
Abstract
The widespread omnivory of consumers and the trophic complexity of marine ecosystems make it difficult to infer the feeding ecology of species. The use of stable isotopic analysis plays a crucial role in elucidating trophic interactions. Here we analysed δ15N, δ13C and δ34S in chick feathers, and we used a Bayesian triple-isotope mixing model to reconstruct the diet of a generalist predator, the yellow-legged gull (Larus michahellis) that breeds in the coastal upwelling area off northwest mainland Spain. The mixing model indicated that although chicks from all colonies were fed with a high percentage of fish, there are geographical differences in their diets. While chicks from northern colonies consume higher percentages of earthworms, refuse constitutes a more important source in the diet of chicks from western colonies. The three-isotope mixing model revealed a heterogeneity in foraging habitats that would not have been apparent if only two stable isotopes had been analysed. Moreover, our work highlights the potential of adding δ34S for distinguishing not only between terrestrial and marine prey, but also between different marine species such as fish, crabs and mussels.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Ecologists have attempted to describe food web complexity and ecosystem dynamics. Substantial evidence indicates that most webs are reticulate, species are highly interconnected and many consumers have omnivorous feeding habits that span a wide trophic spectrum over their lifetimes (e.g. McCann et al. 1998; Polis and Strong 1996). This structural and dynamic complexity is of particular relevance to marine ecosystems, especially in habitats with enhanced productivity such as upwelling areas, where high species richness contributes to increased linkage density and exceptional resource abundance (Bode et al. 2003; Link 2002). Consequently, inferring consumer feeding ecology in such high-productivity ecosystems is particularly difficult, especially in the case of generalist top predators such as some seabird species, which furthermore, have the ability to exploit resources in seemingly disparate habitats (Methratta 2004; Polis and Hurd 1996). Thus, when seabirds exhibit an opportunistic foraging strategy and wide dietary plasticity, the use of complementary techniques that provide an integrative view of their feeding ecology has been helpful (Inger and Bearhop 2008; Ramos et al. 2009a).
Traditional methods, such as direct observations, analysis of regurgitated pellets, gut analysis or field experiments, can yield valuable dietary data (e.g. identification of specific prey taxa). However, the application of these techniques can be logistically difficult and in some cases may provide only limited information (Votier et al. 2003). Stable isotope signatures of consumer tissues and food resources can be used as an additional technique to infer diet (Quillfeldt et al. 2005; Ramos et al. 2009a; Sanpera et al. 2007). Stable isotope analysis (SIA) and isotopic mixing models are thus now used to study the trophic ecology (Parnell et al. 2008; Phillips and Jillian 2003; Phillips and Koch 2002). In marine ecosystems, δ13C and δ15N have commonly been used for seabird diet reconstruction (Hobson 1993; Hobson et al. 1994). Although using two isotopes may be provide useful information about simple food webs, this approach may have limited discriminatory power for generalist predators and scavengers if the food web is complex (Forero et al. 2004; Hobson and Welch 1992). In such cases, the incorporation of additional isotopic signatures has been recommended (Thompson et al. 1999; Schmutz and Hobson 1998). However, only occasionally has δ34S been used as a third dietary tracer, mainly to distinguish between marine and freshwater/terrestrial feeding (Knoff et al. 2001, 2002; Hebert et al. 2008; Hobson et al. 1997; Ramos et al. 2009a) and occasionally to differentiate between different marine resources where prey can be particularly diverse.
During the last decade, several isotopic mixing models have been developed to quantify the relative importance of different food sources from the stable isotope ratios just mentioned. The first models were algebraically constrained and restricted by the number of sources and isotopes used in the study in such a way that solutions could only be obtained when the number of sources was equal to or less than the number of isotopes plus one (Phillips 2001). Although these models proved useful for specific applications, the inherent complexity of natural systems often requires the inclusion of a large number of resources. A major advance was made when multi-source mixing models were developed (IsoSource; Phillips and Gregg 2001) which allowed an unlimited number of sources to be included independently of the number of isotopes. Recently, the development of Bayesian approaches (Inger and Bearhop 2008; Moore and Semmens 2008; Parnell et al. 2008) has allowed the incorporation not only of isotopic values, elemental concentrations and fractionation factors into mixing models, but also the uncertainties involved in all these values. This has provided results that are markedly more robust when it comes to quantifying feeding preferences.
