 22). Long-distance movement of approximately 160 km is illustrated (b) from Heron Island Reef of the Capricorn-Bunker reefs to the recapture location at Abrahams Reef of the Swain reefs
Vertical activity space and depth utilisation
Patterns in vertical activity space (vKUD) were estimated from depth use (m) and linear movement among receivers (km) for the same 13 individuals assessed for horizontal activity space (Table 1). Individuals used small areas and were detected from 0 to 30 m in depth, since bathymetry within detection range of receivers was <30 m. Despite location (northwest or southwest Heron Island Reef), vertical core use area (50 % vKUD) varied among individuals, with 23 % of individuals using shallow areas (≤10 m), 23 % >10 m, and 54 % utilising a range of depths (e.g. Fig. 5a, b). Vertical activity space extent (95 % vKUD) ranged between the surface and approximately 20 m with areas overlapping for some individuals but not for others (e.g. Fig. 5a, b). No clear patterns in depth use were present, as mean depth use varied among individuals (Fig. 6). Some individuals displayed relatively consistent depth use with month and time of day, while others were more variable. Individual ID 4027 displayed an interesting pattern in depth use by month, moving from shallow to deeper habitat use between May and October (Fig. 7). No relationship was observed between FL and 50 % vKUD (linear regression: r 2 < 0.01, F 1,10 < 0.01, P = 0.980; or polynomial: r 2 = 0.03, F 2,9 = 0.15, P = 0.861), or between FL and 95 % vKUD (linear regression: r 2 = 0.11, F 1,10 = 1.28, P = 0.284; or polynomial: r 2 = 0.14, F 2,9 = 0.73, P = 0.508).

Depth and space utilisation of L. miniatus at northwest (a) and southwest (b) Heron Island Reef. Activity space estimates of 50 % KUD (solid colour) and 95 % KUD (lines) are indicated for three individuals at each location, a ID 7045, ID 6710 and ID 6707 (dotted, solid and dashed lines, respectively); b ID 6721, ID 4027 and ID 4022 (solid, dashed and dotted lines, respectively) with reef distance (m) from the northwest point of Heron Island Reef. Location and depth of receivers are indicated by triangles

Mean depth (m) use of individuals by fork length (mm). Boxes represent the 25–75th percentiles, the median is represented by lines, and whiskers indicate 10th and 90th percentiles

Vertical KUDs of individual 4027 by month (April to October), representing depth use (m) by reef distance (m). Core use area (50 % vKUDs) is indicated by solid colour within the extent of activity space (95 % vKUDs) represented by circles. Triangles denote location and depth of acoustic receivers
Mean depth utilisation was best described by one model that included month as the only parameter (Table 2). This model (M1) predicted shallower mean depths were used in February compared with other months. Although the effect of month appeared minimal, this model provided a significantly better fit to the data than the null model (Table 2), and ∆AICc values were >2 for all other models.
Discussion
Lethrinids have been portrayed as both sedentary as adults with small home ranges (Sale 1991; Taylor and Mills 2013) and as mobile predators (Jones 1991; Carpenter 2001) that potentially migrate long distances (Williams et al. 2010; Currey et al. 2014), although limited empirical evidence has been collected to date. Results from this study indicate that individual L. miniatus display variability in presence, residence and depth utilisation, with an absence of consistent trends based on time (month, day and time of day) or size of individuals. Intra-specific variability in movement is common among reef fish (e.g. Kalish 1991; Kaunda-Arara and Rose 2004a, b; Marshell et al. 2011; O’Toole et al. 2011), with a general pattern involving a proportion of the population that remains resident while the other proportion moves greater distances (Grüss et al. 2011; Chapman et al. 2012). This study revealed that the majority of individuals were detected in a small area (<4 km2) for periods of up to 12 months. Despite limited horizontal movement, individuals often utilised the entire water column (to 30 m in depth). From what movement data exist for lethrinid fishes, the majority of individuals appear to display variable depth use and high site fidelity, with more extensive movement by a small contingent of the population.
