
Abstract
The survival of marine predators depends on behavioural plasticity to cope with changes in prey distribution. Variability in behaviour might predict plasticity and is easier to assess than plasticity. Using miniaturized GPS loggers over several breeding seasons in two Norwegian Northern gannet (Morus bassanus) colonies, we investigated if and how the variability within and between individuals, but also between colonies and years, affected foraging strategies. Results revealed strong individual variability (foraging trip durations, foraging effort and different foraging areas). Individuals from both colonies showed preferred commuting routes, flight bearings and feeding hotspots. Individuals from the largest colony used larger and more foraging areas than individuals from the small colony. Feeding hotspots and foraging ranges varied amongst years in the largest colony only. Our study demonstrated that gannets show flexibility by changing prey fields that are driven by shifting oceanographic conditions.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Ecosystems are consistently exposed to environmental variations where spatio-temporal modifications of abiotic conditions impact living organisms and challenge food-web stability. Indeed, primary productivity, which is strongly related to abiotic parameters, varies notably in time and space and affects both resource distribution and abundance and its accessibility to consumers (Coe et al. 1976; Olff et al. 2002; Townsend et al. 2003; Brown et al. 2010). In a context of global and long-lasting deterioration of ecosystems, behavioural plasticity of individuals plays a key role in a population’s ability to adapt to rapid environmental changes (Nussey et al. 2007). This is particularly true for long-lived species whose microevolutionary responses might be too slow (Wingfield 2003). The ability to handle food shortages depends on the variety of individual foraging tactics in response to multi-scale spatio-temporal changes in prey distribution. To determine whether long-lived species, such as marine predators, can buffer variations in food abundance and location (e.g. distance from the breeding colony), their behavioural plasticity should be tested. However, such testing is difficult in wild populations, mainly because of the difficulty of assessing simultaneously variations in the availability of prey and in predators’ foraging behaviours. Nevertheless, we can measure individual flexibility in foraging behaviour and differences (thus variability) showed by colonies in the same and subsequent years. The lack of such flexibility/variability can be interpreted as either there are no environmental changes influencing prey availability, or the individuals lack plasticity in their behaviour to compensate for environmental changes.
We evaluated the foraging variability in the Northern gannet (Morus bassanus, hereafter named ‘gannet’), a central place forager constrained during the breeding period to return to the colony to feed its single chick at short intervals. Gannets are large seabirds able to travel hundreds of kilometres from their breeding site, which feed on a broad spectrum of prey species and size (Montevecchi and Barrett 1987; Hamer et al. 2000). This mobility, and a broad diet, enables gannets to buffer environmental variability (Montevecchi et al. 2009), and previous studies have highlighted their ability to face inter-annual variations in prey availability (Garthe et al. 2011), consistency in foraging behaviour and foraging area fidelity at the intra-individual level (Hamer et al. 2007), but also contrasting foraging tactics between colonies (Garthe et al. 2006; Hamer et al. 2001). To our knowledge, however, no multi-scale analysis of gannet foraging variability has yet been made. We therefore explored how variability in gannet foraging responses is partitioned at both the intra- and inter-individual levels, as well as the importance of inter-annual and inter-colonial variations. In the light of the rapid recovery of the gannet populations and the rapid expansion of their distribution area with the establishment of new colonies over the last century (Nelson 2002), we hypothesized that variability in their foraging behaviour is largely expressed at an individual level. Individuals may develop foraging flexibility to face environmental variations and changes in food distribution. Additional variability may, however, emerge from groups of individuals (i.e. a colony), thereby extending the variety of foraging patterns. We predicted that any variability in foraging behaviour will thus occur at both the intra- and the inter-individual levels, as well as at and the inter-colony level, and that variability might depend on yearly environmental conditions and changing prey fields (Garthe et al. 2007, 2011). To investigate such foraging variability in gannets, we analysed GPS tracks of successive foraging trips for birds from two distant colonies during 3 consecutive years.
Materials and methods
The fieldwork was conducted in North Norway in July 2008 and 2009, at Store Ulvøyholmen (68°51′N, 14°51′E), Vesterålen and in July 2007, 2008 and 2009 at Storstappen (71°14′N, 25°30′E), near the North Cape (Fig. 1). Authorizations to enter the colonies were granted by the County Governors of Nordland and Finnmark, while permission to handle the birds was given by the Norwegian Directorate for Nature Management and the Norwegian Animal Research Authority.

Localization of the two studied colonies along the Norwegian coast: Store Ulvøyholmen and Storstappen
GPS deployments
We deployed GPS data loggers on breeding adults rearing 2–5-week-old chicks caught in the colonies using a noose pole. Birds were caught when both parents were present at the nest, and we selected the departing bird in the pair identified as the one adopting the ‘sky pointing’ posture (Ropert-Coudert et al. 2004a, b). Handling and attachment of the GPS data logger lasted <10 min in all cases. The bird’s head was covered to reduce stress. The devices were attached to the lower back of the birds using white Tesa tape (Hamburg, Germany) to match plumage colour. These techniques have been used in several previous studies without noticeable impact on the birds (Garthe et al. 2007; Grémillet et al. 2006; Hamer et al. 2009; Pichegru et al. 2010). In 2007 and 2008, devices were deployed for maximum 48 h and recorded at least one foraging trip. In 2009, devices were deployed for 5–6 days, recording at least three consecutive foraging trips for each bird. All birds returned to their nest after the deployment and continued to breed. We caught birds haphazardly to avoid any bias in sex ratio (Lewis et al. 2002) and within the outermost 10-m fringe of the colony. Several birds caught at both colonies had been ringed as chicks, indicating that old birds also bred in the area accessible with the pole (mean age of ringed birds in both colonies (N = 25) was 16.3 years old—eight birds were more than 20 years old). We therefore assumed that working on the fringe of the colony where one might expect to find young/inexperienced birds (Nelson 2002) did not bias the age structure of our sample.
