
Abstract
Deep-sea hydrothermal vent ecosystems host both symbiotic and non-symbiotic invertebrates. The non-symbiotic vent fauna is generally assumed to rely on free-living chemoautotrophic bacteria as their main food source but other sources such as detritus have recently been suggested to be a part of the invertebrate diets. Little is known about how food availability influences the distribution of vent organisms on a small scale. In addition, the feeding ecology and role of small, often numerically dominant invertebrates, the meiofauna is poorly understood at vents. In this study, we used stable carbon and nitrogen isotopic analysis to investigate the role of particulate detritus in the diets of macro- and meiobenthic invertebrates within three vent assemblages at Axial Volcano, Juan de Fuca Ridge, and Northeast Pacific. Particulate organic matter of a detrital origin became more important in the diet of invertebrates in assemblages typically associated with low-hydrothermal flow intensities. Meiobenthic species occupied several different feeding guilds and trophic levels in the assemblages investigated. We conclude that small-scale spatial variability in food sources is an important feature of vent food webs and that spatial patterns observed here and elsewhere are shaped by variations in hydrothermal discharge.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Detritus at deep-sea hydrothermal vents can be highly diverse, being composed of body parts of invertebrates, faunal and microbial mucous secretions, microbial cell debris and photosynthetically derived material (Levesque and Juniper 2002; Levesque et al. 2005 this study). This compositional complexity suggests that trophic pathways other than direct grazing on free-living microbial primary producers can be of importance to the non-symbiotic hydrothermal vent fauna (Levesque et al. 2005). Further, the contribution of detritus to the diet of consumers may change as the vent sites age. Levesque and Juniper (2002) and Levesque et al. (2005) provide evidence for substantial spatial and temporal differences in the composition of particulate organic matter (POM) pools at vents on the Juan de Fuca Ridge. Their stable isotope data suggest that POM found in later successional stage vent communities and on the vent periphery tends to be more detrital in nature, in comparison with the POM from new vents and near vent openings. Also, stable isotope ratios show that the macrobenthic non-symbiotic fauna is distributed in a fragmented food web around vents, partitioning food sources (Levesque et al. 2006). At smaller spatial scales, little is known about variability in POM pools; although decimetre-scale faunal patchiness related to hydrothermal discharge is common on hydrothermal edifices of the Juan de Fuca Ridge (Sarrazin et al. 1997, 1999).
Another aspect of detrital food webs that is poorly known in the vent environment is the role of the meiofauna. Meiobenthic species from deep-sea hydrothermal vents are often endemic and their diversity is generally low compared to other marine environments (Vanreusel et al. 1997; Heptner and Ivanenko 2002). However, high-local abundances of some meiobenthic species at vents (Tsurumi et al. 2003) suggest that they could have a significant impact on the turnover of organic matter. Most discussions of the feeding ecology of meiobenthos at deep-sea hydrothermal vents have so far been speculative (Heptner and Ivanenko 2002) and their possible contribution as food sources for higher trophic levels has not been investigated. In other marine and freshwater habitats, meiofauna are known to increase food web complexity (Schmid-Araya et al. 2002) and meiobenthic invertebrates are often tightly linked to higher trophic levels such as macrofauna (Ólafsson 2003 and references therein) and fish (Smith and Coull 1987; Aarnio and Bonsdorff 1993). Meiofauna can prey on bacteria (Hicks and Coull 1983; Montagna 1984) and since chemoautotrophic bacteria and archaea form the base of food webs in hydrothermal vent ecosystems (Fisher 1990; van Dover and Fry 1994; Karl 1995; Colaço et al. 2002), the meiofauna could represent a trophic link between primary producers and macrobenthic species (Limén et al. 2007).
Stable isotope analysis of organisms and potential food sources can provide an overview of food web structure at hydrothermal vents and identify important differences between communities (Colaço et al. 2002; van Dover and Fry 1989, 1994; Fisher et al. 1994; Levesque et al. 2003). Few vent food web studies that have used stable isotope techniques have included meiobenthic species (metazoan organisms passing through a 0.5–1 mm mesh sieve and retained on a 40 μm mesh sieve) (Limén and Juniper 2006; Limén et al. 2006).
We report here on a study of three neighbouring faunal assemblages at the base of a hydrothermal sulphide edifice on Axial Volcano, on the Juan de Fuca Ridge. The primary goal of the study was to increase our understanding on the importance of detrital POM as a food source under different hydrothermal flow regimes, through stable isotope analysis of meiofaunal and macrofaunal species and POM. The contrasting nature of the habitats from where the samples were collected also permitted a limited examination of how meiofaunal composition and trophic relationships varied in relation to previously known patterns of macrofaunal distribution.
