
Abstract
Investigations of distributional and density patterns of marine predators often reveal areas where high abundances of one or many species overlap in space and time (‘biological hotspots’); however, mechanisms underlying hotspot formation are often unclear, leading to difficulties determining spatial and temporal boundaries of protected areas. On the northeast Newfoundland coast, I previously described annually persistent aggregations of a key forage fish species, capelin (Mallotus villosus): (1) two pre- and post-spawning staging areas in deep (>150 m) bathymetric channels, (2) a cluster of four persistently used demersal spawning sites (17–40 m), and (3) a coastal migratory route (<50 m). Through at-sea surveys repeated over 8 years (2000–2003, 2007, and 2009–2011), I show that the majority of predator hotspots identified were spatially associated with (i.e., within 10 km) these persistent capelin areas for breeding seabirds (common murre: 85.2 ± 4.6 %; northern gannet: 66.9 ± 6.6 %), overwintering seabirds (great and sooty shearwaters: 88.0 ± 6.9 %), and baleen whales (humpback, minke, and fin whales: 86.8 ± 8.6 %). Most predator hotspots were closer to the spawning (3.8–14.0 km) relative to the staging areas (13.1–27.6 km), especially for murres and shearwaters. Interspecific differences were attributed to variation in maximum dive depths and dietary preferences. Predators only aggregated within the spawning area, while capelin were spawning, suggesting that interannual variation in association with predator and capelin hotspots was attributed to variation in survey timing relative to capelin spawning. As these areas of persistent capelin are bound by static bathymetric and large-scale oceanographic features and can be delimited in time based on the capelin spawning period, they may be important candidate areas for protection.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Increasing anthropogenic threats to marine predators, such as fisheries bycatch (Lewison et al. 2004), has led to a pressing need to identify important marine habitats for protection (e.g., Hooker and Gerber 2004; Baum et al. 2003; Worm et al. 2005). Investigations of marine predator distributional and density patterns often reveal areas where high abundances of one or many species overlap during certain periods of the year, often referred to as ‘biological hotspots’ (Hooker and Gerber 2004). At-sea surveys that integrate distributional patterns of multiple trophic levels simultaneously with physical habitat features on several temporal and spatial scales (e.g., Hunt et al. 1998; Ainley et al. 2009; Embling et al. 2012; Cox et al. 2013) provide insight into the biophysical features underlying these hotspots. Similarly, combining remote sensing data (e.g., depth, temperature, salinity, chlorophyll) with predator aggregations at sea, as revealed by tracks from predator-borne positioning devices, locations of high bycatch in fisheries and/or predator-focused surveys, has provided an understanding of predator-habitat associations for large predatory fish (e.g., Worm et al. 2005), seabirds (e.g., Louzao et al. 2009; Vilchis et al. 2006; Nur et al. 2011), sea turtles (e.g., Howell et al. 2008), and whales (e.g., Gill et al. 2011). The spatial and temporal scales of variability, or persistence, of habitat features can influence whether these areas are repeatedly used by predators (Davoren et al. 2003; Sigler et al. 2012), as higher persistence likely results in reduced time and energy spent searching for prey (e.g., Gende and Sigler 2006). Therefore, identifying mechanisms underlying the formation of biological hotspots and their persistence in space and time is critical for delineating candidate marine areas for protection (place-based management; Hyrenbach et al. 2000).
Capelin (Mallotus villosus) is the focal forage fish species in many northern marine ecosystems (Carscadden and Vilhjalmsson 2002). On the east coast of Newfoundland, capelin migrate from offshore wintering areas to the coast during the spring to spawn (Carscadden and Nakashima 1997), providing an abundant food source and attracting many breeding and nonbreeding seabirds and whales. The Funk Island Seabird Ecological Reserve lies approximately 60 km from the northeast Newfoundland coast. This island was the largest colony of the extinct great auk (Pinguinus impennis; Montevecchi and Tuck 1987) and includes the largest breeding colony of common murres and the fourth largest and most oceanic northern gannet colony in North America (Cairns et al. 1989). Many other breeding and nonbreeding seabird species are present near this colony during the summer, including puffins, kittiwakes, fulmars, gulls, and shearwaters along with baleen whales (if not provided in text, scientific names are provided in Table 1; Davoren 2007). Most of these species have high proportions of capelin in their diet during the summer (Brown et al. 1981; Whitehead and Carscadden 1985; Davoren and Montevecchi 2003; Davoren et al. 2010), as did the great auk (Hobson and Montevecchi 1991), indicating that many predators rely on capelin as prey in this area during the summer currently as well as historically.
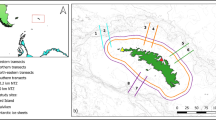
Within foraging ranges of breeding murres (60–80 km; Burke and Montevecchi 2009) and gannets (60–120 km; Garthe et al. 2011) from Funk Island, the bathymetry is characterized by gradually increasing depth from the coast, with the exception of two deep-water channels running perpendicular to the coast. Capelin shoals aggregate within these channels every year before and after spawning (Davoren et al. 2006). One staging area (‘Trench,’ Fig. 1b) is characterized by a deep bathymetric trench (180–200 m) bordered by a shallow shelf (~50 m) on both sides, where the water column is divided into three distinct layers: warm surface (0–50 m: 0–17 °C) and bottom (150–200 m: 0–4 °C) layers divided by cold water (50–150 m: <0 °C; Davoren et al. 2006). This cold intermediate layer (CIL) results from the southerly Labrador Current, which dominates the oceanography on the Newfoundland shelf (Petrie et al. 1988), resulting in few fine-scale oceanographic features in the study area (Davoren et al. 2006). The other staging area (‘Wadham,’ Fig. 1b) is located in a less pronounced channel (120–150 m) bordered by the 100 m contour. Within these staging areas, capelin exhibit diel vertical movement patterns (Davoren et al. 2006; Regular et al. 2010), whereby the bathymetry and thermal stratification features combine to allow capelin to remain in warm water at depth during the day to minimize predation by air-breathing predators and move into warmer surface water to feed at night (Davoren et al. 2006).