The shelf and coastal waters off the northwest Iberian Peninsula are characterized by high levels of primary production and prey resource abundance as a result of intense upwelling. The high productivity of this marine ecosystem is of special relevance to the seabird community, especially to the yellow-legged gull (Larus michahellis), which is the most abundant breeding seabird and typically a generalist feeder (Munilla 1997a). In order to infer the feeding ecology of the different colonies of these gulls around this hotspot area, we combined the conventional pellet analysis with SIA. As few studies have explored the potential of δ34S and the application of Bayesian mixing models to infer the feeding ecology of a generalist consumer, here we (1) investigated the usefulness of adding δ34S to δ13C and δ15N to infer diet and (2) compared a Bayesian mixing model (SIAR; Parnell et al. 2008) with a multi-source model (IsoSource; Phillips and Gregg 2001) to assess the importance of incorporating uncertainty into such models.
Materials and methods
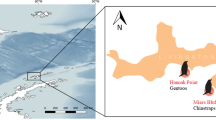
During the breeding season of 2004, yellow-legged gulls were sampled at five insular colonies in northwest Spain. Three of the colonies, Cies, Ons and Sálvora, are located off the western mainland coast and the other two colonies, Pantorgas and Ansarón, off the northern mainland coast (Fig. 1). The sampling scheme involved collecting 7–9 scapular feathers from 2- or 3-week-old chicks (n = 79) and freshly regurgitated pellets from adults (n = 219) during the chick-rearing period (from 27th May to 28th June). Chick biometric data (weight, and wing and tarsus length) were also collected. The main potential prey of yellow-legged gull chicks—inferred from fresh adult pellets and from reports in the literature (Munilla 1997a)—was also sampled in both the western and northern areas. Mussels were collected from coastal areas close to the gull colonies, earthworms from neighbouring agricultural fields and fish and crabs were obtained from local fishermen. As chicken and pork remains are obtained by yellow-legged gulls from nearby refuse dumps, samples were collected from local markets.

Geographical location of the yellow-legged gull (Larus michahellis) breeding colonies included in this study
Pellet analysis
We identified prey taxa in 219 freshly regurgitated adult pellets collected exclusively during the chick-rearing period. For each pellet, the presence or absence of prey types were recorded to describe diet using frequency and percentage of occurrence (FO or PO). In addition to refuse (represented to a large extent by chicken and pork remains), a total of 5 different prey types were identified to genus or species level: Mytilus galloprovincialis (mussels), Polybius henslowii (pelagic crabs), Micromesistius poutassou and Trachurus trachurus (fish) and Lumbricidae spp. (earthworms). We also identified other occasional prey such as Pollicipes pollicipes (cirripedes), other fish species (e.g. labrids and gadids) and small mammals. However, since they only appeared in a few pellets (1–4) in just one of the five colonies, and data collected over the same period from the same colonies during previous years (Munilla 1997a) confirmed their sporadic appearance, they were not included as potential prey in the isotopic mixing model.
Stable isotope analysis and mixing models
Feathers were cleaned in a solution of NaOH (1 M), oven dried at 60°C and kept in polyethylene bags until analysis. In the case of prey, soft tissues were extracted from Mytilus galloprovincialis, whereas the whole animal was processed for the remaining prey types (except P. henslowii, whose appendages were removed). To homogenize samples for SIA, both freeze-dried prey and feathers were ground to an extremely fine powder using an impactor mill (Freezer/mill 6750—Spex Certiprep-) operating at liquid nitrogen temperature. Once homogenized, an aliquot of prey was lipid extracted using several chloroform–methanol (2:1) rinses following Folch et al. (1957) in order to minimize the differences in δ13C caused by variable tissue lipid content (Hobson and Clark 1992).
Weighed sub-samples of the powdered feathers and prey (approximately 0.36 mg for δ13C and δ15N; 3.6 mg for δ34S) were placed into tin capsules and crimped for combustion. Isotopic analyses were carried out by elemental analysis–isotope ratio mass spectrometry (EA–IRMS) using a ThermoFinnigan Flash 1112 (for N and C)/1108 (for S) elemental analyser coupled to a Delta isotope ratio mass spectrometer via a CONFLOIII interface. Analyses were performed at the Serveis Científico-Tècnics of the University of Barcelona.