Materials and methods
Study site
Axial Volcano (46°55′N, 130°00′W) is situated in the Northeast Pacific in the central part of Juan de Fuca Ridge. An active vent field, ASHES, is located in the south west of the Axial caldera at a depth of about 1,500 m. The vent field has been active since 1986 when it was first investigated (Embley et al. 1990). The study site, “Hell”, is a sulphide chimney complex with active high-temperature venting at the summit. Venting through the porous edifice walls supplies the sides of the chimney and the immediate vicinity of the base with dilute hydrothermal fluid, and these areas of diffuse flow are colonized by vent fauna.
Sampling
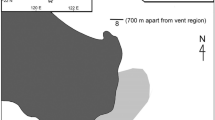
Three samples were taken from the Hell structure in July 2002 using the remotely operated vehicle ROPOS. Samples were taken with the ROPOS suction sampler at three different locations, the first sample was taken on the chimney just above a visible outflow (Fig. 1). The second sample was taken at the base of the chimney, about 30 cm away from the first sample and the third sample approximately 40 cm from the second sample, and further away from the visible source of venting. The relationship between macrofaunal species distribution and hydrothermal flow on Juan de Fuca sulphide chimneys has been described and modeled by Sarrazin et al. (1997, 1999, 2002). Based on the Sarrazin et al. model, and visual observations of hydrothermal flow and macrofaunal species composition (Fig. 1; Table 1) at each site, we concluded that vent flow intensity decreased from site 1 to site 3. Paralvinellid worms and the copepod Stygiopontius quadrispinosus were the dominant macro- and meiobenthic invertebrates at site 1. Both are common on the surfaces of sulphide chimneys subject to intense hydrothermal flow (Tunnicliffe et al. 1993; Sarrazin et al. 1997, 1999; Tsurumi et al. 2003). At site 2, the gastropod Lepetodrilus fucensis was most abundant, known to thrive in an environment of medium to low-flow intensities (Sarrazin et al. 1997, 1999). Buccinum sp. was the dominant gastropod at site 3. High abundances of Buccinum sp. have only been observed within a narrow range of weak hydrothermal conditions (Sarrazin et al. 1999).

Pork Chop chimney at the base of the Hell sulphide edifice complex, showing the three sampling sites. The distance between Sites 1 and 2 was approximately 30 cm, and between Sites 2 and 3, approximately 40 cm. Scale is 10 cm
During collection of fauna and particulate organic matter with the ROPOS suction sampler, the suction hose was positioned within 2–3 cm of the substratum surface and a low-suction pressure was applied to avoid drawing in POM from neighbouring habitats. Alternation between suction and non-suction modes allowed collected material to accumulate at the bottom of 2 L-acrylic sampling jars mounted in line with the suction hose. Two layers of 200 μm nylon mesh at the outflow of the jars retained particles inside the jars, while allowing sufficient flow to permit collection of the material. In this way, the suction sampler was used to concentrate particles in collection jars, rather than serving as an in situ filtering device. The suction hose was flushed with seawater between samples, to minimize cross-contamination. Samples remained in the suction jars at ambient bottom temperature (2°C) until brought to the surface. On board, the samples were transferred to vials and frozen at −80°C until analysis.
In the laboratory
Samples were thawed and invertebrates were identified under a stereomicroscope. Depending on the size of the animal, specimens were prepared for stable isotope analysis either individually or pooled into a mass sufficient for the analysis (Table 1). Polychaetes were first dissected in order to remove gut contents and then acidified with 0.1 N HCl in glass vials, rinsed once with Milli-Q water and dried in an oven at 55°C for 24 h. The dried animals were ground in the vials with a glass rod and a calculated amount was transferred to a tin capsule (8 × 5 mm D1008-Elemental Microanalysis Limited). Small invertebrates, juvenile macrofauna and meiofauna were acidified in the tin capsules. A thick tin capsule (“smooth wall tin capsule” D4057, Elemental Microanalysis Limited) was used to prevent capsule damage by HCl. Stable carbon and nitrogen isotopic composition were measured using a Micromass Isoprime isotope ratio mass spectrometer, in line with a Carlo Erba C/N element analyzer. Meiofauna and juvenile macrofauna were analyzed with a trap current of 600 μA and macrofauna with a trap current of 200 μA. Stable isotopic compositions are reported relative to Vienna Pee-Dee Belemnite and atmospheric nitrogen as:
where X is 13C or 15N, and R is 13C/12C or 15N/14N.