Location of the study area (square) on the northeast Newfoundland coast (a), indicating mesoscale survey lines (white dashed lines) and fine-scale survey line (Lumsden–Gull Island), along with known beach (diamonds) and deep-water, or demersal, spawning sites of capelin (stars), and highlighting four areas (circles): persistent capelin spawning area (Gull Island), persistent staging areas of capelin (Trench, Wadhman), and the Funk Island Seabird Ecological Reserve (b). The survey line along the coast represents the coastal migratory route of capelin. The gray scale indicates depth contours
From the southerly staging area (‘Trench’), capelin migrate north along the coast to beach and deep-water, or demersal, spawning sites (Davoren 2013; Fig. 1b). Four closely spaced (<0.5 km) demersal spawning sites near Gull Island (Fig. 1b) have been persistently used since their discovery in 2002 (Penton and Davoren 2012). These sites range in depth (17–40 m) and are characterized by bottom temperature (>2 °C) suitable for incubating eggs adherent to sediments. These sites are located in permanent bathymetric depressions (2–10 m deeper than the surrounding area), which retain suitable capelin spawning sediment (0.5–16 mm) by providing shelter from transport by bottom currents (Penton and Davoren 2012). Other demersal spawning sites not located in depressions are used for only 1–2 years after discovery, presumably due to the removal of suitable sediment by currents leaving exposed bedrock (Penton and Davoren 2012). After spawning, capelin move north of these spawning sites along the coast and then offshore to the northerly staging area (‘Wadham’; Fig. 1b; Davoren 2013). As capelin spawn in multiple waves (Davoren et al. 2008), capelin shoals are commonly observed in shallow water (<50 m) between the staging and spawning areas as well as at and nearby all areas (Davoren et al. 2006; Davoren 2013).
In this manuscript, I use annually repeated at-sea surveys to investigate whether high abundances of marine endothermic predators are spatially associated with areas of persistently high capelin abundance: (1) demersal spawning sites (‘Gull Island’ ~55 km from Funk Island; Fig. 1b); (2) persistent staging areas (‘Trench’ ~75 km from Funk Island; ‘Wadham’ ~25 km from Funk Island; Fig. 1b); and (3) the coastal migratory route of capelin (survey line along the coast within the 50 m depth contour; Fig. 1b). Using surveys conducted during 2000–2003, I previously showed that common murres, as well as the combined biomass of multiple top predators, were associated with persistent demersal spawning sites of capelin (Davoren et al. 2003; Davoren 2007). Here, I extend the temporal scale (2000–2003, 2007, and 2009–2011), include all persistent areas of capelin, and examine interspecific differences in association with the numerically dominant endothermic predators known to feed on capelin, including two breeding seabirds (common murres, northern gannets) as well as nonbreeding seabirds (sooty and great shearwaters) and baleen whales (fin, minke, and humpback whales). These species have contrasting energetic costs, speed, and primary modes of travel (swim, flight), foraging techniques (pursuit-dive, plunge-dive, pursuit-swim, surface-seize), maximum dive depths, and foraging range constraints (breeders constrained by colony location and nonbreeders not constrained), which likely influence the association of each predator species/group with each persistent prey aggregation. As the physical (bathymetric, oceanographic) characteristics of these persistent capelin areas are static among years (Davoren et al. 2006; Penton and Davoren 2012), this investigation may aid in delineating important marine habitats for protection in this region.
Materials and methods
Study design
To determine density and distributional patterns of seabirds and marine mammals, a mesoscale ship survey was conducted annually from 2000 to 2003, 2007, and 2009 to 2011. The survey consisted of nine east–west (across shelf) lines at a 9-km north–south spacing and one line along the coast (Fig. 1b). Survey lines were focused to the southwest of Funk Island initially due to traditional capelin fishing grounds (L. Easton personal communication), capelin spawning sites (Penton and Davoren 2012), and anecdotal observations of predator aggregations enroute to Funk Island, and later due to capelin–seabird–whale aggregations observed primarily along the coast (Davoren et al. 2003, Burke and Montevecchi 2009) and seabirds primarily returning to Funk Island from the coast (Davoren et al. 2003). Surveys were conducted 12 h day−1 during daylight (0800–2000 Newfoundland Daylight Savings Time = Greenwich Mean Time (GMT) minus 2.5 h) while traveling 11–15 km h−1. The route deviated slightly among years, owing to different wind direction and speed conditions, but the same paths were followed each year. Surveys from 2000 to 2003 were conducted aboard the 23 m Fisheries and Oceans research vessel Shamook in mid-late July, whereas those from 2007 to 2011 were conducted from the 15 m commercial fishing vessel Lady Easton II in late July–early August (Table 1). Surveys were consistently conducted during the chick-rearing period of breeding birds at Funk Island when capelin (total length: 60–200 mm) dominate the prey field in the study area (Davoren et al. 2006). Owing to variable timing of capelin spawning (up to 3 weeks; Davoren et al. 2012) and a 1–2 week shift in the timing of surveys among years, surveys varied in their temporal overlap with pre-spawning, spawning, and post-spawning periods of capelin among years (Table 1). During surveys in 2007 and 2009–2011, the most northerly line was not conducted because birds and mammals were infrequently observed.
To investigate whether predator abundance was influenced by prey abundance within one persistent prey area, I examined whether seabird and whale abundance near the persistent spawning sites increased during capelin spawning. A 10 km fine-scale survey from Lumsden to Gull Island (Fig. 1b) was repeated every 2–5 days from early July to mid-August during 2009–2011. During each fine-scale survey, the presence/absence of capelin eggs at the four persistently used demersal spawning sites near Gull Island was monitored to determine the timing of spawning.
Survey methods
During surveys, the number of seabirds was counted using standard strip-transect methods (method Ib, Tasker et al. 1984). One observer made continuous seabird counts from the bridge (2–3 m high) out to 300 m in a 90° arc from the tip of the bow to the port side of the vessel. A second observer was always present to help count when high-density aggregations were encountered. Marine mammals were counted both within and outside of the 300 m strip transect, conforming to a line-transect method. Although counting methods for marine mammals generally involve higher vantage points (Moulton and Mactavish 2004), consistent methods resulted in systematic biases over all surveys. Observer watches were undertaken continuously between 0800 and 2000, but never exceeded 2 h (range: 1–2 h) to avoid fatigue. Animal detectability conditions, including sea state (Beaufort wind force scale), visibility, glare, and precipitation, were recorded at the start of each survey and updated as conditions changed. Counts with behavioral descriptions (on water, feeding, flying, and direction) were entered on a laptop computer with counting software (D. Senciall, Birds and Beasty Counter, 1998, Fisheries and Oceans Canada, version 1.0). The laptop was interfaced with the navigational system of the vessel, and counting software was used to append a position (latitude and longitude) and GMT to each bird entry. Predators also were counted between ports and the start/end of survey lines as well as between lines, leading to variable areas covered among years in addition to the standard survey lines.