Stable isotope ratios were expressed in conventional notation as parts per thousand (‰), according to the following equation:
where X is 15N, 13C or 34S and R is the corresponding ratio 15N/14N, 13C/12C or 34S/32S.
The standards for 15N, 13C and 34S are atmospheric nitrogen (VAIR), Pee Dee Belemnite (VPDB) and Canyon Diablo Troilite (VCDT), respectively. International standards (IAEA) were inserted every 12 samples to calibrate the system and compensate for any drift over time. Precision and accuracy for δ13C measurements was ≤0.1‰, ≤0.3‰ for δ15N and ≤0.3‰ for δ34S.
In order to obtain the relative contributions of the different food sources, we used a Bayesian stable-isotope mixing model (SIAR; Parnell et al. 2008) which allows the inclusion of isotopic signatures, elemental concentrations and fractionation together with the uncertainty of these values within the model. As δ34S is less commonly used than δ13C and δ15N, we examined the results of SIAR using just δ13C and δ15N and then with the three isotopes, to find out how the inclusion of δ34S affected the estimates. In addition, we compared our results from SIAR with those obtained from the multiple-source linear mixing model proposed by Phillips and Jillian (2003) with a minor modification to incorporate the elemental concentration of each prey type (referred to hereinafter as the IsoSource model). Although the methods are intrinsically different, the IsoSource model uses the same information as SIAR except for the standard deviation, so by comparing results from the models the relevance of including this variability can be assessed. In order to use mixing models, the isotopic values for food sources must be adjusted by appropriate fractionation factors (Gannes et al. 1998). We obtained the most appropriate fractionation factor values from the literature: 5, 2.2 and 1.3‰ for δ15N, δ13C and δ34S, respectively, in the case of refuse, 4, 2.7 and 1.3‰ for earthworms (Bearhop et al. 2002; Hobson and Bairlein 2003; Peterson et al. 1985), and 3, 0.9 and 1.9‰ (Hobson and Clark 1992; Ramos et al. 2009a) for fish. Fish fractionation factors were also assumed for mussels and pelagic crabs since no experimental studies report fractionation factors of seabird diet based on marine invertebrates.
Statistical analysis
Values of stable isotope ratios were checked for normality using Q-Q plots. Descriptive statistics, estimates of the effects of interest and their 95% confidence intervals were used to show the results. Localities were compared by means of a one-way analysis of variance, and Levene’s test was used to check for homoscedasticity; Welch’s correction was applied accordingly. To test for “a posteriori” pairwise differences, we used Tukey’s procedure or the Tamhane test when variances were heterogeneous. Spearman correlations were used to examine the relationships between the three isotopes for each colony and an ANCOVA analysis to test the effect of chick size on isotopic signatures. Statistical analysis was carried out using SPSS 15.0. The Bayesian mixing model and the IsoSource model were fitted using R software (R Development Core Team 2005).
Results
Adult pellets
Polybius henslowii, a pelagic portunid crab, was the most common prey type found in adult pellets from the chick-rearing period (PO = 70%). In all colonies except Pantorgas, the majority of pellets were composed of hard parts of marine prey (mussels, pelagic crabs and fish) (Table 1). Pellets from Pantorgas indicated a predominant consumption of earthworms (PO = 62%), whereas pellets from Ons reflected a higher consumption of refuse (PO = 21%).
Prey, mixing models and chick diet
No correlation was found between chick size and isotopic signatures, so the observed differences in prey consumption between colonies are unlikely to have been caused by differences in the age of the chicks sampled. δ13C values of chick feathers showed a significant colony effect (F 4,79 = 73.19, P < 0.001) resulting in a decreasing southwest-northeast trend (Fig. 2). Cies and Ons showed the highest values of δ13C (no significant differences between them), Sálvora and Ansarón showed intermediate values and Pantorgas showed the lowest. Values of δ15N and δ34S also differed between colonies (F 4,79 = 17.06, P < 0.001; F 4,79 = 46.57, P < 0.001, respectively) but only the mean for chicks at Pantorgas was significantly different from the rest (Fig. 2). We found a significant correlation between δ15N and δ13C values in both areas (r s = 0.51, P < 0.001 and r s = 0.54, P < 0.001 for northern and western colonies, respectively); however, only in the case of the northern colonies, strong positive correlations were found between δ13C and δ34S (r s = 0.69, P < 0.001) and δ15N and δ34S (r s = 0.73, P < 0.001).