GPS specifications and data filtering
Two models of GPS data loggers from Technosmart (Rome, Italy) were deployed after waterproofing in heat-shrink tubing. In 2007 and 2008, GPSs ‘Gipsy 1’ (95 × 48 × 24 mm; 65 g, i.e. 2 % of adult body mass) were programmed to record positions every 10 s for a maximum duration of 48 h. In 2009, GPSs ‘Gipsy 2’ (110 × 45 × 25 mm; 50 g, i.e. 1.6 % of adult body mass) were set to record every 10 s over 5 consecutive days. Successive trips recorded in 2009 were used to describe repeatability at the individual level. Using direct observations, we noted that trips of <30 min were of birds bringing back nest material and not food, and they were excluded from the analysis. For each trip, we calculated path length (adding all the distances between two consecutive GPS fixes), the maximum distance from the nest reached at sea, the time spent flying (i.e. when speed >10 km h−1) and a path sinuosity index to identify fishing events and locate feeding locations (see Grémillet et al. 2004, 2006 for details and validation). All GPS devices have been tested before deployment to validate accuracy of provided speed values (following Grémillet et al. 2004).
Statistical analyses of foraging path characteristics
Statistical analyses were performed using R 2.11.1 (http://www.r-project.org). First, we addressed the level of foraging variability at the intra- and inter-individual levels and the effect of spatio-temporal factors. A global analysis of variance was conducted using a comparison of generalized linear mixed models (GLMM) with year and colony as fixed effects and bird identity as random effect associated to an autocorrelation structure of order 1, to correct for potential pseudoreplication. A principal component analysis (PCA) was performed to identify correlations between the 6 descriptive variables of the foraging trips: trip duration, total path length, maximum distance to the nest, speed, flying time and sinuosity. Candidate models were run on the two first components of the PCA separately, and we compared them two by two (Zuur et al. 2009) to select the best model. For each comparison, a one-way ANOVA (fixed effects) and LRT method (random effect) were used to determine the model best supported by the dataset. For each component of the PCA (Dim.1 and Dim.2), model selection highlighted the mechanisms and the factors influencing variability in foraging behaviour.
In a second phase, we analysed variability of trip duration, path length, maximum distance and speed with GLMMs (year as fixed effect, identity as random effect associated to an autocorrelation structure of order 1) within each colony. The GLMM was compared with a null model (without year as fixed effect) using a one-way ANOVA. Inter-annual variability was tested using a one-way ANOVA and post hoc Tukey tests.
Spatial analyses
To identify and describe the use of the particular feeding areas by gannets and to understand whether they respond to changes in prey distribution by varying their behaviour, foraging trips and feeding positions were plotted and analysed using Arcgis 9.2 (ESRI, France) and Hawth’s Tools 3.27 extension. We used Minimum Convex Polygons (100 %) to define the visited area for individual trips. The Kernel Density Estimate tool was used to transform feeding positions into density estimates (Wood et al. 2000) and to determine the most intensively exploited areas within the visited area of each colony.
-
(a)
Intra-individual variability
We analysed individual variability using three complementary parameters measured on successive trips in 2009.
Visited area fidelity
For every bird, we calculated a cumulative visited area for each additional trip to determine if or how the prospected area increased over 5 days of recording. To assess repeatability at an intra-individual scale, we calculated the overlap percentage of each pair of consecutive trips and the overlap percentage of all trips. To do so, we pooled spatially the two surfaces (not the sum of areas values), and calculated the percentage overlap with respect to the pooled surface. For each colony and each bird, we compared the ‘first trip’ visited area with the overall ‘5-days’ visited area calculated from all trips over the 5 days.
Bearing fidelity
We measured the bearings followed by birds when they left and returned to the nest to examine bearing consistency across successive trips. Since tracks were not all linear, we measured the azimuth on the first linear part at a minimum distance of 3 km from the colony. Using circular statistics for paired samples (Zar 1998), we calculated for all pairs of two successive trips (1) the difference between the azimuth of the first outbound trip (O1) and the azimuth of the second outbound trip (O2) and (2) the difference between the azimuth of the first inbound (I1) trip and the azimuth of the second outbound trip (O2). A mean difference of angle was calculated for each bird. We then calculated differences of angles between foraging trips recorded from different individuals (outbound trip from bird 1 vs. bird 2, etc.). Using two sample t tests, we compared differences in angles from two successive trips of a single bird with differences in angles between two unrelated trips. If repeatability in bearing is high, then the difference in angles should be lower for successive trips of the same bird than for unrelated trips.
Feeding area fidelity
We determined for each bird the total number of distinct feeding areas (feeding events separated by a distance of 5 km) and the total number of feeding events. We also estimated the mean number of repeated visits to the same feeding areas.
-
(b)
Inter-individual and inter-colony variability
Inter-individual variability in visited area was examined for each colony via an analysis of preferred directions, feeding locations and visited area overlap amongst individuals of the same colony. The cumulative visited area for each additional bird was calculated for each colony.
-
(c)
Inter-annual variability
Kernel density estimates of an entire colony were used as proxies of inter-annual variability in feeding hotspot distributions. We did not, however, find a method to estimate overlapping percentage between 2 years using kernels results. To estimate this overlap, we thus used Minimum Convex Polygons (MCP 100 %) to calculate the total area visited during the first trip of each bird. We then compared the percentage overlap between areas visited on two consecutive years for each colony. We are aware that MCP introduces a bias (i.e. overestimation), but this bias is shared by years and the result gives a comparable measure of overlap.