Analysis of survey data
Survey segments with lower detectability conditions (i.e., <300 m) were excluded from the analysis; however, this was minimal as good weather and visibility conditions were required to conduct offshore lines using 15–23 m ships. Counts of all shearwaters and gannets feeding, floating on the water, and flying within the strip transect were included in analyses; however, flying murres were excluded, as alcids are not likely to forage during flight. This focused the analysis on potentially foraging seabirds. To show interannual variability in the density of each species within the study area, the total number of each seabird and mammal species observed during each survey was divided by the total surveyed distance to calculate linear densities (animals km−1; Table 1). I also calculated the percentage each species contributed to the overall number of seabirds and mammals separately during each survey/year (Table 1).
To investigate the distributional and density patterns of the numerically dominant seabird species (common murre, northern gannet, sooty shearwater, great shearwater) in each year, predator counts were imported into ArcMap 10 (ESRI 2012). A 4.5 × 4.5 km grid was generated using the Vector Grid tool in ET Geowizards (Tchoukanski 2012) and superimposed on the study area. Grid cell size was set at 4.5 km2, similar to Davoren et al. (2003), because the maximum visual range of seabirds on the water is estimated to be 4.5 km (Haney et al. 1992), allowing individuals on the edge of cells to cue to the foraging activities of other marine predators within this range. The Spatial Join tool (ArcMap attribute table) was used to assign each predator count to a 4.5 km2 cell in the grid for each species in each year. The resulting attribute table was exported and summarized in Microsoft Excel 2011 to determine the total number of birds of each species in each 4.5 km2 cell in each survey year. The summarized data were imported back into ArcMap. Using the Spatial Join tool, I measured the distance between the center point of each 4.5 km2 cell and four points of interest: Funk Island, the persistent spawning area (Gull Island), and the persistent staging areas of capelin (Trench, Wadham; Fig. 1b). The same procedure was followed for baleen whales, but the total number of all baleen whale observations was summarized in each 4.5 km2 cell in each year instead of the total number of individuals, due limitations of counting methods. Overall, this provided data sets of the number of individuals per 4.5 km2 for each of the numerically dominant seabird species, and the number of observations per 4.5 km2 for all baleen whales, over the entire surveyed area in each year.
Within each year, all cells were categorized as having high, moderate, and low counts for each dominant seabird species separately. High, moderate, and low count categories were held constant across years. The frequency distribution of counts per 4.5 km2 cell was highly left skewed, indicating that the majority (70–90 %) of nonzero cells in the grid contained few birds, whereas very few (1–3 %) contained much higher counts, similar to other marine predator data sets (e.g., Sigler et al. 2012). Owing to this, I ordered the counts per 4.5 km2 cell for each species in each year from highest to lowest and then calculated the percentage that the cell count contributed to the total count, following Sigler et al. (2012). Cumulative percentages were calculated for each year separately, and all 4.5 km2 cells that contributed to the top 50 % of the total count for a species in a year were categorized as ‘high,’ 51–90 % as ‘moderate,’ and 91–100 % as ‘low.’ The lowest count values for ‘high,’ ‘moderate,’ and ‘low’ categories for each species in each year were determined, and the median value of each category over all years was used to standardize the categories across all years. For instance, the ‘high’ category for murres was defined as 120–2,500 birds per 4.5 km2 cell over all years. The procedure was similar for baleen whales; however, the number of observations was categorized as ‘low’ (i.e., one observation per 4.5 km2) and ‘high’ (i.e., more than one observation per 4.5 km2). All 4.5 km2 cells categorized as ‘high’ for each predator in each year are hereafter referred to as ‘hotspots.’ Interannual variation in abundances of some predators, especially baleen whales and shearwaters, and standardized count categories across years resulted in the absence of hotspots for some species in certain years.
To examine whether predator hotspots were associated with persistent spawning and staging areas of capelin (hereafter referred to as ‘prey hotspots’) within a year, I simply counted the number of predator hotspots in each year that were within a certain range of these previously defined prey hotspots. As capelin shoals continuously move between the persistent staging and spawning areas along the coast throughout spawning (Davoren 2013), predator hotspots were considered associated with each prey hotspot if they were present within 10 km of the center point of each prey hotspot. I also counted the number of predator hotspots found along the coastal migratory route of capelin, or the survey line along the coast within the 50 m depth contour, but outside of the spawning and staging-Trench prey hotspots (Fig. 1b). For breeding seabirds (murres, gannets), I counted the number of predator hotspots that occurred within 10 km of Funk Island, to examine whether these predators preferentially forage closer to the breeding colony, as predicted for central place foragers. Each area of interest represented a different percentage of the total surveyed area: 6 % spawning, 3 % staging-Trench, 10 % staging-Wadham, 4 % Funk Island, and 11 % coastal migratory route of capelin. Predator hotspots found outside these areas were classified as located in ‘other’ areas. I used chi-squared tests to investigate differences in the frequency of association of predator hotspots with persistent prey hotspots, the coastal migratory route, Funk Island, and other areas all years combined.
As another measure of association, I quantified the proximity of predator hotspots to each prey hotspot by calculating the distance of the center of each predator hotspot to the center of each prey hotspot. Mean distances reported are the distance from the edge of the prey hotspot to the edge of the predator hotspot (i.e., distance between the center of the prey hotspot and the center of the predator hotspot minus 12.25 km). To examine whether predator hotspots were closer to a particular prey hotspot, I compared the mean distances of predator to prey hotspots among prey hotspots for each predator species separately (all years combined) using ANOVAs. Similarly, I compared the mean distances of hotspots of breeding predators (murres, gannets) to Funk Island. I also examined interannual and interspecific differences using ANOVAs.