Dual stable isotope plots of nitrogen-carbon (upper panels) and sulphur-carbon (lower panels) showing isotopic signatures of potential food prey (filled symbols; mean ± SD) and chick feathers (empty symbols) sampled in the western and northern areas. Sampled size of prey and chick is given in square brackets. Lumb Lumbricidae spp.; Micr Micromesistius poutassou; Myti Mytilus galloprovincialis; Trac Trachurus trachurus; Poly Polybius henslowii; Refu Refuse
The six main prey types presented significant differences in the three isotopes, in both northern and western areas (F Welch 5,6 = 56.2, F Welch 5,16 = 173.1 for δ13C; F Welch 5,6 = 140.3, F Welch 5,17 = 76.4 for δ15N and F Welch 5,7 = 68.5, F Welch 5,17 = 571.2 for δ34S, northern and western, respectively; all P < 0.001). In both areas, post hoc Tamhane’s comparisons showed that refuse had the lowest values of δ34S, earthworms the lowest values of δ13C, pelagic crabs the highest values of δ13C and fish the highest values of δ15N (Fig. 2). With regard to the signatures of potential prey used in mixing models, aggregation of species with similar functional significance has been recommended when the isotopic signatures of prey are not significantly different (Phillips et al. 2005). However, because we are interested on the overall fish contribution to gull’s diet but also on the relative contribution of the two fish species, although their isotopic signatures are very similar in both localities, we considered them separately in the mixing model.
When we used δ13C, δ15N and δ34S, the SIAR Bayesian mixing model showed that although the consumption of fish was predominant in all the colonies (95% credibility interval: 0–44% Trachurus trachurus, 23–85% Micromesistius poutassou), a regional pattern emerged for two particular prey types. Whereas chicks from western colonies showed a higher consumption of refuse (14–21% Cies, 9–11% Ons, 10–19% Sálvora) (Fig. 3b), those from northern colonies were shown to consume more earthworms (11–22% Ansarón, 27–36% Pantorgas) (Fig. 4b). Crabs and mussels were indicated as the prey types least consumed in both western (0–23%) and northern colonies (0–12%). In contrast, when we used just δ13C and δ15N, although very similar ranges were obtained in the case of the most frequent resources for northern colonies (Fig. 4a), the model had insufficient discriminatory power in the case of western colonies (Fig. 3a). When we used two isotopes, refuse was not reflected in the diet of chicks from Cies, Ons and Sálvora, and the ranges of fish consumption varied and became wider (Fig. 3a). However, it is in the case of the three-isotope IsoSource mixing model that the ranges of fish consumed by western colonies become wider (0–87%) and the consumption percentages of minor prey types become homogenized. On the other hand, although P. henslowii presented a high variability in its δ13C signature, and more negative values have been reported (Carabel et al. 2006), we found no changes in the prey consumption ranges obtained from the mixing models when we replaced the values obtained in our analysis by more negative ones obtained from the literature.

Results of SIAR (95, 75 and 50% credibility intervals) and IsoSource mixing model (95th, 75th and 50th percentiles) showing estimated prey contributions to chick diet in each of the three western colonies. Panels in the first and second column show contributions calculated with SIAR using two -δ15N and δ13C- (a) and three -δ15N, δ13C and δ34S- (b) isotopes. Panels in the third column show IsoSource results using δ15N, δ13C and δ34S (c) with a tolerance of 1.5‰. Lumb Lumbricidae spp.; Micr Micromesistius poutassou; Myti Mytilus galloprovincialis; Trac Trachurus trachurus; Poly Polybius henslowii; Refu Refuse

Results of SIAR (95, 75 and 50% credibility intervals) and IsoSource mixing model (95th, 75th and 50th percentiles) showing estimated prey contributions to chick diet in the two northern colonies. Panels in the first and second column show contributions calculated with SIAR using two -δ15N and δ13C- (a) and three -δ15N, δ13C and δ34S- (b) isotopes. Panels in the third column show IsoSource results using δ15N, δ13C and δ34S (c) with a tolerance of 2‰. Lumb Lumbricidae spp.; Micr Micromesistius poutassou; Myti Mytilus galloprovincialis; Trac Trachurus trachurus; Poly Polybius henslowii; Refu Refuse
Discussion
Isotopic analysis of chick feathers was combined with the analysis of adult pellets to examine chick feeding ecology and its spatial variation. Although isotopic mixing models are very helpful in providing quantitative indices of food item contributions to a consumer’s diet (Caut et al. 2008; Ramos et al. 2009a), they require prior conventional diet analysis in order to correctly select potential preys. In our study, pellet data collected during the chick-rearing period combined with data from several previous years (Munilla 1997a) provided us with the necessary taxonomic resolution to select potential prey reliably. In spite of the intrinsic biases of this method (Votier et al. 2003), the description of adult pellets (Table 1) confirmed that although Pantorgas earthworms also represented an important dietary component, yellow-legged gulls from northwest Spain are, to a great extent, marine foragers (Munilla 1997a, b).