The particulate matter was fractionated into four size categories in order to investigate the variation of the δ13C of different fractions. The wet sediment was passed through sieves to obtain the four size classes; >500, 250–500, 125–250 and <125 μm. Each fraction was dried, ground and prepared for stable isotope analysis similar to invertebrates.
Data treatment: baseline correction
Vent fluid dissolved inorganic carbon (DIC) at Axial Volcano is enriched in 13C compared to seawater DIC (Levesque et al. 2005). With increasing distance from the source of venting, increasing dilution of vent fluid with seawater decreases the average δ13C of the DIC pool, which will be reflected in primary producers as they fix CO2. This, in turn, will be reflected in the consumer species, and so requires a correction for variation at the base of the food web (baseline correction) in order to make data comparable between different locations (Vander Zanden and Rasmussen 1999; Post 2002; Levesque et al. 2006). At hydrothermal vents, invertebrates depending exclusively on chemoautotrophic bacterial endosymbionts, such as the tubeworm Ridgeia piscesae, reflect the isotopic composition of primary producers, which in turn will vary with the isotopic composition of the source DIC. In order to correct for decreasing 13C enrichment in DIC from the most intensive hydrothermal flow at site 1 to the low-flow regime at site 3, we used the isotopic signature of the tubeworm Ridgeia piscesae for baseline corrections at our three sites (Levesque et al. 2006). The correction factor at site 2 was calculated as the difference between the isotopic signature of Ridgeia at site 1 with Ridgeia at site 2, and similarly at site 3 invertebrates were corrected for the differences between Ridgeia at this site and site 1. The maximum correction was less than 1.5‰.
Data treatment: mixing model
We used a two-source mixing model (Phillips and Gregg 2001) in order to calculate the relative contributions of chemoautotrophic bacteria (δ13C = −11‰) and particulate organic matter, (POM; δ13C = −17, −20, and −26‰ at site 1, 2 and 3, respectively) to the diet of organisms.
We were unable to separate chemoautotrophic bacteria from POM and other debris in order to measure their isotopic signature directly. Instead, we based our chemosynthetic end member on the stable carbon isotope signature of the sulphide worm Paralvinella sulphincola. This polychaete has been shown to feed directly, and likely exclusively, on microbial biofilms on the substratum around its tube opening (Morineaux et al. 2002; Grelon et al. 2006). Trophic-level corrected stable carbon isotope signatures for P. sulfincola from site 1 (−10.0‰) were close to the circa −11‰ values typically attributed to carbon fixed by chemolithoautotrophs using form II Ribulose biphosphate carboxylase (Robinson and Cavanaugh 1995).
The second endmember in our mixing model, POM, is composed of debris including decaying body parts, mucus and small fractions of photosynthetically derived material and living bacteria (Levesque et al. 2005). The δ13C of POM in the calculations comprised all size fractions in their original proportions, from each site.
The following two equations were used to calculate the relative contributions of the two food sources to the diet of the consumer organisms at each site:
POMtot, the total POM fraction (POM + adhering chemoautotrophic bacteria); POMfrac, the POM fraction without chemoautotrophic bacteria; bacta, chemoautotrophic bacteria (adhering to POM); m = 0.005, representing the relative contribution of bacterial carbon to the particulate matter (POMtot) carbon pool as determined in similar environments at Axial Volcano vents (Levesque et al. 2005)
Org, organism (0.4 was used to correct for the carbon isotopic fractionation in the organisms (Post 2002)); x, the proportion of bacteria in the diet of the organism, bactf, bacterial mats and biofilms not associated with POM. This bacterial source would be available for direct grazing by the fauna. See “Discussion” for further explanation.
Data treatment: statistics
Since only single samples were taken from each vent assemblage, no parametric test could be performed to compare community composition and stable isotopic signatures of invertebrates between sites. Replicate measurements of stable isotopic composition of separate specimens or specimen pools within each sample are presented to illustrate the variation among individuals from each site.
Spearman rank correlation was used to examine the relationship of invertebrate carbon isotopic compositions between the three vent assemblages, and the relationship of δ13C and C/N ratios of the particulate matter.
Results
Mixing model
Carbon mixing model calculations with chemoautotropic bacteria and POMfrac as end members, showed that the relative contribution of chemoautotrophic bacteria to the diet of organisms was highest at site 1 and that POM became more important in the diet of organisms at sites 2 and 3 (Fig. 2). Chemoautotrophic bacteria formed, on average, 70% of the diet of the organisms at site 1 and 25% at the other two sites. At site 1, the two Paralvinellid worms derived their food entirely from bacteria, while the remaining four species depended on a mixture of the two sources where bacteria contributed 40–90% of the diet. No exclusive bacterial feeders were found at sites 2 and 3, whereas POM formed the main diet of three species at site 2 and two species at site 3 (Fig. 2). Also, at least 70% of the combined faunal diet (combined histograms in Fig. 2) at sites 2 and 3 was derived from POM (Fig. 2).