Within-population dispersal variability may be more widespread among fishes than originally thought. Residents are individuals that do not make migratory movements, while migratory individuals disperse more broadly (Chapman et al. 2012). Based on telemetry data in this study, horizontal movement of L. miniatus can be categorised into movements at small (<1 km) and medium (few km) scales for residents and at a large scale (10s to 100s km) for migrants.
Small-scale movements along the reef edge were observed for half (13) of the L. miniatus individuals tracked, which visited a single receiver located adjacent to their capture and release location over periods of up to 12 months. Although mainly detected during the day, this result is consistent with other studies where lethrinids have been located in the same area months after release by acoustic monitoring (Taylor and Mills 2013), underwater surveys (Nanami and Yamada 2009) and recapture data (Kaunda-Arara and Rose 2004a). High site fidelity was displayed by individuals that inhabited the reef edge at distances of 270–340 m either side of a receiver. Similar mean minimum activity space size has been reported for other species such as 547 m for Kyphosus sectatrix (Eristhee and Oxenford 2001) and 223 m for Plectropomus leopardus (Zeller 1997). Although some L. miniatus were observed to be highly resident (residency index > 0.7), many individuals had a low residency index (< 0.3). Spending less than half of their time within the detection range of a receiver, individuals with a low residency index may have been undetected due to structural habitat complexity (sites were characterised by dense coral cover or bommies on sand, e.g. Claisse et al. 2011), residence adjacent to a detection area (and thus not detected), or moved to areas outside the detection range. Considering the estimated detection range, individuals further than 270–340 m from a receiver would not be detected. Although ID 4031 was recaptured close to the release location after four months at liberty and could indicate site fidelity, movement patterns of other individuals outside the array detection range are unknown. Use of small areas (< 0.004 and 0.029 km2) for Lethrinus harak and (0.008 km2) for L. obsoletus has been reported (Nanami and Yamada 2009; Taylor and Mills 2013). However, individuals were not observed at night (Nanami and Yamada 2009) or left the monitored area for an average of four days during spawning season (Taylor and Mills 2013), which indicate the activity space of these species may be larger than estimated. It is also possible that L. miniatus may move away from the reef edge at night (and thus beyond detection range) to forage in nearby sandy habitats (Carpenter 2001), like the congener Lethrinus nebulosus (Chateau and Wantiez 2008). Standardised individual fish detections were significantly greater during the day than at night lending support to this hypothesis. A diel pattern of movement between spatially separate habitats for feeding and resting is common for many reef fishes, with many species feeding at night in sand adjacent to reef areas (e.g. Hobson 1973; Holland et al. 1993; Mazeroll and Montgomery 1998; Meyer et al. 2010). Thus, results better reflect day time space use and could be an underestimate of total activity space. No inter-reef movements between the other monitored reefs were recorded. Therefore, further research involving receivers deployed away from the reef edge and enhanced coverage could investigate whether L. miniatus move to nearby habitats at night or are located close to the reef edge.
Medium-scale movement was exhibited by half (13) of the tagged individuals, which roamed more widely along the reef to a number of receivers. Activity spaces incorporated the area adjacent to the reef (largely during the day) and 95 % hKUDs for medium-scale movements extended up to 3.7 km2. These estimates are relatively small in area compared with the daily linear distances mobile predators such as Sphyraea barracuda travel (e.g. 12 km, O’Toole et al. 2011), but are similar to those for other lethrinids. Consistent movement outside coastal boundaries of Kenyan marine reserves (6.3 and 10 km2 in size) by L. mashena and L. miniatus demonstrated travel across approximately 5 km of continuous reef, with the distance moved positively associated with days at liberty for L. miniatus (like the current study but to a lesser degree; Kaunda-Arara and Rose 2004a). Similarly, Kaunda-Arara and Rose (2004b) demonstrated that each of the L. miniatus (29 % of the 348 tagged fish) and L. nebulosus (56 % of 93 fish) recaptured inside and outside of marine reserves were within 5 km of their release site. Thus, these studies support the observations here of site fidelity and small-moderate activity space for L. miniatus.