Timing of spawning
Capelin eggs adhere to sediment when fertilized. Therefore, the timing of spawning is easily determined by monitoring sediment for the presence of eggs. A 0.3 m2 Ponar Grab was used to collect sediment at or near (~50 m) the same location, determined by GPS, within the four closely spaced and persistently used spawning sites near Gull Island (Fig. 1b). Sediment was examined for egg presence starting in early July 2009–2011, and egg-bearing gravel was collected when eggs were first detected and continued until eggs were not found (Davoren et al. 2012). A 20 mL sample of eggs/sediment (200–500 eggs) was preserved in Stockard’s solution (% volume: 4 glacial acetic acid, 5 formaldehyde [37], 6 glycerin, and 85 water), which clears the yolk sac to determine egg developmental stages (Frank and Leggett 1981). At least 50 eggs from each sample were examined to determine the presence of eggs in early developmental stages (stages I–II), indicative of recent fertilization (Fridgeirsson 1976). The start of spawning was determined by the presence of eggs in samples on each day, whereas the end of spawning was determined by the absence of eggs in stages I–II. The total duration of spawning reflects the period shoals of spawning capelin were present at these four spawning sites.
Results
Over all years, 27 species of seabirds were encountered in the study area. The numerically dominant species was the common murre (43 %), followed by sooty shearwater (29 %), great shearwater (11 %), and northern gannet (8 %; Table 1). A total of 10 species of marine mammals was observed in the study area, numerically dominated by baleen whales (44 %), including humpback, fin, and minke whales, followed by dolphin spp. (34 %) and seal spp. (21 %). Species composition and relative densities varied among years. Shearwaters and dolphins were observed in higher densities, but seals in lower densities, when surveys were timed to overlap with capelin spawning (2007, 2009–2011) relative to earlier surveys (2000–2003). Predator hotspots were observed for murres and gannets in all years, whereas hotspots were observed in 5 years for sooty shearwaters, 2 years for great shearwaters, and 6 years for baleen whales. Due to interannual variation in abundance of shearwater species, I combined great and sooty shearwaters into ‘shearwaters.’
The distribution of predator hotspots in relation to the three persistent prey hotspots varied among years for murres (Fig. 2), gannets (Fig. 3), shearwaters (Fig. 4), and baleen whales (Fig. 5). With all years combined, a higher frequency of predator hotspots was associated with the three persistent prey hotspots relative to all other areas (i.e., Funk Island, coastal migratory route, and other) for murres (χ 21 = 10.903, p < 0.0001; Fig. 2; Table 2), shearwaters (χ 21 = 8.10, p < 0.001; Fig. 4; Table 2), and baleen whales (χ 21 = 4.167, p < 0.05; Fig. 5; Table 2), but not for gannets (χ 21 = 0.071, p > 0.05; Fig. 3; Table 2). Many gannet hotspots were associated with the breeding colony at Funk Island (~25 % per year; Table 2). Redoing the analysis with the three persistent prey hotspots combined with Funk Island resulted in a higher frequency of gannet hotspots associated with these four areas relative to all other areas (i.e., coastal migratory route and other; χ 21 = 5.786, p < 0.01; Fig. 3; Table 2). Predator hotspots of all species/groups were associated with the three persistent prey hotspots in all years of the study (Table 2). Within prey hotspots, a higher frequency of predator hotspots was associated with the spawning hotspot relative to the staging hotspots (Table 2), but this was only significant for shearwaters (χ 21 = 5.028, p < 0.05). Similarly, the number of years in which predator hotspots were associated with each prey hotspot varied, but murre and shearwater hotspots were associated with the spawning hotspot in all or most (80 %) years, respectively. Overall, the majority (~86.8–91.5 %) of predator hotspots were associated with persistent prey hotspots, the coastal migratory route as well as Funk Island, representing only 34 % of the surveyed area, with few hotspots (8.5–13.2 %) found outside these areas (Table 2).

Annual (2000–2003, 2007, and 2009–2011) distributional patterns of common murres on the water (blue circle 1–29 birds per 4.5 km2; green circle 30–119; red circle 120–2,500) during the mesoscale survey (white dashed line) on the northeast Newfoundland coast, highlighting the persistent capelin spawning and staging areas (black circles ~10 km2). See Fig. 1b for depth contours and labels

Annual (2000–2003, 2007, and 2009–2011) distributional patterns of all northern gannets flying and on the water (blue circle 1–9 birds per 4.5 km2; green circle 10–29; red circle 30–300) during the mesoscale survey (white dashed line) on the northeast Newfoundland coast, highlighting the persistent capelin spawning and staging areas (black circles ~10 km2). See Fig. 1b for depth contours and labels

Annual (2000–2003, 2007, and 2009–2011) distributional patterns of all great and sooty shearwaters flying and on the water (blue circle 1–19 birds per 4.5 km2; green circle 20–119; red circle 120–6,000) during the mesoscale survey (white dashed line) on the northeast Newfoundland coast, highlighting the persistent capelin spawning and staging areas (black circles ~10 km2). See Fig. 1b for depth contours and labels

Annual (2000–2003, 2007, and 2009–2011) distributional patterns of all humpback, minke, and fin whales (blue circle 1 whale observation per 4.5 km2; red circle >1) during the mesoscale survey (white dashed line) on the northeast Newfoundland coast, highlighting the persistent capelin spawning and staging areas (black circles ~10 km2). See Fig. 1b for depth contours and labels. Note baleen whales were not observed during the mesoscale survey in 2001 and 2003
The mean distances of predator hotspots to each persistent prey hotspot differed significantly within species (Table 3). Post hoc pairwise Tukey tests revealed that hotspots of each predator were significantly closer to the spawning hotspot relative to the Trench staging hotspot for murres (p = 0.001), gannets (p = 0.007), and shearwaters (p = 0.027), as well as relative to the Wadham staging hotspot for murres (p = 0.001), shearwaters (p = 0.008), and whales (p = 0.002). Alternately, whale hotspots were equidistant to the spawning and the Trench staging hotspots (p = 0.611) but were significantly closer to the Trench than the Wadham staging hotspot (p = 0.032; Fig. 5). Gannet hotspots were equidistant to the spawning and Wadham staging hotspot (p = 0.998) but were significantly closer to the Wadham than the Trench staging hotspot (p = 0.004; Fig. 3). Although hotspots of breeding birds (murres, gannets) were observed at or near Funk Island in some years (Table 2; Figs. 2, 3), hotspots of murres were significantly closer to all prey hotspots (p < 0.0001) than Funk Island, whereas gannet hotspots were significantly closer to the spawning hotspot (p = 0.045) and the Wadham staging hotspot (p = 0.026), but not the Trench staging hotspot relative to Funk Island (Figs. 2, 3).