The proportion of chemoautotrophic bacteria and POM in the diet of macro- and meiobenthic organisms from the three sites, based on a two-source mixing model. Abbreviations stand for: Paralvinella sulfincola (P. s) Paralvinella palmiformis (P. p), Lepidonotopodium sp. (L.), Stygiopontius quadrispinosus (S. q), Lepetodrilus fucensis (L. f), Depressigyra globulus (D. g), Scale worm (Sc. worm), Amphysamytha galapagensis (A. g), Euphilomedes climax (E. c), Buccinum sp. (B.), Aphotopontius forcipatus (A .f), Nereis sp. (N.), Ammothea verenae (A. v), Paracanthonchus sp. (P.), Copidognathus papillatus (C. p), juvenile crab (juv crab)
The carbon isotopic signature of particulate matter (comprising all size fractions) became more depleted and the C/N ratio increased from site 1 to 3 (P < 0.001, R = −0.82, Table 2; Fig. 3). The range of carbon isotopic composition among the size fractions of particulate matter also increased with distance from the vent fluid source (Fig. 4; Table 2). The range of δ13C of the different size-fractionated classes of particulate matter was 0.6, 1.1 and 4.7‰ at sites 1–3, respectively, (Fig. 4, Table 2). There was no similar trend for within-site variation in nitrogen isotopic (Fig. 4, Table 2).

The relation between δ13C and C/N of the size fractionated particulate matter from the three sites along the transect, P < 0.001, Spearman R = −0.82

Baseline corrected stable carbon (δ13C) and nitrogen (δ15N) isotopic composition of macro- and meiobenthic invertebrates at sites 1, 2 and 3. Error bars represent standard error (SE). The box in each graph represent the isotopic range of the size fractionated particulate matter
Invertebrates
The δ13C baseline corrections for the invertebrates at sites 2 and 3, respectively, 0.8 and 1.5‰ are incorporated in the following results. All non-symbiotic species became more 13C-depleted from site 1 to 3 (R = −0.69, P < 0.001, Fig. 4). The range of carbon isotopic signatures of invertebrates was broader at sites 2 and 3 (9‰) compared to site 1 (6‰) (Fig. 4). At site 1, Paralvinella sulfincola (δ13C = −10.0‰) was the most 13C-enriched species and the juvenile gastropods, Buccinum sp. (δ13C = −24.4‰) and Depressigyra globulus (δ13C = −24.4‰) at site 3 were the most 13C-depleted invertebrates (Fig. 4). The range of stable nitrogen isotopic composition of invertebrates (excluding the vestimentiferan Ridgeia piscesae) was higher at site 3 being on average 6‰ compared to 4.0‰ at sites 1, 2 (Fig. 4). The nitrogen isotopic ratio was lowest in the copepod Stygiopontius quadrispinosus at site 1 (δ15N = 4.7‰) and highest in the juvenile crab at site 3 (δ15N = 10.4‰; Fig. 4).
The copepod species, Aphotopontius forcipatus and Stygiopontius quadrispinosus, co-occurred at site 2. The stable carbon isotopic composition of A. forcipatus was −22.2‰, while that of S. quadrispinosus was −13.5‰. Both species were found in relatively low abundances in the sample from this site, 18 and 61 specimens, respectively. The total number of specimens for each species was too low to obtain results for stable nitrogen isotopes.
Among the meiobenthic species within the three assemblages, ostracods, Euphilomedes climax at site 2 and the nematode species Paracanthonchus sp. at site 3 were most enriched in δ15N, measuring 7.8 and 7.6‰, respectively.
Small sized Buccinum sp. were more 13C-depleted than adults at sites 2 and 3 (Fig. 4). At site 3, three distinct size classes of Buccinum sp. were found and the intermediate size class of Buccinum sp. was more depleted in δ15N than smaller and larger specimens of the same species. Small-sized Depressigyra globulus at site 3 were relatively depleted in 13C compared to larger D. globulus at site 2 (Fig. 4).
Meiofauna and macrofauna distribution
A total of 13 meiobenthic species and groups were found in the three vent assemblages. While only one meiobenthic species was found at site 1, six species were found at site 2 and ten at site 3. The numerically most important meiobenthic groups were nematodes and copepods. The copepod Stygiopontius quadrispinosus was the only meiobenthic species at site 1, close to the chimney, where it was the most abundant invertebrate. The male and female ratio was highly skewed in favor of females (Table 1). Nematodes, composed of five species, were numerically dominant at site 3. Two species, Paracanthonchus sp. and Leptolaimus sp., comprised 94% of the total number of nematodes or 64 and 30%, respectively, (Table 1). A stable isotopic signature was obtained for one nematode species, Paracanthonchus sp., which was the only species found in sufficient numbers for the analysis. The ostracod species Euphilomedes climax was the second most abundant meiobenthic organism at site 2 and the third most abundant at site 3 (Table 1).