Variability in L. miniatus movement patterns also indicated large-scale movements by a contingent of the population. Long-distance movements of reef fish have been recorded in a number of studies; however, they are less common occurrences. It is uncertain whether the paucity of large-scale movement data is evidence of the rarity of these occurrences or whether it is simply a reflection of fishing effort. Kaunda-Arara and Rose (2004b) reported movements of 30–180 km by small numbers of three species (Siganus sutor, Plectorhinchus flavomaculatus and L. mahsena), which represented 0.2 % of the individuals tagged. The lethrinid travelled 148 km from the release point in 63 days which fits with long-distance movements of L. miniatus recaptured here (~160 km, current study) and two individuals recaptured in the same region that travelled in the same direction (180 km and 220 km: B. Sawynock, unpubl data; Williams et al. 2010). Similarly, carangids detected in an acoustic array close to their release location in Hawaii showed high site fidelity, with 75.5 % of individuals recaptured within 0.5 km of their release site, while one individual was located 72 km from the study site (1.2 % of recaptures). Long-distance movement over longer time periods (multiple years) was hypothesised to explain the regional differences in L. miniatus population demographics (Williams et al. 2010), with net migration of individuals among regions. Further, isotopic signatures (δ18O and δ13C) in L. miniatus otoliths suggested potential directionality of movement with ontogeny, over a scale of 10s of kilometres (Currey et al. 2014). Although some individuals remained in a similar environment through life, differences in δ18O and δ13C with ontogeny indicated the majority of juvenile individuals inhabited isotopically different environments to adults (Currey et al. 2014). These long-distance movements by a contingent of the population may be important for the connectivity and replenishment of populations (Kaunda-Arara and Rose 2004b; Grüss et al. 2011), allowing the mixing of populations necessary for maintaining genetic diversity. This is a potential explanation for the broad-scale movement of L. miniatus that occurs across both connected and fragmented habitat.
Depth utilisation has typically been reported for pelagic or deep-sea species through use of archival tags, sonar, catch records and acoustic telemetry (e.g. Brill et al. 1999; Brill and Lutcavage 2001; Musyl et al. 2003; Cartamil and Lowe 2004; Godo et al. 2004). However, little research has focused on reef fish movement within the water column. Studies have concentrated on estimates of reef fish abundance with depth, for example, larger Lutjanus kasmira were reported to occur on shallow reef slopes while smaller individuals inhabit deeper slopes and tongue-and-groove habitats (Friedlander et al. 2002). Depth use has also been inferred from acoustically tagged individuals via the placement of receivers at different depths, e.g. the majority of Zebrasoma flavescens were found more frequently in shallow areas <10 m depth (Claisse et al. 2011). However, directly quantifying vertical position over time allows a better understanding of depth-use patterns of reef fishes at a temporal scale.
Depth use of L. miniatus was not related to time of day or size of fish and was variable among individuals. Variation in individual depth use has been reported for other reef and temperate species, with repeatable movement patterns related to the time of day for some (Bellquist et al. 2008), while others displayed variation in movement among individuals (Starr et al. 2002). Although some L. miniatus displayed greater use of the water column during the night, the pattern was not consistent, and some individuals may have moved to deeper sandy areas that exist out of the detection range. Some individuals used consistent depths throughout the day, and others varied among depths with no apparent pattern. For other species such as trevally Pseudocaranx dentex, vertical movements of offshore-tagged individuals were more associated with currents and tides than for the smaller inshore-tagged individuals that mainly remained at depths of around 10 m (Afonso et al. 2009). Interestingly, like the overlap observed in horizontal space use for other lethrinids (Nanami and Yamada 2009; Taylor and Mills 2013), L. miniatus utilised vertical and horizontal activity spaces that somewhat overlapped, further highlighting their non-territoriality (Carpenter 2001). Vertical movement of L. miniatus was highly variable among individuals and time of day, suggesting no predictable patterns are present.
Patterns in depth use not only occur diurnally, but across a number of temporal scales. Taylor and Mills (2013) identified diel movement patterns related to tides for some lethrinid individuals, as well as in relation to lunar phase linked to spawning activity. Month was the parameter that best explained the variation in mean depth use of L. miniatus in this study. However, there was no strong trend in mean depth observed with time of year. Individual ID 4027 was the only example that provided a clear shift in depth use over a number of months (and utilised shallower depths in winter). Differences in environmental factors such as water temperature, which fluctuate in a predictable manner with seasons throughout the year, may be related to movement of individuals. Thus, environmental parameters should be investigated as potential drivers for depth use over this temporal scale through further research.