As the distance of predator hotspots to each persistent prey hotspot was highly correlated within a year (e.g., far from Trench means close to Funk Island), I examined interannual variability in the distance of predator hotspots to the spawning prey hotspot due to its central position in the study area. The mean distance of predator hotspots to the spawning hotspot varied among years for murres (F 7,52 = 3.396, p = 0.005), being significantly farther north in 2007 relative to 2009 (p = 0.021; Fig. 2), and gannets (F 7,38 = 4.885, p = 0.001), being significantly farther north in 2000 than 2009 (p = 0.040) and in 2007 than 2003 (p = 0.034), 2009 (p = 0.003), and 2010 (p = 0.005; Fig. 3). The distance of predator hotspots to the spawning hotspot did not vary annually for shearwaters (F 4,32 = 0.767, p = 0.554; Fig. 4) or baleen whales (F 5,18 = 0.996, p = 0.448; Fig. 5).
Interspecific differences were observed in the distance of predator hotspots to the spawning prey hotspot (F 3,166 = 3.107, p = 0.028) and the Trench staging hotspot (F 3,166 = 3.587, p = 0.015), but not the Wadham staging hotspot (F 3,166 = 2.507, p = 0.061). These differences were due to shearwater hotspots being significantly closer than gannet hotspots to the spawning hotspot (p = 0.028) and the Trench staging prey hotspot (p = 0.010), especially in 2007. In addition, gannet hotspots were significantly closer to Funk Island than murre hotspots (F 1,90 = 8.307, p = 0.005; Table 2).
Predator abundance and timing of spawning
The total number of seabirds (murres, gannets, and shearwaters combined) along a standardized, 10 km survey line (Lumsden to Gull Island; Fig. 1b) within the spawning hotspot varied among years (2009–2011) but peaked soon after the date of first spawning and remained high for 15–20 days (Fig. 6). A slightly different pattern was observed in 2009 when capelin did not spawn at the four persistent spawning sites due to unusually cold water temperatures (Penton and Davoren 2012), but were present at the sites over a two-week period (Davoren 2013). Overall, the mean number of seabirds (all species combined) recorded within this spawning hotspot was significantly lower before and after capelin spawning (mean ± SE: 114.5 ± 39.7) relative to during spawning (828.4 ± 242.0; F 1,26 = 7.358, p = 0.012). Similarly, whales were present significantly more frequently during spawning (46.7 %) than before and after spawning (7.7 %; χ 21 = 5.184, p < 0.05).

Total number of common murres, northern gannets, and great and sooty shearwaters recorded during the 10 km fine-scale survey (Lumsden to Gull Island) within the capelin spawning hotspot in relation to the duration of capelin spawning at the four persistently used spawning sites of capelin on the northeast coast of Newfoundland during 2009–2011. Day 0 represents the date of first spawning over all four sites, negative values represent days before spawning, and positive values represent days after the first date of spawning. Spawning duration was 19 days in 2009, 14 days in 2010, and 12 days in 2011. Note in 2009, high abundances of spawning adults were present over 19 days at the sites, but spawning did not occur
Discussion
The majority (67–88 %) of high abundance areas, or ‘hotspots,’ of seabird and baleen whale predators examined in this study were associated with areas where their forage fish prey, capelin, could be persistently located, representing ~30 % of the surveyed area. Associations of predator hotspots with persistent capelin areas occurred in all years, and the distribution of 86.8–91.5 % of predator hotspots could be explained by associations with persistent areas of capelin as well as the breeding colony. Predator hotspots of all species were closest to the persistent capelin spawning hotspot, especially for shearwaters, whereas association with the two deeper staging prey hotspots varied among species. The total number of seabirds was significantly higher within the capelin spawning hotspot and whales were more frequently present during capelin spawning relative to before and after spawning, indicating that predators aggregate within this area after shoals of spawning capelin arrive at or nearby these spawning sites. Therefore, variation in the timing of capelin spawning in the study area (Davoren et al. 2012) in relation to survey timing (Table 1) likely resulted in the observed interannual differences in the presence, as well as density and distributional patterns, of these and other predator species.
The high percentage of predator hotspots associated with persistent capelin hotspots combined with the association of predator hotspots with persistent capelin hotspots in all years underscores the importance of the predictability of the food source to foraging site selection by predators. Marine predators may prefer sites where prey are predictably located because they can minimize energetically costly search activities or foraging effort (Gende and Sigler 2006), presumably by using memory-based techniques, as suggested for murres (Davoren et al. 2003) and shown for gannets in the study area (Garthe et al. 2007, 2011; Montevecchi et al. 2009). There is mounting evidence of memory-based foraging (e.g., Irons 1998; Hamer et al. 2001) and the preferential use of predictable aggregations of prey by seabirds within breeding seasons, as well as marine birds and mammals during nonbreeding when foraging ranges are not limited (i.e., noncentral place foragers; Gende and Sigler 2006; Sigler et al. 2012). On an annual scale, site fidelity to foraging grounds has been illustrated for humpback whales (e.g., Gulf of Maine, Clapham et al. 1993) and sooty shearwaters (e.g., Grand Bank, Hedd et al. 2012) in the Northwest Atlantic. The long-term persistence of highly abundant prey at particular locations could lead to local knowledge of established feeding grounds over generations (‘hinterland,’ Cairns 1989) by both breeding and overwintering predators.