Results
Sample sizes
We recorded a total of 341 foraging trips performed by 101 birds (N = 21, 23 and 14 on Storstappen in 2007, 2008 and 2009, respectively; N = 23 and 20 on Store Ulvøyholmen in 2008 and 2009, respectively). Each bird performed 1–14 foraging trips and we recorded a minimum of 3 consecutive trips for 39 birds. Because several analyses are informative on multiple scales, results are given for each type of analysis. The four considered scales (intra-individual, inter-individual, inter-annual and inter-colony) will be addressed in the discussion. Values are presented as mean ± SE.
Global variance analysis
The PCA analysis revealed a strong positive correlation between total trip duration, path length, maximum distance to the nest and flying time, and a negative correlation between speed and sinuosity. The two first components of the PCA explained 77.2 % of the variance. The first component ‘Dim.1’ aggregated four of the six variables in relation to the duration and distance to the nest during the foraging trip (i.e. total trip duration, path length, maximum distance to the nest and flying time) and explained 55.8 % of the variance. The second component ‘Dim.2’ (flight speed negatively correlated to sinuosity of the path) explained 21.4 % of the variance.
From the GLMMs (Table 1), we selected the following models:
Year and Colony had a significant effect on the observed variability of Dim.1, as well as inter-individual variability (expressed in the model by the random effect ‘identity’). Intra-individual variability was high, implying a weak correlation between two successive trips (autocorrelation factor ϕ = 0.06). Thus, the correlation structure did not improve the model significantly. Year and Identity had a significant effect on the observed variability of Dim.2. The correlation structure estimate (ϕ = 0.55) indicated a positive autocorrelation in speed and sinuosity of foraging. The Colony did not influence variability of Dim.2. We could not test any interaction effect between Year and Colony on the foraging behaviour, because no data were available in 2007 for Store Ulvøyholmen. To examine the foraging variability in detail, we pursued foraging behaviour analysis within colonies.
Spatial analyses
-
(a)
Intra- and inter-individual variability
At Store Ulvøyholmen, trip durations varied from 30 min to 22.8 h, with maximum distances from the nest of 3–98 km and total path lengths from 18 to 360 km, all years together. At Storstappen, foraging trip durations varied from 30 min to 23.5 h, with maximum distances from the nest ranging from 5 to 132 km and total path lengths from 18 to 595 km, all years together.
Visited area fidelity
Individual visited areas increased when new trips were recorded. On Store Ulvøyholmen, the size of most individual visited areas reached a plateau beyond four trips. On Storstappen, gannets seemed to prospect new areas on each new foraging trip, although the size of individual visited areas started to reach a plateau beyond seven trips (Fig. 2). On Store Ulvøyholmen, the overlap between two successive trips of a single bird varied between 0 and 96 %, with a mean of 27 %. The overlap for all trips of a single bird varied between 5 and 56 %, with a mean of 27 %. On Storstappen, the overlap between two successive trips varied between 0 and 76 %, with a mean of 11 %. The overlap for all trips of a single bird varied between 1 and 55 %, with a mean of 12 %. At Store Ulvøyholmen, the visited area increased from 1,052 to 1,796 km2 if the first trip of each bird and all trips are considered (‘first trip’ vs. ‘5-days’ visited areas, Table 2; Fig. 3a). In the same way, the ‘first trip’ visited area was 9,672 km2 and the ‘5-days’ visited area was 13,856 km2 at Storstappen (Table 2; Fig. 3b). Cumulative visited areas (calculated for both ‘first trip’ and ‘5-days’) increased in the relation with the number of birds, but the visited area size stopped increasing beyond six and seven birds at Store Ulvøyholmen and Storstappen, respectively (Fig. 4).

Cumulative visited area per individual gannet in relation with the number of foraging trips. Each solid line represents a single bird

Visited areas calculated from the first trip (solid line) and all trips over a 5-day period (dashed line). a Store Ulvøyholmen (N = 12) and b Storstappen (N = 14)

Cumulative visited area per gannet colony in relation with the number of equipped birds and duration of GPS deployment. 5-day deployment in black dots, 1-day deployment in gray triangles
Bearing fidelity
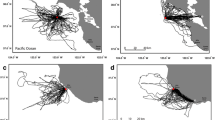
Differences in azimuths from successive trips were similar to azimuths from unrelated trips at Store Ulvøyholmen (p = 0.22 for O1 vs. O2; p = 0.70 for I1 vs. O2). At Storstappen, differences in azimuth between two unrelated trips were lower than the difference of azimuth between 2 successive trips (p = 0.01 for O1 vs. O2; p = 0.04 for I1 vs. O2). At the inter-individual level, a wide range of bearings was used, but three main routes were preferentially followed by gannets in both colonies (Fig. 5).

Main foraging routes followed by gannets in 2007 (dotted line), in 2008 (dashed line and in 2009 (solid line in a Store Ulvøyholmen and b Storstappen
Feeding area fidelity
Gannets from Store Ulvøyholmen performed a mean of 6.3 ± 1.7 trips during 5 days of recording. Each bird foraged in average 8.8 ± 2.3 times in 3.9 ± 1.2 distinct areas. Each feeding area was visited at least twice by the same individual during the 5-day period. On Storstappen, each gannet performed an average of 8.3 ± 2.8 trips during 5 days of recording, and foraged 18.3 ± 5.2 times in 8.3 ± 2.1 distinct areas. Each feeding area was visited at least twice during the 5 days.
At the inter-individual level, kernel density analyses revealed that most of the gannets from Store Ulvøyholmen fed close to the colony, either inshore off Hadseløya or along the north coast of Lofoten (Fig. 6a). At Storstappen, feeding hotspots were northwest of the colony, north off the North Cape and in the fjord south off the colony (Fig. 6b).