It is important to consider the limitations of acoustic telemetry when interpreting and estimating reef fish movement patterns from presence data. Coral reef environments can be particularly difficult to operate within because receiver performance (i.e. detection range) can be low due to structural habitat complexity and environmental noise (Claisse et al. 2011; Welsh et al. 2012; Cagua et al. 2013). Further, receiver detection ranges are not static as the detectability of sentinel tags fluctuates with dynamic nature of the environment. Multiple habitat types including reef crests, reef slopes, sand and semi-enclosed lagoons may also influence the ability to detect fish because signal attenuation may differ between habitats. However, each receiver in this study was placed in a position to minimise barriers to transmitter signal (structurally complex reef), with an estimated receiver detection range of 270–340 m across sand and reef habitat. This detection range was within those recently reported for coral reefs: 60 and 90 m for receivers located on the reef base and reef crest, respectively (using smaller, less powerful Vemco V9 transmitters: Welsh et al. 2012); and 120 m (V9) and 530 m (V16) by Cagua et al. (2013) at reefs in the Red Sea. Cagua et al. (2013) also determined that topography and environmental noise held the greatest influence on detection distance compared with other environmental factors (e.g. wind, thermocline and depth). Likewise, Payne et al. (2010) illustrated how a reduction in detection frequency was caused by factors other than animal movement (e.g. increased environmental noise) and can provide opposite interpretations of animal movement patterns when corrected for. Monitoring detections of sentinel tags over time provided frequencies of detection over a specified time period. After standardising the fish data to sentinel data as suggested by Payne et al. (2010), fewer L. miniatus detections were observed at night and greater fish activity was observed during the day. Additionally, the ability to monitor sentinel detections over a long time period (e.g. months) rather than days (e.g. Welsh et al. 2012) ensured temporal fluctuations in environmental conditions were adequately incorporated in this study.
Movement patterns and horizontal and vertical space use of L. miniatus were highly variable among individuals. Long-distance travel by at least one migratory individual, periods of non-detection and potential movement away from the reef edge at night provide potential evidence to support a more mobile lifestyle. Yet, it is uncertain how common long-distance movements are, and whether undetected individuals travelled far or remained nearby the array, so further research is required. For the majority of individuals, presence in the array over a period of up to 12 months indicated a relatively small area of reef edge space use, where some individuals appeared resident at one receiver while others moved more broadly along the reef edge. Therefore, the variability in activity space estimates (for individuals over periods of up to 12 months) indicates that spatial management measures that encompass individual reefs (> 4 km2) could provide protection from fishing for a substantial proportion of the adult L. miniatus population.
References
Afonso P, Fontes J, Holland KN, Santos RS (2009) Multi-scale patterns of habitat use in a highly mobile reef fish, the white trevally Pseudocaranx dentex, and their implications for marine reserve design. Mar Ecol Prog Ser 381:273–286
Appeldoorn RS, Aguilar-Perera A, Bouwmeester BLK, Dennis GD, Hill RL, Merten W, Recksiek CW, Williams SJ (2009) Movement of fishes (Grunts: Haemulidae) across the coral reef seascape: a review of scales, patterns and processes. Caribb J Sci 45(2–3):304–316
Bellquist LF, Lowe CG, Caselle JE (2008) Fine-scale movement patterns, site fidelity, and habitat selection of ocean whitefish (Caulolatilus princeps). Fish Res 91:325–335