A higher percentage of hotspots of most predators were associated with and closer to the spawning capelin hotspot relative to the staging hotspots, suggesting a higher relative attraction to the spawning area. As capelin shoals are primarily associated with the seabed in the study area (Davoren et al. 2006), capelin is more accessible at shallow (<40 m) spawning sites relative to the deeper (>150 m) staging areas. Shallower prey is especially important for species with lower maximum dive depths, including sooty shearwaters (maximum dive depth: ~70 m; Weimerskirch and Sagar 1996; Shaffer et al. 2006), great shearwaters (~19 m; Ronconi et al. 2010), and gannets (22 m; Garthe et al. 2000). Although murres perform deep dives (~150 m) in the study area, individuals have decreased foraging efficiency (i.e., bottom time relative to the dive cycle) at depth (Hedd et al. 2009), possibly explaining the higher association with the shallow spawning area. In contrast, baleen whales regularly exploit capelin shoals at depths in the staging areas (e.g., Whitehead 1983), possibly explaining the similar association of these whales with spawning and the Trench staging hotspots. Capelin exhibit diel vertical movement patterns within both staging areas (Davoren et al. 2006), thereby enhancing their accessibility to air-breathing predators near the surface at dawn and dusk. Although some species in the study area appear to take advantage of this higher accessibility to a greater extent (murres; Regular et al. 2010) than others (gannets, Davoren et al. 2010), diel vertical movement by capelin may explain the association of hotspots of shallow divers with staging areas, namely gannets with the Wadham staging area. Overall, predictable, highly abundant, and accessible prey likely result in critical foraging habitats for marine predators.
The association of gannet hotspots with persistent capelin hotspots differed from other predators, indicated by a lower association (67 %) relative to other predators (85–88 %), as well as a higher association (24.6 %) and proximity (25.1 km) to the breeding colony at Funk Island relative to murres (5.7 % and 33.3 km, respectively). The latter was unexpected because gannets have larger foraging ranges than murres in the study area (Burke and Montevecchi 2009; Garthe et al. 2011), presumably due to lower energy expenditure during flight. The former was expected, however, due to dietary variation of breeding gannets in the study area among years (e.g., Montevecchi 2007). Breeding gannets at Funk Island prefer large pelagic fishes (e.g., Atlantic mackerel Scomber scombrus, Atlantic saury Scomberesox saurus, Atlantic herring Clupea harengus), as capelin comprise a lower proportion of the diet when these larger fish are present within foraging ranges (Montevecchi 2007). In contrast, murres rely on capelin as their primary prey in the study area (Davoren and Montevecchi 2003), and capelin are important in the diets of shearwaters (Brown et al. 1981) and baleen whales (Whitehead and Carscadden 1985) in coastal Newfoundland. Breeding gannets dive in different locations in the study area when their diet shifts from primarily large pelagic species to capelin (Garthe et al. 2011); however, dive locations of breeding gannets returning to Funk Island with capelin as well as other species (e.g., saury, mackerel) are often concentrated within or nearby the Wadham staging area (Garthe et al. 2007, 2011; Garthe unpublished data). This suggests this site may represent important habitat for other fish species in addition to capelin. Overall, dietary preferences and interannual variability in the abundance and distributional patterns of large pelagic fish may explain the lower association of gannet hotspots with persistent capelin hotspots and proximity to the breeding colony.
Early spawning of capelin is associated with altered diets and prey switching of breeding murres and gannets in the study area, likely due to the absence of capelin at persistent spawning and staging hotspots during chick-rearing (Davoren et al. 2012). In support, hotspots of gannets and murres were found farther north of the persistent spawning hotspot in 2007, when surveys were conducted near the end of spawning, relative to 2009 (also 2003 and 2010 for gannets), when conducted during early to mid-spawning (Davoren et al. 2012). This suggests that the distribution patterns of breeding predators reflect the distribution of capelin along their spawning migration. In contrast, as nonbreeding predators are not constrained by foraging limits, shearwaters and baleen whales generally were in low abundance when the survey overlapped with the capelin pre-spawning (2000–2003) compared to spawning period (2007–2011). This suggests that these nonbreeding predators do not arrive in the study area until capelin abundance reaches a certain level, as has been shown for whales in coastal Newfoundland (Piatt and Methven 1992) as well as interannualy variable numbers of great and sooty shearwaters in coastal areas of Nova Scotia (e.g., Brown et al. 1981).
Hotspots of all predators were found outside of persistent capelin areas in most years, suggesting that other biophysical factors may also be important influences on the distribution of endothermic predators in the study area. These factors may include varying abundances of alternative prey, as shown for gannets in the study area (e.g., Garthe et al. 2011) and/or fine-scale spatial and temporal oceanographic features, as shown for other species in other regions (e.g., Hunt et al. 1998; Embling et al. 2012; Cox et al. 2013). The use of both persistent and nonpersistent areas suggests that predators likely use a variety of techniques to locate prey aggregations, including cueing to the foraging activities of other predators (murres to murres, Davoren et al. 2003; gannets to gannets, Montevecchi et al. 2009; gannets to whales, Davoren et al. 2010). The distribution of most predator hotspots (67–88 %), however, could be explained by associations with persistent areas of capelin. This provides compelling evidence that areas with persistently high densities of capelin play an important role in shaping the distributional patterns of endothermic predators in the study area and likely in supporting an important seabird colony in North America during a key period in the annual cycle as well as high numbers of nonbreeding seabirds and mammals overwintering in the area. These persistent capelin areas can be delimited in space, based on static bathymetric and large-scale oceanographic features and in time based on the capelin spawning period, suggesting that they have high conservation value (Hyrenbach et al. 2000) and may be important candidate areas for protection.