Relative density of gannet feeding locations around the breeding sites in 2007, 2008 and 2009, from kernel density estimate analysis, Jenks Natural thresholds in a Store Ulvøyholmen (N = 23 and 20 in 2008 and 2009) and b Storstappen (N = 21, 23 and 14 in 2007, 2008 and 2009)
-
(b)
Inter-annual variability
At Store Ulvøyholmen, we found no significant difference between years in the speed of flight, trip duration and the maximum distance to nest between years (ANOVA: duration p = 0.31, max. distance p = 0.17, speed p = 0.11, Table 2) but path lengths were longer in 2008 than in 2009 (Tukey: p = 0.05, Table 2). At Storstappen, trip duration, path length, maximum distance to the nest and flight speed differed significantly between years (ANOVA: duration p = 0.003, path length p < 0.001, max distance p < 0.001 and speed p = 0.034). Trip duration was shorter in 2007 than in 2008 (Tukey: p < 0.001, Table 2) and in 2009 (Tukey: p = 0.02, Table 2). Maximum distance was significantly lower in 2007 than in 2008 (Tukey: p < 0.001, Table 2) and in 2009 (Tukey: p < 0.001, Table 2). Total path length was also significantly different between all years (Tukey: 2007–2008 p < 0.001; 2007–2009 p = 0.001; 2008–2009 p = 0.01, Table 2). Flight speed only differed between 2007 and 2008 (Tukey: p = 0.005, Table 2).
During the different breeding seasons of the study, the gannets from both sites showed preferential routes (Fig. 5), but there were higher inter-annual variations from Storstappen.
Gannets from Store Ulvøyholmen travelled in the same directions both years (Fig. 5a) and, although feeding locations were very similar, they were more tightly clustered in 2009 than in 2008 (Fig. 6a). Gannets from Storstappen foraged within 20 km of the coast in 2007, whereas in 2008, they travelled further offshore in two main directions 50–100 km from the colony. In 2009, they foraged 40–80 km from the colony, following the same 3 main routes as in 2007 (Fig. 5b); though, several birds also moved round the North Cape and southwards to Kamøyfjord and Porsangerfjord. Kernel density estimates showed important variations in feeding locations, although several feeding hotspots were consistent throughout the study period (Fig. 6b).
-
(c)
Colony visited area
We calculated the annual home range for each colony as a Minimum Convex Polygon (100 %) based on the first foraging trip of each bird. At Store Ulvøyholmen, visited areas in 2008 and 2009 were of similar size (Table 2) and overlapped by 70 % of the total area (Fig. 7a). At Storstappen, areas visited in 2007 were smaller than in 2008 and 2009 (Table 2). Visited areas overlapped by 34 % of the total area between 2007 and 2008 and by 75 % of the total area between 2008 and 2009 (Fig. 7b).

Annual visited areas in 2007 (hatched line), 2008 (dotted line) and 2009 (solid line), made from the sum of the first foraging trips of each bird in a Store Ulvøyholmen (N = 23 and 20 in 2008 and 2009) and b Storstappen (N = 21, 23 and 14 in 2007, 2008 and 2009)
Discussion
In this study, we analysed components of variability in foraging behaviour of Northern gannets within individuals and between colonies and years. Northern gannets are known to be flexible predators, exploiting a large panel of prey (Montevecchi and Barrett 1987) and responding rapidly to changes in prey distribution by increasing their foraging effort (Garthe et al. 2007; Hamer et al. 2007) or consuming alternative prey (Garthe et al. 2011; Montevecchi and Myers 1997), including fishery discards (Votier et al. 2010). To our knowledge, however, no study encompassed all levels of variability concurrently. Improvements in animal-borne GPS technology allowed us to successfully obtain a reasonable sample of successive foraging trips within individuals. Here, we showed that gannets exhibit high intra-individual flexibility, as well as different foraging tactics at the inter-individual level, which may facilitate their adaptation to variations in prey conditions in space and time. Variability in a top predator’s foraging tactics is the condition sine qua non for behavioural plasticity. Plasticity plays a key role in the ability of organisms to adjust to environmental variability. Understanding mechanisms and limits of animals’ flexibility is crucial for predicting population viability (Komers 1997).
Individual flexibility
Intra-individual flexibility in gannets was illustrated by the great variability in the quantitative and spatial features of their foraging trips. An individual may perform long and short foraging trips successively but its flight speed and path sinuosity remain consistent. The low percentage of overlap in foraging areas between successive trips of the same individual suggests that birds can prospect different areas over a short period of time, thus cumulating individual knowledge of their environment over successive trips. As a consequence, the area explored by the colony as a whole was larger when considering the foraging trips of all birds over several (in our study five) days than when considering only one trip (i.e. the first recorded) per bird.
On the other hand, some birds showed repeatability in foraging tactics, returning on average twice (up to 5 times) to the same feeding area before heading in a different direction on a subsequent foraging trip. Analyses of flight compass bearings revealed that birds switched regularly between them, strongly suggesting memory-based foraging tactics (Davoren et al. 2003a; Montevecchi et al. 2009). When birds were returning in the same area, foraging paths appeared to be very similar, suggesting that prey-patch position may be anticipated by birds (Pettex et al. 2010). Gannets might thus use experience gained during the course of the breeding season to return to profitable areas (Grémillet et al. 2004; Hamer et al. 2007), as shown in other seabird species such as black-legged kittiwakes Rissa tridactyla (Irons 1998) and great cormorant Phalacrocorax carbo (Grémillet et al. 1999) or in other marine top predators such as southern elephant seals Mirounga leonina (Bradshaw et al. 2004) and Antarctic fur seals Arctocephalus gazella (Bonadonna et al. 2001).