Brill RW, Lutcavage ML (2001) Understanding environmental influences on movements and depth distributions of tunas and billfishes can significantly improve population assessments. Am Fish Soc Symp 25:179–198
Brill RW, Block BA, Boggs CH, Bigelow KA, Freund EV, Marcinek DJ (1999) Horizontal movements and depth distribution of large adult yellowfin tuna (Thunnus albacares) near the Hawaiian Islands, recorded using ultrasonic telemetry: implications for the physiological ecology of pelagic fishes. Mar Biol 133(3):395–408
Bunt CM, Kingsford MJ (2014) Movement, habitat utilization and behaviour of coral trout Plectropomus leopardus during and after the reproductive period on the southern Great Barrier Reef. Mar Ecol Prog Ser 496:33–45
Cadrin CX, Secor D (2009) Accounting for spatial population structure in stock assessment: past, present, and future. In: Beamish RJ, Rothschild BJ (eds) The future of fisheries science in North America, vol 31. Fish and fisheries series. Springer, Dordrecht, pp 405–426
Cagua EF, Berumen ML, Tyler EHM (2013) Topography and biological noise determine acoustic detectability on coral reefs. Coral Reefs 32:1123–1134
Calenge C (2006) The package adehabitat for the R software: tool for the analysis of space and habitat use by animals. Ecol Model 197:516–519
Carpenter KE (2001) Lethrinidae. Emperor (emperor snappers). In: Carpenter KE, Niem V (eds) FAO species identification guide for fishery purposes. The living marine resources of the Western Central Pacific. Volume 5. Bony fishes part 3 (Menidae to Pomacentridae). FAO, Rome, pp 3004–3051
Cartamil DP, Lowe CG (2004) Diel movement patterns of ocean sunfish Mola mola off southern California. Mar Ecol Prog Ser 266:245–253
Chapman MR, Kramer DL (2000) Movements of fishes within and among fringing coral reefs in Barbados. Environ Biol Fishes 57:11–24
Chapman BB, Skov C, Hulthén K, Brodersen J, Nilsson PA, Hansson L-A, Brönmark C (2012) Partial migration in fishes: definitions, methodologies and taxonomic distribution. J Fish Biol 81(2):479–499
Chateau O, Wantiez L (2008) Human impacts on residency behaviour of spangled emperor, Lethrinus nebulosus, in a marine protected area, as determined by acoustic telemetry. J Mar Biol Assoc UK 88(4):825–829
Chateau O, Wantiez L (2009) Movement patterns of four coral reef fish species in a fragmented habitat in New Caledonia: implications for the design of marine protected areas. ICES J Mar Sci 66:50–55
Claisse JT, Clark TB, Schumacher BD, McTee SA, Bushnell ME, Callan CK, Laidley CW, Parrish JD (2011) Conventional tagging and acoustic telemetry of a small surgeonfish, Zebrasoma flavescens, in a structurally complex coral reef environment. Environ Biol Fishes 91:185–201
Claydon JAB, McCormick MI, Jones GP (2012) Patterns of migration between feeding and spawning sites in a coral reef surgeonfish. Coral Reefs 31(1):77–87
Currey LM, Williams AJ, Mapstone BD, Davies CR, Carlos G, Welch DJ, Simpfendorfer CA, Ballagh AC, Penny AL, Grandcourt EM, Mapleston AJ, Wiebken AS, Bean K (2013) Comparative biology of tropical Lethrinus species (Emperors): challenges for multi-species management. J Fish Biol 82:764–788
Currey LM, Heupel MR, Simpfendorfer CA, Williams AJ (2014) Inferring movement patterns of a coral reef fish using oxygen and carbon isotopes in otolith carbonate. J Exp Mar Biol Ecol 456:18–25
Duong T (2007) ks: Kernel density estimation and kernel discriminant analysis for multivariate data in R. J Stat Softw 21:1–16
Egli DP, Moy W, Naisilisili W (2010) Fish movement in MPAs on coral reefs in Kubulau, Fiji. Technical report no. 05/10. Wildlife Conservation Society-Fiji, Suva
Eristhee N, Oxenford HA (2001) Home range size and use of space by Bermuda chub Kyphosus sectatrix (L.) in two marine reserves in the Soufriere Marine Management Area, St Lucia, West Indies. J Fish Biol 59(A):129–151
Fisheries Queensland (2013) Coral reef fin fish fishery 2011–12 fishing year report. Department of Agriculture, Fisheries and Forestry, Brisbane