Altogether, 12 macrobenthic species and groups were found in the three assemblages. The most abundant species was the gastropod Lepetodrilus fucensis, which were present at all three sites. Close to the chimney, Paralvinella sp. dominated in abundance. At the last site, furthest away from the chimney base, Buccinum sp. was most frequent. Similar to meiofauna, the highest number of species was found at the site furthest away from the chimney (Table 1).
Discussion
The mixing model
The mixing model based on chemoautotrophic bacteria and POMfrac as endmembers indicates that most species have a mixed diet at the three sites. There were, however, notable exceptions to this trend, with species entirely relying on bacteria occurring at site 1 and species specializing in POM at sites 2 and 3. The Paralvinellid worms, dominant at site 1, appeared as exclusive feeders on free-living chemolithoautotrophic bacteria/Archaea. In situ feeding studies have shown that the polychaete Paralvinella sulfincola constantly probes and grazes most of the chimney surface within its habitat (Grelon et al. 2006; Morineaux et al. 2002). Filamentous bacteria growing on the tubes of the worms may also serve as a food source (Morineaux et al. 2002).
According to the mixing model, juvenile Depressigyra globulus and Buccinum sp. (Gastropoda) depend on detritus at sites 2 and 3. At site 2, the carbon isotopic signature 2–5 mm of Buccinum sp. was more 13C-depleted than the POM fraction and therefore outside the range of our presumed end members. One explanation may be that these smaller snails may be selectively feeding on parts of the detritus depleted in carbon. The detrital pool found at vents has been found to be quite heterogeneous (Levesque et al. 2005). At each of our of sites, some POM size fractions had notably different isotopic signatures, suggesting separate origins and a mixed composition. Alternatively, the δ13C signature of small snails may reflect feeding in a different location prior to recruiting to the sampling site.
In the mixing model, we presumed that the bacterial fraction was 0.5% of the total particulate organic carbon as calculated from similar environments in the North East Pacific (Levesque et al. 2005). This may be an underestimation of the actual bacterial fraction available to consumers. If the grazing pressure is high, the bacterial turnover rate may, in turn, be high, so that the bacterial fraction present on the POM may not mirror the bacterial food source available to consumers who can selectively graze or digest bacteria attached to particles. However, the results from the mixing model did not change significantly even when we simulated a situation where the bacterial fraction would be 10 times higher or 5%. Food sources are often patchy in the environment and biofilms of bacteria on hard sulphide substrates and on tubes of Paralvinellid worms are frequent. Although neither microbial biofilms nor POM were quantitatively sampled in this study, the data do suggest that microbial carbon was most available to consumers at site 1. A study along a thermal gradient of a shallow-water hydrothermal vents showed that the abundance of bacteria and archaea was highest, close to the vent indicating that the microbial activity is stimulated by the high temperature and vent fluid flow (Sievert et al. 2000). Similarly, a colonization experiment in the vicinity of deep-sea hydrothermal vents showed that thick biofilms developed within days at sites under the influence of vent fluid flow but a sparser biofilm formed at a low temperature site with little vent influence (Guezennec et al. 1998).
The wide range of POM δ13C at site 3 indicates that the detrital pool was more heterogeneous than at the other two sites. This was reflected in the wide range of carbon isotopic signatures in the consumer species. Food resource partitioning seems to be more important at site 3 compared to the two other sites. Trophic complexity was also greatest at site 3, where a predator trophic level with an enriched 15N signal was apparent, represented by juvenile crabs and nereid polychaetes. It thus appears that spatial differences in vent flow intensity, in addition to already demonstrated effects on sulphide edifice community composition (Sarrazin et al. 1997, 1999), can determine the importance of detritus in the diet of vent organisms, and trophic complexity.