A previous study using Platform Transmitter Terminals (PTTs) on gannets showed marked differences between two colonies in individual fidelity to foraging areas (Hamer et al. 2001). At one colony in the North Sea (Bass Rock), birds followed very similar bearings on successive trips, while bearings followed by birds from a colony in the Celtic Sea (Great Saltee) were much less consistent. Our results from precise foraging positions recorded by GPS devices corroborate such findings, showing that individual gannets from the same colony can be both consistent and flexible in the routes followed, in the areas exploited and in the durations and distances of foraging trips. How gannets decide to switch from a foraging area to another is unknown, but public information may play a role in the choice of compass bearing when departing from the colony (Ward and Zahavi 1973; Weimerskirch et al. 2010) although the relevance of this theory for vertebrates is still widely discussed because of a lack of conclusive experimental studies (Richner and Heeb 1995). For example, gannets from Bass Rock were very consistent in the bearings of successive flights but the distance travelled varied amongst trips of similar bearing (Hamer et al. 2001). The authors therefore suggested that gannets were heading to a known area and may use local information from conspecifics at sea to shorten or extend their search. Although local enhancement at sea may also occur, in our case, variations in bearing at the departure from the colony between successive trips might be better explained by the use by individuals of information provided by flocks of gannets returning from the sea, or memory-based mechanisms.
Inter-individual variability
The GLMM analysis highlighted an important inter-individual variability in foraging effort (Table 1). In addition, we recorded a wide range of trip durations (from 30 min to 23 h in both sites) and maximum distances from the nest (from <5 to >100 km). The bearing consistency also differed amongst individuals, from birds switching constantly between exploited foraging areas in all directions to birds following the same routes each time. Decision-making could be related to the success of previous foraging trips and a decrease of prey in patches or an avoidance of intra-specific competition. Prey depletion experiments in bumblebees Bombus impatiens have highlighted that individuals rapidly respond to an unsuccessful foraging bout by shifting foraging areas, and by returning to known profitable areas, even of lower quality (Townsend-Mehler and Dyer 2012). Similar experiments in great tits Parus major revealed marked differences in performance between individuals in their ability to find a more profitable site after resource depletion (Smith and Sweatman 1974). Contrasting site fidelity between gannets might reflect differences in foraging flexibility amongst individuals.
Our results showed that gannets prospected the whole potential feeding area around their colony. Cumulative visited areas revealed an inter-individual variability, but spatial inter-individual variability remained relatively low compared with individual flexibility. Recording several trips of a limited number of birds was sufficient to depict the whole area visited by birds from the colony. Our results suggested that to define the ‘visited area’ of a given colony in a given breeding period, it may be more informative to track successive trips of a small number of individuals rather than just one trip of a large number of individuals. There was considerable consistency amongst colonies in the choice of routes and feeding areas. Gannets from both colonies preferentially followed three directions and there was an annual occurrence of feeding hotspots at both sites. Although predictability of prey is higher at large spatial scales, habitats with specific features such as shelf edges and frontal zones benefit from an enhanced productivity and providing relatively stable prey concentrations for predators (Weimerskirch 2007). The feeding grounds along the North Norwegian coast are very productive due to the convergence of the low-saline Norwegian Coastal Current and the warm-saline North Atlantic Currents close to land off the Vesterålen and the North Cape area (Barrett et al. 2006). Food samples collected from regurgitates around the colony and from the handled birds were mainly composed of large and energy-rich prey at both sites: 25–30 cm herring Clupea harengus and mackerel Scomber scombrus at Storstappen, and herring, mackerel and saithe Pollachius virens at Store Ulvøyholmen (Pettex et al. unpublished data). Marked colony-preferred routes and feeding sites might thus reflect a good predictability of prey at a small spatio-temporal scale in both colonies (Davoren et al. 2003a). This is also concordant with the short foraging ranges observed at both sites, considering the gannets’ ability to travel great distances to find prey (Hamer et al. 2007).
Inter-colony variability
Preferred routes and feeding hotspots occurred at both sites, with birds from Storstappen exploiting feeding areas twice as large as those from Store Ulvøyholmen. The degree of overlap for two successive trips was consistently much lower at Storstappen (11 %) than at Store Ulvøyholmen (27 %).
In 2008 and 2009, gannets from Storstappen explored and foraged an area 8 times larger, made longer foraging trips and travelled further than gannets from Store Ulvøyholmen. This is consistent with the hypothesis that foraging effort and visited area are positively correlated to the size of the colony (Lewis et al. 2001) with Store Ulvøyholmen and Storstappen holding 308 and 1,244 breeding pairs in 2008, respectively (Barrett 2008). One might thus expect gannets from Storstappen to put in a greater foraging effort than those from Store Ulvøyholmen due to possible intra-specific competition for food. The duration of foraging trips did not, however, differ between the two sites, suggesting that (a) there was no such competition or (b) the gannets from Store Ulvøyholmen may have rested for longer periods on the sea surface during their trips (Ropert-Coudert et al. 2004a, b) than their conspecifics from Storstappen. Whereas gannets from both colonies might experience good environmental conditions that limit competition, differences in travelled distances and visited areas may also reflect differences in the distribution and availability of prey around the two study colonies. Variability in foraging behaviour in relation to contrasting environmental conditions between breeding sites has already been documented in gannets (Garthe et al. 2006; Hamer et al. 2001) and amongst other marine predators, such as gentoo penguin Pygoscelis papua (Lescroel and Bost 2005) and Northern fur seals Callorhinus ursinus (Robson et al. 2004). Whereas further work on more sites would be necessary to reach any firm conclusion, birds from the two breeding sites in this study behaved differently.