Fitzpatrick BM, Harvey ES, Heyward AJ, Twiggs EJ, Colquhoun J (2012) Habitat specialization in tropical continental shelf demersal fish assemblages. PLoS ONE 7(6):e39634
Fletcher WJ, Santoro K (2013) Status reports of the fisheries and aquatic resources of Western Australia 2012/13: the state of the fisheries. Department of Fisheries, Western Australia, Perth
Friedlander AM, Parrish JD, DeFelice RC (2002) Ecology of the introduced snapper Lutjanus kasmira (Forsskal) in the reef fish assemblage of a Hawaiian bay. J Fish Biol 60:28–48
Godo OR, Hjellvik V, Iversen SA, Slotte A, Tenningen E, Torkelsen T (2004) Behaviour of mackerel schools during summer feeding migration in the Norwegian Sea, as observed from fishing vessel sonars. ICES J Mar Sci 61(7):1093–1099
Grüss A, Kaplan DM, Guénette S, Roberts CM, Botsford LW (2011) Consequences of adult and juvenile movement for marine protected areas. Biol Conserv 144:692–702
Heupel MR, Simpfendorfer CA (2014) Importance of environmental and biological drivers in the presence and space use of a reef-associated shark. Mar Ecol Prog Ser 496:47–57
Heupel MR, Simpfendorfer CA, Heuter RE (2004) Estimation of shark home ranges using passive monitoring techniques. Environ Biol Fishes 71:135–142
Heupel MR, Semmens JM, Hobday AJ (2006) Automated acoustic tracking of animals: scales, design and deployment of listening station arrays. Mar Freshw Res 57:1–13
Hobson ES (1973) Diel feeding migrations in tropical reef fishes. Helgolainder wiss. Meeresunters 24:361–370
Holland KN, Brill RW, Chang RKC (1990) Horizontal and vertical movements of yellowfin and bigeye tuna associated with fish aggregating devices. Fish Bull 88:493–507
Holland KN, Peterson JD, Lowe CG, Wetherbee BM (1993) Movements, distribution and growth rates of the white goatfish Mulloides flavolineatus in a fisheries conservation zone. Bull Mar Sci 52:982–992
Holland KN, Lowe CG, Wetherbee BM (1996) Movements and dispersal patterns of blue trevally (Caranx melampygus) in a fisheries conservation zone. Fish Res 25(3–4):279–292
Jones GP (1991) Post recruitment processes in the ecology of coral reef fish populations. In: Sale PF (ed) The ecology of fishes on coral reefs. Academic Press, San Diego, pp 294–328
Kalish JM (1991) Oxygen and carbon stable isotopes in the otoliths of wild and laboratory-reared Australian salmon (Arripis trutta). Mar Biol 110:37–47
Kaunda-Arara B, Rose GA (2004a) Out-migration of tagged fishes from marine reef national parks to fisheries in coastal Kenya. Environ Biol Fishes 70(4):363–372
Kaunda-Arara B, Rose GA (2004b) Long-distance movements of coral reef fishes. Coral Reefs 23:410–412
Kramer DL, Chapman MR (1999) Implications of fish home range size and relocation for marine reserve function. Environ Biol Fishes 55(1):65–79
Marshell A, Mills JS, Rhodes KL, McIlwain JL (2011) Passive acoustic telemetry reveals highly variable home range and movement patterns among unicornfish within a marine reserve. Coral Reefs 30(3):631–642
Mazeroll AI, Montgomery WL (1998) Daily migrations of a coral reef fish in the Red Sea (Gulf of Aqaba, Israel): initiation and orientation. Copeia 4:893–905
Meyer CG, Papastamatiou YP, Clark TB (2010) Differential movement patterns and site fidelity among trophic groups of reef fishes in a Hawaiian marine protected area. Mar Biol 157(7):1499–1511
Musyl MK, Brill RW, Boggs CH, Curran DS, Kazama TK, Seki MP (2003) Vertical movements of bigeye tuna (Thunnus obesus) associated with islands, buoys, and seamounts near the main Hawaiian Islands from archival tagging data. Fish Oceanogr 12(3):152–169
Nanami A, Yamada H (2009) Site fidelity, size, and spatial arrangement of daytime home range of thumbprint emperor Lethrinus harak (Lethrinidae). Fish Sci 75(5):1109–1116
Newman SJ, Williams DM (1996) Variation in reef associated assemblages of the Lutjanidae and Lethrinidae at different distances offshore in the central Great Barrier Reef. Environ Biol Fishes 46(2):123–138