POM stable isotope characteristics
In the low-flow habitat (site 3) at the periphery of the venting chimney, photosynthetically derived detrital material may make a measurable contribution to the POM pool. Organic matter in pelagic sediment from this area has δ13C values around −22 to −26‰ (Levesque et al. 2005), therefore a higher proportion of this allochtonous material is related to a lower δ13C of particulate matter. A greater relative contribution of photosynthetically derived detritus to POM at the periphery is also supported by the higher C/N ratio with increasing distance from the chimney. Sediment from the same area has an average C/N ratio of 17.4 (Levesque et al. 2005). The negative relationship between C/N and δ13C (Fig. 3) supports the hypothesis that increasing depletion in 13C of POM with increasing distance from the vent is at least in part due to an increasing fraction of nitrogen-poor pelagic sediment in the POM pool. The trend in δ13C versus C/N from site 1 to site 3 also argues against an alternative explanation for 13C depletion in POM and faunal tissues away from the intense discharge at site 1. This alternative explanation would require increasing (from site 1 to 3) input of fresh organic material from another chemosynthetic source, one with a more negative δ13C signature, the so-called −30‰ or from I Rubisco end member proposed for some vent food webs (Van Dover and Fry 1994; Robinson and Cavanaugh 1995; Limén and Juniper 2006). Similar patterns in isotopic signatures and C/N ratio of particulate matter were observed in a 2-year study of another location on Axial Volcano, along a gradient away from a point source of venting (C. Levesque, unpublished data). Nevertheless, the large range of carbon isotopic composition of POM size fractions at site 3 is indicative of organic carbon originating from several sources.
Meiofauna
Stable carbon and nitrogen isotopic signatures of meiobenthic species in the three assemblages indicate that they belong to various feeding groups. The copepod Stygiopontius quadrispinosus is often found in high abundance on sulphide chimney surfaces exposed to hot vent fluids (Tsurumi et al. 2003, this study). Its relatively depleted δ15N signature in combination with its spatial distribution indicates that the species is a first order consumer. Results from the mixing model indicated that it grazes on a mixture of chemolitoautotrophic bacteria and POM close to the vent, with bacteria constituting at least 60% of its food source.
Evidence for food partitioning between the co-habiting copepods Stygiopontius quadrispinosus and Aphotopontius forcipatus was found at site 2. Their stable carbon signatures were −14.3 and −23.0‰, respectively. The mixing model indicated that, again, 60% of the diet of S. quadrispinosus comes from bacteria, while A. forcipatus depends entirely on POM. Similarly, the stable carbon isotopic composition of these two species co-occurring at another site from the ASHES vent field in 2001 showed a similar pattern; −15.5‰ for S. quadrispinosus and −20.4‰ for A. forcipatus (Limén, unpublished data). The δ15N for A. forcipatus, measured in 2001, was 6.5‰ but no reliable δ15N data for S. quadrispinosus could be obtained from this site since we had only 29 specimens and a minimum of 100 is needed for reliable δ15N analysis (Limén and Marty 2004). Similarly, in this study, only at site 1 did we have enough individuals of S. quadrispinosus (δ15N = 4.7‰) for nitrogen isotope analysis. Tsurumi et al. (2003), in a study of the spatial distribution of these two copepod species, also concluded that they exploited different food sources. They found that S. quadrispinosus dominated at high-flow sites near chimneys while A. forcipatus was more common at low-flow sites.
Nematodes, the most abundant meiobenthic taxon at site 3, were dominated by two species: Paracanthonchus sp. and Leptolaimus sp. The two genera have been previously recorded from deep-sea sediments (Jensen 1991; Vanaverbeke et al. 1997; Vanhove et al. 1999, 2004; Steyaert et al. 1999; Gambi et al. 2003) and found to be frequent in hydrothermal sediments of the North Fiji Basin (Vanreusel et al. 1997). Experimental data (Modig and Ólafsson 1998) showed that two Leptolaimus species, frequent in the Baltic Sea, are adapted to survive severe environmental conditions such as long periods of low-oxygen conditions. Leptolaimus sp. has a minute mouth opening and is considered to be a “microvore” (Moens and Vincx 1997), feeding on small food items such as bacteria. Chemoautotrophic bacteria and/or heterotrophic bacteria are most likely the main food source for Leptolaimus sp. at site 3. Our sample did not contain sufficient numbers of Leptolaimus sp. to be able to confirm this with stable isotopic data. Paracanthonchus sp., found at site 3, has a wide mouth opening armed with a large tooth. Species within this genus is usually considered to be epigrowth feeders relying on diatoms in shallower marine habitats (Ólafsson et al. 1999). Here, the nitrogen isotopic composition of Paracanthonchus sp. (+8.74), indicates that it has a predatory/scavenging feeding habit as it was relatively enriched in 15N. Several nematodes show opportunistic feeding and can change feeding behavior depending on the availability of food sources (Moens and Vincx 1997). A large part of the particulate matter at hydrothermal vents consists of decaying body parts of macrobenthic animals, which may serve as food for other organisms such as Paracanthonchus sp. The ostracod Euphilomedes climax (Kornicker 1991) was relatively enriched in 15N at site 2. Most members of its family (Philomedidae) are detritus feeders but examination of E. climax gut contents in other habitats has shown that it may also ingest small invertebrates such as copepods (Kornicker 1991). The stable nitrogen isotopic composition of the ostracod observed here supports the hypothesis that it can exploit food sources other than bacteria and detritus, i.e. possibly small invertebrates and/or decaying animals.