References
Ainley DG, Dugger KD, Ford RG, Pierce SD, Reese DC, Brodeur RD, Tynan CT, Barth JA (2009) Association of predators and prey at frontal features in the California Current: competition, facilitation, and co-occurrence. Mar Ecol Prog Ser 389:271–294
Baum JK, Myers RA, Kehler DG, Worm B, Harley SJ, Doherty PA (2003) Collapse and conservation of shark populations in the Northwest Atlantic. Science 299:389–392
Brown RGB, Barker SP, Gaskin DE, Sandeman MR (1981) The foods of great and sooty shearwaters Puffinus gravis and P. griseus in eastern Canadian waters. Ibis 123:19–30
Burke CM, Montevecchi WA (2009) The foraging decisions of a central place foraging seabird in response to fluctuations in local prey conditions. J Zool 278:354–361
Cairns DK (1989) The regulation of seabird colony size: a hinterland model. Am Nat 134:141–146
Cairns DK, Montevecchi WA, Threlfall W (1989) Researcher’s guide to Newfoundland seabird colonies. Memorial University of Newfoundland occasional papers in biology, no 14
Carscadden J, Nakashima BS (1997) Abundance and changes in distribution, biology, and behavior of capelin in response to cooler waters of the 1990s. In: Forage fishes in marine ecosystems: proceedings of the international symposium on rose of forage fishes in Alaska. Sea Grant Program report no 97-01, Fairbanks, AL, pp 457–468
Carscadden JE, Vilhjalmsson H (2002) Capelin—what are they good for? ICES J Mar Sci 59:863–869
Clapham PJ, Baraff LS, Carlson CA, Christian MA, Mattila DK, Mayo CA, Murphy MA, Pittman S (1993) Seasonal occurrence and annual return of humpback whales, Megaptera novaeangliae, in the southern Gulf of Maine. Can J Zool 71:440–443
Cox SL, Scott BE, Camphuysen KCJ (2013) Combined spatial and tidal processes identify links between pelagic prey species and seabirds. Mar Ecol Prog Ser 479:203–221
Davoren GK (2007) The spatial and temporal overlap of a biological hotspot and fishing activities: negative human–wildlife interactions in the Northwest Atlantic. Conserv Biol 21:1032–1045
Davoren GK (2013) Divergent use of spawning habitat by male capelin (Mallotus villosus) in a warm and cold year. Behav Ecol 24:152–161. doi:10.1093/beheco/ars147
Davoren GK, Montevecchi WA (2003) Signals from seabirds indicate changing biology of capelin stocks. Mar Ecol Prog Ser 258:253–261
Davoren GK, Montevecchi WA, Anderson JT (2003) Search strategies of a pursuit-diving marine bird and the persistence of prey patches. Ecol Monogr 73:463–481
Davoren GK, Anderson JT, Montevecchi WA (2006) Shoal behavior and maturity relations of spawning capelin (Mallotus villosus) off Newfoundland: demersal spawning and diel vertical movement patterns. Can J Fish Aquat Sci 63:268–284
Davoren GK, Buren A, Burke C, May C, Penton P, Record N, Rose-Taylor C, Andrews D, Reinfort B, Anderson JT, Bell T, deYoung B, Garthe S, Koen-Alonso M, Montevecchi WA (2008) An ecosystem-based research program for capelin (Mallotus villosus) in the Northwest Atlantic: overview and results. J Northwest Atl Fish Sci 39:35–48
Davoren GK, Garthe S, Montevecchi WA, Benvenuti S (2010) Influence of diel prey behaviour, prey type and whales on the foraging activities of a marine avian predator in a low Arctic ecosystem. Mar Ecol Prog Ser 404:275–287
Davoren GK, Penton PM, Burke C, Montevecchi WA (2012) Water temperature and timing of capelin spawning determine seabird diets. ICES J Mar Sci 69:1234–1241. doi:10.1093/icesjms/fss032
Embling CB, Illian J, Armstrong E, van der Kooij J, Sharples J, Camphuysen KCJ, Scott BE (2012) Investigating fine-scale spatio-temporal predator-prey patterns in dynamic marine ecosystems: a functional data analysis approach. J Appl Ecol 49:481–492
ESRI (2012) ArcGIS help professional library. http://help.arcgis.com/en/arcgisdesktop/10.0/help/index.html#/. Accessed 1 June 2012
Frank KT, Leggett WC (1981) Prediction of egg development and mortality rates in capelin (Mallotus villosus) from meteorological, hydrographic, and biological factors. Can J Fish Aquat Sci 38:1327–1338
Fridgeirsson E (1976) Observations on spawning behaviour and embryonic development of the Icelandic capelin. Rit Fiskid 5:3–40
Garthe S, Benvenuti S, Montevecchi WA (2000) Pursuit-plunging by gannets (Morus bassanus) feeding on capelin (Mallotus villosus). Proc R Soc Lond B 267:1717–1722
Garthe S, Montevecchi WA, Davoren GK (2007) Flight destinations and foraging behavior of northern gannets preying on a small forage fish in a low Arctic ecosystem. Deep Sea Res Part II 54:311–320
Garthe S, Montevecchi WA, Davoren GK (2011) Inter-annual changes in prey fields trigger different foraging tactics in a large marine predator. Limnol Oceanogr 56:802–812
Gende SM, Sigler MF (2006) Persistence of forage fish ‘hot spots’ and its association with foraging Steller sea lions (Eumetopias jubatus) in southeast Alaska. Deep Sea Res Part II 53:432–441
Gill PC, Morrice MG, Page B, Pirzl R, Levings AH, Coyne M (2011) Blue whale habitat selection and within-season distribution in a regional upwelling system off southern Australia. Mar Ecol Prog Ser 421:243–263
Hamer KC, Phillips RA, Hill JK, Wanless S, Wood AG (2001) Contrasting foraging strategies of gannets Morus bassanus at two North Atlantic colonies: foraging trip duration and foraging area fidelity. Mar Ecol Prog Ser 224:283–290
Haney JC, Fristrup KM, Lee DS (1992) Geometry of visual recruitment by seabirds to ephemeral foraging flocks. Ornis Scand 23:49–62
Hedd A, Regular PM, Montevecchi WA, Buren AD, Burke CM, Fifield DA (2009) Going deep: common murres dive into frigid water for aggregated, persistent and slow-moving capelin. Mar Biol 156:741–751
Hedd A, Montevecchi WA, Otley H, Phillips RA, Fifield DA (2012) Trans-equatorial migration and habitat use by sooty shearwaters Puffinus griseus from the South Atlantic during the nonbreeding season. Mar Ecol Prog Ser 449:277–290