O’Toole AC, Danylchuk AJ, Goldberg TL, Suski CD, Philipp DP, Brooks E, Cooke SJ (2011) Spatial ecology and residency patterns of adult great barracuda (Sphyraena barracuda) in coastal waters of the Bahamas. Mar Biol 158(10):2227
Palumbi SR (2004) Marine reserves and ocean neighbourhoods: the spatial scale of marine populations and their management. Annu Rev Environ and Resour 29(1):31–68
Payne NL, Gillanders BM, Webber DM, Semmens JM (2010) Interpreting diel activity patterns from acoustic telemetry: the need for controls. Mar Ecol Prog Ser 419:295–301
Pinheiro J, Bates D, DebRoy S, Sarkar D, R Development Core Team (2013) nlme: Linear and nonlinear mixed effects models. R package version 3.1-109
Pittman SJ, McAlpine CA (2001) Movements of marine fish and decapod crustaceans: process, theory and application. Adv Mar Biol 44:205–294
Quinn TP, Brodeur RD (1991) Intra-specific variations in the movement patterns of marine animals. Am Zool 31(1):231–241
Sale PF (1991) The ecology of fishes on coral reefs. Academic Press, New York
Simpfendorfer CA, Heupel MR, Hueter RE (2002) Estimation of short-term centers of activity from an array of omnidirectional hydrophones and its use in studying animal movements. Can J Fish Aquat Sci 59:23–32
Simpfendorfer CA, Heupel MR, Collins AB (2008) Variation in the performance of acoustic receivers and its implication for positioning algorithms in a riverine setting. Can J Fish Aquat Sci 65:482–492
Starr RM, Heine JN, Felton JM, Cailliet GM (2002) Movements of bocaccio (Sebastes paucispinis) and greenspotted (S. chlorostictus) rockfishes in a Monterey submarine canyon: implications for the design of marine reserves. Fish Bull 100(2):324–338
Taylor BM, Mills JS (2013) Movement and spawning migration patterns suggest small marine reserves can offer adequate protection for exploited emperor fishes. Coral Reefs 32(4):1077–1087
Welsh JQ, Fox RJ, Webber DM, Bellwood DR (2012) Performance of remote acoustic receivers within a coral reef habitat: implications for array design. Coral Reefs 31(3):693–702
Williams AJ, Little LR, Punt AE, Mapstone BD, Davies CR, Heupel MR (2010) Exploring movement patterns of an exploited coral reef fish when tagging data are limited. Mar Ecol Prog Ser 405:87–99
Zeller DC (1997) Home range and activity patterns of the coral trout Plectropomus leopardus (Serranidae). Mar Ecol Prog Ser 154:65–77
Acknowledgments
We thank the staff and students of the Fishing and Fisheries Research Group for assisting with this project, particularly A. Tobin, F. de Faria, S. Moore, E. Lédée, S. Munroe, V. Udyawer, M. Espinoza and J. Matley. Infrastructure and field support provided by the Australian Animal Tagging and Monitoring System facility of the Integrated Marine Observing System and the Australian Institute of Marine Science was greatly appreciated. Map data was provided by the Commonwealth of Australia (Great Barrier Reef Marine Park Authority), Google™earth and DigitalGlobe. Funding for this research was provided by the Australian Research Council awarded to M.R.H. Additional research funding was granted to L.M.C. from James Cook University (JCU) School of Earth and Environmental Sciences (SEES). L.M.C. was also supported by an AIMS@JCU Postgraduate Research Scholarship and stipend. We also thank the editor and two anonymous reviewers for their constructive comments on this manuscript. All research activities were conducted under GBRMPA permit numbers G10/33754.1 and G10/33758.1 and Queensland Department of Agriculture, Fisheries and Forestry permit number 144482. Treatment of all animals was conducted under ethical guidelines approved by JCU animal ethics number A1566.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by J. D. R. Houghton.
Rights and permissions
About this article

Cite this article
Currey, L.M., Heupel, M.R., Simpfendorfer, C.A. et al. Sedentary or mobile? Variability in space and depth use of an exploited coral reef fish. Mar Biol 161, 2155–2166 (2014). https://doi.org/10.1007/s00227-014-2497-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00227-014-2497-4