Diet selection is important in gulls for successful breeding and recruitment, and fish often constitutes a highly preferred resource (Annett and Pierotti 1999; Lewis et al. 2001; Pedrocchi et al. 1996). In contrast, marine invertebrates represent lower-quality nutrition, due to their lower calorific content and the high proportion of hard indigestible structures. However, due to their abundance and accessibility, crabs and mussels may be extensively consumed by gulls (Glutz von Blotzheim and Bauer 1982; Isenmann 1976). Although the pelagic crab P. henslowii is known to be a food resource extensively exploited by adult yellow-legged gulls during their breeding period in NW Spain (Munilla 1997b), the Bayesian three-isotope mixing model showed that fish is the main prey fed to chicks and that crabs and mussels are a supplementary resource (Figs. 3b, 4b).
The exploitation by yellow-legged gulls of food sources derived from human activities has often been described in the literature (Bosch et al. 1994; Duhem et al. 2005; Pons 1992; Ramos et al. 2009b). Refuse dumps offer yellow-legged gulls a high-energy reliable source which allows them to improve their breeding success (Pons 1992). Also, terrestrial invertebrates become an easy prey when crop fields are close to breeding colonies and can also be of considerable importance in chick diet (Ramos et al. 2009b). The mixing model not only showed a high consumption of fish, but also a clear geographical difference in the consumption of terrestrial prey between northern and western colonies. While chicks from northern colonies were fed with higher percentages of earthworms (Fig. 4b), refuse constituted a more important source in the diet of chicks from western colonies (Fig. 3b). This indicates that nearby foraging habitats such as crops and refuse dumps also provide alternative resources that this generalist and opportunistic species exploit during the chick-rearing period.
Mixing models used to quantify the relative contributions of potential prey to a consumer’s diet have undergone substantial modifications since their first application. The IsoSource mixing model derives all possible solutions that could result in a signature close enough (defined by a tolerance parameter) to the mean value observed in consumers. Because IsoSource feasible solutions can be derived independently of the number of sources and isotopes, this model has been thoroughly applied to infer the feeding ecology of invertebrates (Laurand and Riera 2006), marine mammals (Hückstädt et al. 2007), birds (Inger et al. 2006a, b; Navarro et al. 2009) and several terrestrial vertebrate species (Caut et al. 2008; Urton and Hobson 2005). Although IsoSource represented a significant advance in determining diet from stable isotope analysis, it must be parameterized using single values. Therefore, it cannot take into account consumer or resource isotopic variability, which, as reflected in our chick feathers and prey data, can be considerable. In this way, Bayesian approaches such as SIAR which incorporate uncertainty into the model have been proposed to solve this remaining problem (Parnell et al. 2008).
When we compared SIAR and IsoSource results, the inclusion of standard deviation within the model appears to have been very important in determining feasible diet proportions for yellow-legged gulls. Whereas SIAR gave relatively narrow credibility intervals, which are useful for deducing prey consumption (Figs. 3b, 4b), the empirical intervals obtained from the IsoSource model are too wide to be informative (Figs. 3c, 4c). This is probably due to the necessity to increase the tolerance parameter to 1.5‰ (western colonies) and 2‰ (northern ones) in order to obtain any solutions, when consumption ranges obtained using a small tolerance (e.g. ±0.1‰) are considered to be feasible solutions (Phillips and Jillian 2003). In the case of western colonies, even with such an increase, the range of results for the two fish species are so broad that they become uninformative about the contribution of each species to chick diet (Fig. 3c).