Conclusion
Spatial variation in POM utilization has previously been observed around basalt-hosted diffuse flow vents where the contribution of POM to faunal diets increased along habitat gradients away from the primary source of venting (Levesque et al. 2006). This study provides first evidence for an analogous situation on sulphide edifices, where flow variations create habitat patchiness rather than horizontal gradients (Sarrazin et al. 1997, 1999), and so influence POM composition and utilization. Our stable isotope data indicate that bacterivory is more frequent where hydrothermal flow is more intense, whereas the large and heterogeneous POM pool can significantly contribute to the diet of organisms in areas of lesser fluid discharge, in this case at the base of an active chimney.
This study has also provided first evidence for spatial variation in meiofaunal assemblages on sulphide edifices, where we found that meiofauna act as bacterial grazers, detritus feeders, predators and scavengers, and occupy several different feeding guilds and trophic levels. Evidence for food partitioning between copepods was also found.
In summary, our results together with those of several recent studies (Colaçao et al. 2002; Limén and Juniper 2006; Levesque et al. 2006; Bergquist et al. 2007) support an emerging view of hydrothermal vent food webs wherein non-symbiotic faunal species partition multiple food sources whose relative abundance and utilization vary at decimetre scales in relation to hydrothermal discharge.
References
Aarnio K, Bonsdorff E (1993) Seasonal variation in abundance and diet of the sand goby, Pomatoschistus minutus (Pallas), in a northern Baltic archipelago. Ophelia 37:19–30
Bergquist DC, Eckner JT, Urcuyo IA, Cordes EE, Hourdez S, Macko SA, Fisher CR (2007) Using stable isotopes and quantitative community characteristics to determine a local hydrothermal vent food web. Mar Ecol Prog Ser 330:49–65
Colaço A, Dehairs F, Desbruyères D (2002) Nutritional relations of deep-sea hydrothermal fields at the Mid-Atlantic Ridge: a stable isotope approach. Deep Sea Res I 49:395–412
Embley RW, Murphy KM, Fox CG (1990) High-resolution studies of the summit of Axial Volcano. J Geophys Res 95:12785–12812
Fisher CR (1990) Chemoautotrophic and methanotrophic symbiosis in marine invertebrates. Rev Aquat Sci 2:399–436
Fisher CR, Childress JJ, Macko SA, Brooks JM (1994) Nutritional interactions in Galapagos Rift hydrothermal vent communities: inferences from stable carbon and nitrogen isotope analyses. Mar Ecol Prog Ser 103:45–55
Gambi C, Vanreusel A, Danovaro R (2003) Biodiversity of nematode assemblages from deep-sea sediments of the Atacama Slope and Trench (South Pacific Ocean). Deep Sea Res I 50:103–117
Grelon D, Morineaux M, Desrosiers G, Juniper SK (2006) Feeding and territorial behavior of Paralvinella sulfincola a polychaete worm at deep-sea hydrothermal vents of the Northeast Pacific Ocean. J Exp Mar Biol Ecol 329:174–186
Guezennec J, Ortega-Morales O, Raguenes G, Geesey G (1998) Bacterial colonization of artificial substrate in the vicinity of deep-sea hydrothermal vents. FEMS Microbiol Ecol 26:89–99
Heptner MV, Ivanenko VN (2002) Copepoda (Crustacea) of hydrothermal ecosystems of the World Ocean. Arthropoda Selecta 11:117–134
Hicks GR, Coull BC (1983) The ecology of marine meiobenthic harpacticoid copepods. Oceanogr Mar Biol Rev 21:67–175
Jensen P (1991) 9 new and less known nematode species from the deep-sea benthos of the Norwegian Sea. Hydrobiologia 222:57–76
Karl DM (1995) Ecology of free-living, hydrothermal vent microbial communities. In: Karl DM (ed) Microbiology of extreme and unusual environments, vol 2. Deep-sea hydrothermal vent habitats. CRC, New York, pp 35–124
Kornicker L (1991) Myodocopid Ostracoda of hydrothermal vents in the eastern Pacific Ocean. Smithson Contrib Zool 516:1–46
Levesque C, Juniper SK (2002) Particulate matter as a food source at a nascent hydrothermal vent on the Juan de Fuca Ridge. Cah Biol Mar 43:289–292