Inter-annual variability
Changes in food distribution can affect predator foraging behaviour, and animals should adapt their foraging effort or exploit different feeding grounds to find their prey (McCafferty et al. 1998; Hamer et al. 2007). Environmental conditions near Store Ulvøyholmen were probably similar in 2008 and 2009, as gannets showed a very similar foraging behaviour. Visited areas were of identical size (ca 1,000 km2) and overlapped widely. Gannets had marked preferred areas and foraged close to the colony, suggesting conditions of favourable and stable prey availability during our study. Conversely, gannets from Storstappen increased their foraging effort (trip duration, maximum distance to the nest and distance travelled) and their visited area significantly between 2007 and 2009 (Table 2). Although several areas were consistently exploited during the three breeding seasons, the general distribution of feeding hotspots varied between years. For instance, in 2008, birds from Storstappen foraged intensely in two areas to the north and northwest of the colony (ca 80 km), while in 2009, most of the fishing activity was limited to a semicircle 40 km from the breeding site and towards Porsangerfjord. This resulted in important variations in the overlap of the colony visited areas between 2007 and the two subsequent years. These changes in foraging effort and feeding areas in gannets may be a response to variations in distribution and/or availability of their prey. Although the Barents Sea holds amongst the most productive grounds in the North Atlantic (Sakshaug et al. 2009), previous studies have highlighted pronounced interannual variations of its productivity and biomass in relation to the North Atlantic Oscillation (Drinkwater 2011). These changes resulted in important variation in the distribution of pelagic fish such as capelin Mallotus villosus and cod Gadus morhua (Loeng and Drinkwater Loeng and Drinkwater 2007), but also herring (Røttingen 1990), an important prey for Storstappen gannets (Montevecchi and Barrett 1987, personal observation).
Ecological, conservation and methodological implications of flexibility in foraging
Our study showed that gannet foraging behaviour varies at the four considered levels and that all derive from individual flexibility. Individual flexibility, and as a consequence inter-individual variability, may give to gannets an advantageous ability to cope with environmental changes. The world population of Northern gannets dramatically declined across the nineteenth century because of a massive exploitation of adults, eggs and chicks (Wanless et al. 2005), but gannets positively and rapidly responded to protection measures during the twentieth century, recovering and recolonizing a large part of their distribution area (Montevecchi and Hufthammer 1990; Wanless et al. 2005). The rapid recovery of gannets over the last decades might be related to their highly flexible foraging behaviour (Garthe et al. 2007; Hamer et al. 2001, 2007) and broad diet spectrum (Montevecchi et al. 2009). Young colonies often experience a long latency period in their development, during which only a small number of pioneers settles, before becoming attractive for other individuals and growing fast (Montevecchi et al. 1987; Moss et al. 2002). Pioneering individuals will not have the benefit of many conspecifics to attend efficiently all profitable feeding areas (Forbes and Kaiser 1994) either via local enhancement (Davoren et al. 2003b) or public information (Ward and Zahavi 1973). Instead, they must rely upon individual flexibility to cope with the unpredictable marine environment. Thus, the foraging flexibility of settlers might be essential for an efficient exploitation of resources and to ensure the durability of the colony during the early stage of its development. It is, however, essential to assess the magnitude of foraging flexibility across the year cycle (e.g. during and outside the breeding season), as well as for different life stages (fledglings and juveniles, breeding and non-breeding individuals, see Votier et al. 2011), to attain a complete picture of behaviours that fuel plasticity in marine top predators such as gannets.
Our results also showed that the distribution of foraging birds might differ amongst years. This must be considered in the future when developing conservation measures intended to protect marine predators. Marine protected areas (MPA) are increasingly implemented to protect pelagic habitats (Wood et al. 2008), but the mobility of targeted species is a challenge to define suitable limits for them (Game et al. 2009). Bio-logging technologies have greatly improved our knowledge of the spatial ecology of marine species (Ryan et al. 2004; Staniland et al. 2004; Piatt et al. 2006; Burger and Shaffer 2008), and concurrently, the dynamic character of marine ecosystems needs to be considered by providing multi-year data to conservation managers (Wilson et al. 2009). Multi-site studies might also be necessary to faithfully describe foraging behaviour on a regional scale and acquire a reliable overview of animal behaviour. These constraints should be taken into account in future studies and management decisions to avoid a mismatch between the complexity of animal behaviour and conservation objectives.
References
Barrett RT (2008) Recent establishments and extinctions of Northern Gannet Morus bassanus colonies in North Norway, 1995–2008. Ornis Norvegica 31:172–182
Barrett RT, Lorentsen SH, Anker-Nilssen T (2006) The status of seabirds breeding in mainland Norway. Atlantic Seabirds 8(3):97–126
Bonadonna F, Lea M, Dehorter O, Guinet C (2001) Foraging ground fidelity and route-choice tactics of a marine predator: the Antarctic fur seal Arctocephalus gazella. Mar Ecol Prog Ser 223:287–297. doi:10.3354/meps223287