The inclusion of the sulphur stable isotope also seems to be a key information source to infer the diet of this generalist species. When we compare the three- and the two-isotope SIAR mixing models, the three isotopes showed higher discrimination power and contributed not only to differentiate refuse consumption, but also to detect small-range contributions of marine species that otherwise had escaped our notice. Whereas for western colonies the three-isotope mixing model reflected the heterogeneity of foraging habitats (Fig. 3b), the exclusion of δ34S from the model shifted the balance to a diet without substantial refuse contribution and to wider and less conclusive ranges for fish (Fig. 3a). Moreover, pelagic crab and mussel consumption ranges are almost undetectable when δ34S is not included in the mixing model. However, this is not the case for northern colonies. Since there was a strong correlation between the δ13C and δ34S values, and between the δ15N and δ34S values, sulphur did not contribute new information to the model, and very small changes can be observed in prey consumption ranges when δ34S is not included (Fig. 4a, b).
The use of three isotopes and the inclusion of variability within the mixing model have allowed us to distinguish a geographical pattern in diet that would otherwise have remained hidden. The simultaneous use of the three isotopes within the SIAR mixing model allowed us to differentiate not only between consumption of terrestrial and marine species, as reported previously, but also between different marine prey such as fish, crabs or mussels. Thus, our work highlights the usefulness of including sulphur for distinguishing foraging preferences and its potential to clarify trophic overlaps among and within species.
References
Annett C, Pierotti R (1999) Long-term reproductive output in western gulls: consequences of alternate tactics in diet choice. Ecology 80:288–297
Bearhop S, Waldron S, Votier SC, Furness RW (2002) Factors that influence assimilation rates and fractionation of nitrogen and carbon stable isotopes in avian blood and feathers. Physiol Biochem Zool 75:451–458
Bode A, Carrera P, Lens S (2003) The pelagic foodweb in the upwelling ecosystem of Galicia (NW Spain) during spring: natural abundance of stable carbon and nitrogen isotopes. ICES J Mar Sci 60:11–22
Bosch M, Oro D, Ruiz X (1994) Dependence of yellow-legged gulls (Larus cachinnans) on food from human activity in two western Mediterranean colonies. Avocetta 18:135–139
Carabel S, Godínez-Dominguez E, Verísimo P, Fernández L, Freire J (2006) An assessment of sample processing methods for stable isotope analyses of marine food webs. J Exp Mar Biol Ecol 336(2):254–261
Caut S, Angulo E, Courchamp F (2008) Dietary shift of an invasive predator: rats, seabirds and sea turtles. J Appl Ecol 45:428–437
Duhem C, Vidal E, Roche P, Legrand J (2005) How is the diet of yellow-legged gull chicks influenced by parents’ accessibility to landfills? Waterbirds 28(1):46–52
Folch J, Lees M, Sloane-Stanley GH (1957) A simple method for the isolation and purification of total lipids from animal tissues. J Biol Chem 226:497–509
Forero MG, Bortolotti GR, Hobson KA, Donazar JA, Bertelloti M, Blanco G (2004) High trophic overlap within the seabird community of Argentinean Patagonia: a multiscale approach. J Anim Ecol 73:789–801
Gannes LZ, del Rio CM, Koch P (1998) Natural abundance variations in stable isotopes and their potential uses in animal physiological ecology. Comp Biochem Physiol A 119:725–737
Glutz von Blotzheim UN, Bauer KM (1982) Handbuch der Vogel Mitteleuropas, Band 8/1 (Charadriiformes). Akademische Verlagsgesellschatt, Wiesbaden
Hebert CE, Bur M, Sherman D, Shutt JL (2008) Sulfur isotopes link overwinter habitat use and breeding condition in Double-crested Cormorants. Ecol Appl 18:561–567
Hobson KA (1993) Trophic relationships among high arctic seabirds: insights from tissue-dependent stable-isotope models. Mar Ecol Prog Ser 95:7–18