Hobson KA, Montevecchi WA (1991) Stable isotopic determinations of trophic relationships of great auks. Oecologia 87:528–531
Hooker SK, Gerber LR (2004) Marine reserves as a tool for ecosystem-based management: the potential importance of megafauna. Bioscience 54:27–39
Howell EA, Kobayashi DR, Parker DM, Balazs GH, Polovina A (2008) Turtlewatch: a tool to aid in the bycatch reduction of loggerhead turtles Caretta caretta in the Hawaii-based pelagic longline fishery. Endanger Species Res 5:267–278
Hunt GL Jr, Russel RW, Coyle KO, Weingartner T (1998) Comparative foraging ecology of planktivorous auklets in relation to ocean physics and prey availability. Mar Ecol Prog Ser 167:241–259
Hyrenbach KD, Forney KA, Dayton PK (2000) Marine protected areas and ocean basin management. Aquat Conserv 10:437–458
Irons DB (1998) Foraging area fidelity of individual seabirds in relation to tidal cycles and flock feeding. Ecology 79:647–655
Lewison R, Crowder L, Read A, Freeman S (2004) Understanding impacts of fisheries bycatch on marine megafauna. Trends Ecol Evol 19:598–604
Louzao M, Becares J, Rodríguez B, Hyrenbach DK, Ruiz A, Arcos JM (2009) Combining vessel-based surveys and tracking data to identify key marine areas for seabirds. Mar Ecol Prog Ser 391:183–197
Montevecchi WA (2007) Binary responses of northern gannets (Morus bassanus) to changing food web and oceanographic conditions. Mar Ecol Prog Ser 352:213–220
Montevecchi WA, Tuck LM (1987) Newfoundland birds: exploitation, study, conservation. Nuttall Ornithological Club, Cambridge, MA
Montevecchi WA, Benvenuti S, Garthe S, Davoren GK, Fifield D (2009) Flexible foraging tactics by a large opportunistic seabird preying on forage- and large pelagic fishes. Mar Ecol Prog Ser 385:295–306
Moulton VD, Mactavish BD (2004) Recommended seabird and marine mammal observational protocols for Atlantic Canada. LGL report SA775-1. Report from LGL Ltd., St. John’s, NL, for Environmental Studies Research Funds, Calgary, AB, p 71
Nur N, Jahncke J, Herzog MP, Howar J, Hyrenback D, Zamon JE, Ainley DG, Wiens JA, Morgan K, Ballance LT, Stralberg D (2011) Where the wild things are: predicting hotspots of seabird aggregations in the California Current system. Ecol Appl 21:2241–2257
Penton PM, Davoren GK (2012) Physical characteristics of persistent deep-water spawning sites of capelin: importance for delimiting critical marine habitats. Mar Biol Res 8:778–783
Petrie B, Akenhead SA, Lazier SA, Loder J (1988) The cold intermediate layer on the Labrador and northeast Newfoundland shelves, 1978–86. NAFO Sci Counc Stud 12:57–69
Piatt JF, Methven DA (1992) Threshold foraging behaviour of baleen whales. Mar Ecol Prog Ser 84:205–210
Regular P, Davoren GK, Hedd A, Montevecchi WA (2010) Crepuscular foraging by a pursuit-diving seabird: tactics of common murres in response to the diel vertical migration of capelin. Mar Ecol Prog Ser 415:295–304
Ronconi RA, Ryan PG, Ropert-Coudert Y (2010) Diving of great shearwaters (Puffinus gravis) in cold and warm water regions of the South Atlantic Ocean. PLoS One 5:e15508. doi:10.1371/journal.pone.0015508
Shaffer SA, Tremblay Y, Weimerskirch H, Scott D, Thompson DR, Sagar PM, Moller H, Taylor GA, Foley DG, Block BA, Costa DP (2006) Migratory shearwaters integrate oceanic resources across the Pacific Ocean in an endless summer. Proc Natl Acad Sci 103:12799–12802
Sigler MF, Kuletz KJ, Ressler PH, Friday NA, Wilson CD, Zerbini AN (2012) Marine predators and persistent prey in the southeast Bering Sea. Deep Sea Res Part II 65–70:292–303
Tasker ML, Hope Jones P, Dixon T, Blake BF (1984) Counting seabirds at sea from ships: a review of methods employed and a suggestion for a standardized approach. Auk 101:567–577
Tchoukanski I (2012) ET GeoWizards 10.2, ET spatial techniques. http://www.ian-ko.com/. Accessed 27 Oct 2012
Vilchis LI, Ballance LT, Fiedler PC (2006) Pelagic habitat of seabirds in the eastern tropical pacific: effects of foraging ecology on habitat selection. Mar Ecol Prog Ser 315:279–292
Weimerskirch H, Sagar PM (1996) Diving depths of sooty shearwaters Puffinus griseus. Ibis 138:786–788
Whitehead H (1983) Structure and stability of humpback whale groups off Newfoundland. Can J Zool 61:1391–1397
Whitehead H, Carscadden JE (1985) Predicting inshore whale abundance—whales and capelin off the Newfoundland coast. Can J Fish Aquat Sci 42:976–981
Worm B, Sandow M, Oschlies A, Lotze HK, Myers RA (2005) Global patterns of predator diversity in the open oceans. Science 309:1365–1369
Acknowledgments
Principal funding was provided by a Natural Science and Engineering Council of Canada postgraduate scholarship (2000–2001), postdoctoral fellowship (2002–2003), Discovery grants (2004–2006, 2007–2011), and University of Manitoba Research Grant (2009) to GKD. Ship time funding was provided by NSERC (2009–2011) and an International Polar Year grant (2007) for work aboard the Lady Easton II and by Fisheries and Oceans Canada for work aboard the Canadian Coast Guard vessel Shamook (2000–2003). I gratefully acknowledge A. D. Murphy for directing, operating, and managing all technical equipment and electronic data aboard the CCG Shamook. Thanks also to G. Leo for help with analysis, as well as C. Burke, P. Penton, B. B. Harter, E. Maxner, L. Hayhurst, J. Allen, K. Wild, and the captains and crews of the CCGS Shamook and Lady Easton II for assistance with field work.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by S. Garthe.
Rights and permissions
About this article
Cite this article
Davoren, G.K. Distribution of marine predator hotspots explained by persistent areas of prey. Mar Biol 160, 3043–3058 (2013). https://doi.org/10.1007/s00227-013-2294-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00227-013-2294-5