Bradshaw CJA, Hindell MA, Sumner MD, Michael KJ (2004) Loyalty pays: potential life history consequences of fidelity to marine foraging regions by southern elephant seals. Anim Behav 68:1349–1360. doi:10.1016/j.anbehav.2003.12.013
Brown CJ, Fulton EA, Hobday AJ et al (2010) Effects of climate-driven primary production change on marine food webs: implications for fisheries and conservation. Glob Change Biol 16:1194–1212. doi:10.1111/j.1365-2486.2009.02046.x
Burger AE, Shaffer SA (2008) Application of tracking and data-logging technology in research and conservation of seabirds. Auk 125:253–264. doi:10.1525/auk.2008.1408
Coe MJ, Cumming DH, Phillipson J (1976) Biomass and production of large African herbivores in relation to rainfall and primary production. Oecologia 22:341–354. doi:10.1007/BF00345312
Davoren GK, Montevecchi WA, Anderson JT (2003a) The influence of fish behaviour on search strategies of common murres Uria aalge in the Northwest Atlantic. Marine Ornithology 31:121–131
Davoren GK, Montevecchi WA, Anderson JT (2003b) Search strategies of a pursuit-diving marine bird and the persistence of prey patches. Ecol Monogr 73:463–481. doi:10.1890/02-0208
Drinkwater KF (2011) The influence of climate variability and change on the ecosystems of the Barents Sea and adjacent waters: review and synthesis of recent studies from the NESSAS project. Progress in oceanography in press, accepted Manuscript. doi:10.1016/j.pocean.2011.02.006
Forbes LS, Kaiser GW (1994) Habitat choice in breeding seabirds: when to cross the information barrier. Oikos 70:377–384
Game ET, Grantham HS, Hobday AJ et al (2009) Pelagic protected areas: the missing dimension in ocean conservation. Trends Ecol Evol 24:360–369. doi:10.1016/j.tree.2009.01.011
Garthe S, Montevecchi WA, Chapdelaine G et al (2006) Contrasting foraging tactics by Northern gannets (Sula bassana) breeding in different oceanographic domains with different prey fields. Mar Biol 151:687–694. doi:10.1007/s00227-006-0523-x
Garthe S, Montevecchi WA, Davoren GK (2007) Flight destinations and foraging behaviour of Northern gannets (Sula bassana) preying on a small forage fish in a low-Arctic ecosystem. Deep Sea Res II 54:311–320. doi:10.1016/j.dsr2.2006.11.008
Garthe S, Montevecchi WA, Davoren GK (2011) Inter-annual changes in prey fields trigger different foraging tactics in a large marine predator. Limnol Oceanogr 56:802–812. doi:10.4319/lo.2011.56.3.0802
Grémillet D, Wilson RP, Storch S, Gary Y (1999) Three-dimensional space utilization by a marine predator. Mar Ecol Prog Ser 183:263–273
Grémillet D, Dell’Omo G, Ryan PG et al (2004) Offshore diplomacy, or how seabirds mitigate intra-specific competition: a case study based on GPS tracking of Cape gannets from neighbouring colonies. Mar Ecol Prog Ser 268:265–279
Grémillet D, Pichegru L, Siorat F, Georges JY (2006) Conservation implications of the apparent mismatch between population dynamics and foraging effort in French Northern gannets from the English Channel. Mar Ecol Prog Ser 319:15–25
H Loeng, K Drinkwater (2007) An overview of the ecosystems of the Barents and Norwegian Seas and their response to climate variability. Deep Sea Res II 54:2478–2500. doi:10.1016/j.dsr2.2007.08.013
Hamer KC, Phillips RA, Wanless S, Harris MP, Wood AG (2000) Foraging ranges, diets and feeding locations of gannets Morus bassanus in the North Sea: evidence from satellite telemetry. Mar Ecol Prog Ser 200:257–264
Hamer KC, Phillips RA, Hill JK et al (2001) Contrasting foraging strategies of gannets Morus bassanus at two North Atlantic colonies: foraging trip duration and foraging area fidelity. Mar Ecol Prog Ser 224:283–290
Hamer KC, Humphreys EM, Garthe S et al (2007) Annual variation in diets, feeding locations and foraging behaviour of gannets in the North Sea: flexibility, consistency and constraint. Mar Ecol Prog Ser 338:295–305
Hamer KC, Humphreys EM, Magalhães MC et al (2009) Fine-scale for aging behaviour of a medium-ranging marine predator. J Anim Ecol 78:880–889. doi:10.1111/j.1365-2656.2009.01549.x
Irons DB (1998) Foraging area fidelity of individual seabirds in relation to tidal cycles and flock feeding. Ecology 79:647–655. doi:10.2307/176960
Komers PE (1997) Behavioural plasticity in variable environments. Can J Zool 75:161–169. doi:10.1139/z97-023
Lescroel A, Bost C (2005) Foraging under contrasting oceanographic conditions: the gentoo penguin at Kerguelen Archipelago. Mar Ecol Prog Ser 302:245–261
Lewis S, Sherratt TN, Hamer KC, Wanless S (2001) Evidence of intra-specific competition for food in a pelagic seabird. Nature 412:816–819. doi:10.1038/35090566
Lewis S, Benvenuti S, Dall’Antonia L et al (2002) Sex-specific foraging behaviour in a monomorphic seabird. Proc R S Lond B Biol Sci 269:1687–1693
McCafferty DJ, Boyd IL, Walker TR, Taylor RI (1998) Foraging responses of Antarctic fur seals to changes in the marine environment. Mar Ecol Prog Ser 166:285–299
Montevecchi WA, Barrett RT (1987) Prey selection by Gannets at breeding colonies in Norway. Ornis Scandinavica 18:319–322
Montevecchi WA, Hufthammer AK (1990) Zooarchaeological implications for prehistoric distributions of seabirds along the Norwegian coast. Arctic 43:110–114
Montevecchi WA, Myers RA (1997) Centurial and decadal oceanographic influences on changes in Northern gannet populations and diets in the north-west Atlantic: implications for climate change. ICES J Mar Sci 54:608–614. doi:10.1006/jmsc.1997.0265
Montevecchi WA, Barrett RT, Rikardsen F, Strann KB (1987) The population and reproductive status of the gannet Sula bassana in Norway in 1985. Fauna Norvegica Serie Cinclus 10:65–72
Montevecchi WA, Benvenuti S, Garthe S et al (2009) Flexible foraging tactics by a large opportunistic seabird preying on forage- and large pelagic fishes. Mar Ecol Prog Ser 385:295–306