Hobson KA, Bairlein F (2003) Isotopic fractionation and turnover in captive Garden Warblers (Sylvia borin): implications for delineating dietary and migratory associations in wild passerines. Can J Zool 81:1630–1635
Hobson KA, Clark RG (1992) Assessing avian diets using stable isotopes II: factors influencing diet-tissue fractionation. Condor 94:189–197
Hobson KA, Welch HE (1992) Determination of trophic relationships within a high Arctic marine food web using delta-13C and delta-15 N analysis. Mar Ecol Prog Ser 84:9–18
Hobson KA, Piatt JF, Pitocchelli J (1994) Using stable isotopes to determine seabird trophic relationships. J Anim Ecol 63:786–798
Hobson KA, Hughes KD, Ewins PJ (1997) Using stable-isotope analysis to identify endogenous and exogenous sources of nutrients in eggs of migratory birds: applications to Great Lakes contaminants research. Auk 114:467–478
Hückstädt LA, Rojas CP, Antezana T (2007) Stable isotope analysis reveals pelagic foraging by the Southern sea lion in central Chile. J Exper Mar Bio Ecol 347:123–133
Inger R, Bearhop S (2008) Applications of stable isotope analyses to avian ecology. Ibis 150:447–461
Inger R, Ruxton GD, Newton J, Colhoun K, Robinson JA, Jackson AL, Bearhop S (2006a) Temporal and intrapopulation variation in prey choice of wintering geese determined by stable isotope analysis. J Anim Ecol 75:1190–1200
Inger R, Ruxton GD, Newton J, Colhoun K, Mackie K, Robinson JA, Bearhop S (2006b) Using daily ration models and stable isotope analysis to predict biomass depletion by herbivores. J Anim Ecol 43:1022–1030
Isenmann P (1976) La décharge d’ordures ménagères de Marseille comme habitat d’alimentation de la Mouette rieuse Larus ridibundus. Alauda 46(2):131–146
Knoff AJ, Macko SA, Erwin RM (2001) Diets of nesting Laughing Gulls (Larus atricilla) at the Virginia coast reserve: observations from stable isotope analysis. Isotopes Environ Health Stud 37:67–88
Knoff AJ, Macko SA, Erwin RM, Brown KM (2002) Stable isotope analysis of temporal variation in the diets of pre-fledged Laughing Gulls. Waterbirds 25:142–148
Laurand S, Riera P (2006) Trophic ecology of the supralittoral rocky shore (Roscoff, France): a dual stable isotope (δ13C, δ 15N) and experimental approach. J Sea Res 56(1):27–36
Lewis S, Wanless S, Wright PJ, Harris MP, Bull J, Elston DA (2001) Diet and breeding performance of black-legged kittiwakes Rissa tridactyla at a North Sea colony. Mar Ecol Prog Ser 221:277–284
Link J (2002) Does food web theory work for marine ecosystems? Mar Ecol Prog Ser 230:1–9
McCann K, Hastings A, Huxel GR (1998) Weak trophic interactions and the balance of nature. Nature 395:794–798
Methratta ET (2004) Top-down and bottom-up factors in tidepool communities. J Exp Mar Biol Ecol 299:77–96
Moore JW, Semmens BX (2008) Incorporating uncertainty and prior information into stable isotope mixing models. Ecol Lett 11:470–480
Munilla I (1997a) Estudio de la población y la ecología trófica de la gaviota patiamarilla (Larus cachinnans) en Galicia. Tesis Doctoral, Universidade de Santiago de Compostela. Santiago de Compostela, 328 pp
Munilla I (1997b) Henslow’s swimming crab (Polybius henslowii) as an important food for yellow-legged gulls (Larus cachinnans) in NW Spain. ICES J Mar Sci 55:631–634
Navarro J, Louzao M, Igual JM, Oro D, Delgado A, Arcos JM, Genovart M, Hobson KA, Forero MG (2009) Seasonal changes in the diet of a critically endangered seabird and the importance of trawling discards. Mar Biol 156:2571–2578
Parnell A, Inger R, Bearhop S, Jackson AL (2008) SIAR: stable isotope analysis in R. http://cran.r-project.org/web/packages/siar/index.html
Pedrocchi V, Oro D, Gonzalez-Solis J (1996) Differences between diet of adult and chick Audouin’s gulls Larus audouinii at the Chafarinas Islands, SW Mediterranean. Ornis Fenn 73:124–130
Peterson BJ, Howarth RW, Garritt RH (1985) Multiple stable isotopes used to trace the flow of organic matter in estuarine food webs. Science 227:1361–1363
Phillips DL (2001) Mixing models in analyses of diet using multiple stable isotopes: a critique. Oecologia 127:166–170
Phillips DL, Gregg JW (2001) Uncertainty in source partitioning using stable isotopes. Oecologia 127:171–179 (see also erratum, Oecologia 128:204)
Phillips DL, Jillian WG (2003) Source partitioning using stable isotopes: coping with too many sources. Oecologia 136:261–269
Phillips DL, Koch P (2002) Incorporating concentration dependence in stable isotope mixing models. Oecologia 130:114–125
Phillips DL, Newsome SD, Gregg JW (2005) Combining sources in stable isotope mixing models: alternative methods. Oecologia 144:520–524
Polis GA, Hurd SD (1996) Linking marine and terrestrial food webs: allochthonous input from the ocean supports high secondary productivity on small islands and coastal land communities. Am Nat 147:396–423
Polis GA, Strong DR (1996) Food web complexity and community dynamics. Am Nat 147:813–846
Pons JM (1992) Effects of changes in the availability of human refuse on breeding parameters in a herring gull Larus argentatus population in Brittany, France. Ardea 80:143–150
Quillfeldt P, McGill RAR, Furness RW (2005) Diet and foraging areas of Southern Ocean seabirds and their prey inferred from stable isotopes: review and case study of Wilson’s storm-petrel. Mar Ecol Prog Ser 295:295–304
R Development Core Team (2005) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. ISBN 3-900051-07-0, URL http://www.R-project.org
Ramos R, Ramírez F, Sanpera C, Jover L, Ruiz X (2009a) Feeding ecology of yellow-legged gulls (Larus michahellis) in the Western Mediterranean: a comparative assessment using conventional and isotopic methods. Mar Ecol Prog Ser 377:289–296
Ramos R, Ramírez F, Jover J, Ruiz X (2009b) Diet of yellow-legged gull (Larus michahellis) chicks along the Spanish Western Mediterranean coast: the relevance of refuse dumps. J Ornithol 150:265–272
Sanpera C, Ruiz X, Moreno R, Jover L, Waldron S (2007) Mercury and stable isotopes in feathers of Audouin’s gull as indicators of feeding habits and migratory connectivity. Condor 109:268–275
Schmutz JA, Hobson KA (1998) Geographic, temporal, and age-specific variation in diets of Glaucous Gulls in Western Alaska. Condor 100:119–130
Thompson DR, Lilliendahl K, Solmundsson J, Furness RW, Waldron S, Phillips RA (1999) Trophic relationships among six species of Icelandic seabirds as determined through stable isotope analysis. Condor 101:898–903
Urton EJM, Hobson KA (2005) Intrapopulation variation in gray wolf isotope (δ15N and δ13C) profiles: implications for the ecology of individuals. Oecologia 145(2):316–325
Votier SC, Bearhop S, MacCormick A, Ratcliffe N, Furness RW (2003) Assessing the diet of Great Skuas, Catharacta skua, using five different techniques. Polar Biol 26:20–26
Acknowledgments
Thanks are given to the Conselleria de Medio Ambiente (“Xunta de Galicia” autonomous regional government) and to the Parque Nacional de las Illas Atlánticas de Galicia, for the facilities to develop this work. We would especially like to thank C. Pérez and C. Díez (University of Vigo, Spain) for help with the collection of feathers, and M. Mulet, M. Salvande Fraga and S. Baños for help with local prey sampling. We also thank to P. Teixidor, P. Rubio, R. Roca, and E. Aracil of the Serveis Científico-Tècnics for their help in stable isotope analysis. R. Moreno was supported by an FPU grant (Ministerio de Educación y Ciencia, Spain). Funding for this work was provided by project VEM2004-08524 from the Spanish Ministerio de Educación y Ciencia and CGL2008-05448-C02-C01 and CGL2008-05448-C02-C02 from the Spanish Ministerio de Ciencia e Innovación.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by U. Sommer.
This manuscript is dedicated to Xavier Ruiz who contributed to this paper and deceased on April 27, 2008.
Rights and permissions
About this article
Cite this article
Moreno, R., Jover, L., Munilla, I. et al. A three-isotope approach to disentangling the diet of a generalist consumer: the yellow-legged gull in northwest Spain. Mar Biol 157, 545–553 (2010). https://doi.org/10.1007/s00227-009-1340-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00227-009-1340-9