Moss R, Wanless S, Harris MP (2002) How small Northern gannet colonies grow faster than big ones. Waterbirds Int J Waterbird Biol 25:442–448
Nelson BJ (2002) The Atlantic Gannet, 2Rev e. Fenix Books
Nussey DH, Wilson AJ, Brommer JE (2007) The evolutionary ecology of individual phenotypic plasticity in wild populations. J Evol Biol 20:831–844. doi:10.1111/j.1420-9101.2007.01300.x
Olff H, Ritchie ME, Prins HHT (2002) Global environmental controls of diversity in large herbivores. Nature 415:901–904. doi:10.1038/415901a
Pettex E, Bonadonna F, Enstipp M et al (2010) Northern gannets anticipate the spatio-temporal occurrence of their prey. J Exp Biol 213:2365–2371. doi:10.1242/jeb.042267
Piatt JF, Wetzel J, Bell K et al (2006) Predictable hotspots and foraging habitat of the endangered short-tailed albatross (Phoebastria albatrus) in the North Pacific: implications for conservation. Deep Sea Res II 53:387–398. doi:10.1016/j.dsr2.2006.01.008
Pichegru L, Grémillet D, Crawford RJM, Ryan PG (2010) Marine no-take zone rapidly benefits endangered penguin. Biol Lett 6:498–501. doi:10.1098/rsbl.2009.0913
Richner H, Heeb P (1995) Is the information center hypothesis a flop? Advances in the study of behavior. Academic Press, Massachusetts, pp 1–45
Robson BW, Goebel ME, Baker JD et al (2004) Separation of foraging habitat among breeding sites of a colonial marine predator, the Northern fur seal (Callorhinus ursinus). Can J Zool 82:20–29. doi:10.1139/z03-208
Ropert-Coudert Y, Gremillet D, Kato A et al (2004a) A fine-scale time budget of Cape gannets provides insights into the foraging strategies of coastal seabirds. Anim Behav 67:985–992
Ropert-Coudert Y, Grémillet D, Ryan P et al (2004b) Between air and water: the plunge dive of the Cape Gannet Morus capensis. Ibis 146:281–290. doi:10.1111/j.1474-919x.2003.00250.x
Røttingen I (1990) A review of variability in the distribution and abundance of Norwegian spring spawning herring and Barents Sea capelin. Polar Res 8:33–42. doi:10.1111/j.1751-8369.1990.tb00373.x
Ryan PG, Petersen SL, Peters G, Gremillet D (2004) GPS tracking a marine predator: the effects of precision, resolution and sampling rate on foraging tracks of African Penguins. Mar Biol 145:215–223
Sakshaug E, Johnsen G, Kovacs K (2009) Ecosystem Barents Sea. Tapir Academic Press, Norway, p 587
Smith JNM, Sweatman HPA (1974) Food-searching behavior of titmice in patchy environments. Ecology 55:1216–1232. doi:10.2307/1935451
Staniland IJ, Reid K, Boyd IL (2004) Comparing individual and spatial influences on foraging behaviour in Antarctic fur seals Arctocephalus gazella. Mar Ecol Prog Ser 275:263–274
Townsend CR, Begon M, Harper JL (2003) Essentials of ecology. Wiley-Blackwell, New Jersey
Townsend-Mehler J, Dyer F (2012) An integrated look at decision-making in bees as they abandon a depleted food source. Behav Ecol Sociobiol 66:275–286. doi:10.1007/s00265-011-1275-2
Votier SC, Bearhop S, Witt MJ et al (2010) Individual responses of seabirds to commercial fisheries revealed using GPS tracking, stable isotopes and vessel monitoring systems. J Appl Ecol 47:487–497. doi:10.1111/j.1365-2664.2010.01790.x
Votier SC, Grecian WJ, Patrick S, Newton J (2011) Inter-colony movements, at-sea behaviour and foraging in an immature seabird: results from GPS-PPT tracking, radio-tracking and stable isotope analysis. Mar Biol 158:355–362. doi:10.1007/s00227-010-1563-9
Wanless S, Murray S, Harris MP (2005) The status of Northern Gannet in Britain and Ireland in 2003/04. British Birds 98:280–294
Ward P, Zahavi A (1973) Importance of certain assemblages of birds as information-centers for food-finding. Ibis 115:517–534
Weimerskirch H (2007) Are seabirds foraging for unpredictable resources? Deep Sea Res Ii Top Stud Oceanogr 54:211–223
Weimerskirch H, Bertrand S, Silva J et al (2010) Use of social information in seabirds: compass rafts indicate the heading of food patches. PLoS One. doi:10.1371/journal.pone.0009928
Wilson LJ, Mc Sorley CA, Gray CM et al (2009) Radio-telemetry as a tool to define protected areas for seabirds in the marine environment. Biol Conserv 142:1808–1817
Wingfield JC (2003) Control of behavioural strategies for capricious environments. Anim Behav 66:807–816. doi:10.1006/anbe.2003.2298
Wood AG, Naef-Daenzer B, Prince PA, Croxall JP (2000) Quantifying habitat use in satellite-tracked pelagic seabirds: application of kernel estimation to albatross locations. J Avian Biol 31:278–286. doi:10.1034/j.1600-048X.2000.310302.x
Wood LJ, Fish L, Laughren J, Pauly D (2008) Assessing progress towards global marine protection targets: shortfalls in information and action. Oryx 42:340–351
Zar JH (1998) Biostatistical analysis, 4th ed. Prentice Hall, New Jersy
Zuur AF, Ieno EN, Walker N, et al (2009) Mixed effects models and extensions in ecology with R. Springer, New York
Acknowledgments
This study was funded by the SEAPOP program (financed by the Ministry of Environment, the Ministry of Petroleum and Energy and the Norwegian Oil Industry Association), the Norwegian Directorate for Nature Management, and Centre National pour la Recherche Scientifique. We warmly thank Sigbjørn Johnsen and Bjørn Dag Jenssen and his family for their logistical help in the field and Cyril Bernard at Centre d’Ecologie Fonctionnelle et Evolutive Montpellier for his precious GIS help. We acknowledge Lorien Pichegru and Jérome Fort and the two reviewers for their valuable comments and suggestions on the manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by S. Garthe.
Rights and permissions
About this article
Cite this article
Pettex, E., Lorentsen, SH., Grémillet, D. et al. Multi-scale foraging variability in Northern gannet (Morus bassanus) fuels potential foraging plasticity. Mar Biol 159, 2743–2756 (2012). https://doi.org/10.1007/s00227-012-2035-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00227-012-2035-1