Levesque C, Juniper SK, Marcus J (2003) Food resource partitioning and competition among alvinellid polychaetes of Juan de Fuca Ridge hydrothermal vents. Mar Ecol Prog Ser 246:173–182
Levesque C, Limén H, Juniper SK (2005) Origin, composition and nutritional quality of particulate matter at deep-sea hydrothermal vents on Axial Volcano, northeast Pacific. Mar Ecol Prog Ser 289:43–52
Levesque C, Juniper SK, Limén H (2006) Spatial organization of food webs along habitat gradients at deep-sea hydrothermal vents on Axial Volcano, Northeast Pacific. Deep Sea Res I 53:726–739
Limén H, Juniper SK (2006) Habitat controls on vent food webs at Eifuku Volcano, Mariana Arc. Cah Biol Mar 47:449–455
Limén H, Marty J (2004) Stable carbon and nitrogen isotope analysis on small-sized samples: a protocol for preparation and analysing microscopic organisms. Application Note, GV Instruments AN13
Limén H, Juniper SK, Tunnicliffe V, Clément M (2006) Benthic community structure on two peaks of an eruptive seamount: Northwest Rota−1 Volcano, Mariana Arc, western Pacific. Cah Biol Mar 47:457–463
Limén H, Stevens C J, Bourass Z, Juniper SK (2007) Copepods—a potential trophic link between hydrothermal vent bacteria and Paralvinellid polychaetes: lipid and stable isotope evidence. Mar Ecol Prog Ser (submitted)
Modig H, Ólafsson E (1998) Responses of Baltic benthic invertebrates to hypoxic events. J Exp Mar Biol Ecol 229:133–148
Moens T, Vincx M (1997) Observations on the feeding ecology of estuarine nematodes. J Mar Biol Assoc UK 77:211–227
Montagna PA (1984) In situ measurement of meiobenthic grazing rates on sediment bacteria and edaphic diatoms. Mar Ecol Prog Ser 18:119–130
Morineaux M, Grelon D, Juniper SK (2002) Utilization of nutritional resources and energy budgets for populations of the hydrothermal vent polychaete Paralvinella sulfincola on Axial Volcano, Juan de Fuca Ridge (Northeast Pacific). Cah Biol Mar 43:241–244
Ólafsson E (2003) Do macrofauna structure meiofauna assemblages in marine soft-bottoms? A review of experimental studies. Vie et Milieu 53:249–265
Ólafsson E, Modig H, van de Bund WJ (1999) Species specific uptake of radio-labelled phytodetritus by benthic meiofauna from the Baltic Sea. Mar Ecol Prog Ser 177:63–72
Phillips DL, Gregg JW (2001) Uncertainty in source partitioning using stable isotopes. Oecologia 127:171–179
Post DM (2002) Using stable isotopes to estimate trophic position: models, methods, and assumptions. Ecology 83:703–718
Robinson JJ, Cavanaugh CM (1995) Expression of form I and form II Ribulose-1,5-biphosphate carboxylase/oxygenase (Rubisco) in chemoautotrophic symbiosis: implications for the interpretation of stable isotope ratios. Limnol. Oceanogr. 40:1496–1502
Sarrazin J, Robigou V, Juniper SK, Delaney JR (1997) Biological and geological dynamics over four years on a high-temperature sulfide structure at the Juan de Fuca Ridge hydrothermal observatory. Mar Ecol Prog Ser 153:5–24
Sarrazin J, Juniper SK, Massoth G, Legendre P (1999) Physical and chemical factors influencing species distributions on hydrothermal sulfide edifices of the Juan de Fuca Ridge, northeast Pacific. Mar Ecol Prog Ser 190:89–112
Sarrazin J, Levesque C, Juniper SK, Tivey MK (2002) Mosaic community dynamics of Juan de Fuca Ridge sulfide edifices: Substratum, temperature and implications for trophic structure. Cah Biol Mar 43:275–279
Schmid-Araya JM, Hildrew AG, Robertson A, Schmid PE, Winterbottom J (2002) The importance of meiofauna in food webs: evidence from an acid stream. Ecology 83:1271–1285
Sievert SM, Ziebis W, Kuever J, Sahm K (2000) Relative abundance of Archaea and Bacteria along a thermal gradient of a shallow-water hydrothermal vent quantified by rRNA slot-blot hybridization. Microbiology 146:1287–1293
Smith LD, Coull BC (1987) Juvenile spot (Pisces) and grass shrimp predation on meiobenthos in muddy and sandy substrata. J Exp Mar Biol Ecol 105:123–136
Steyaert M, Garner N, van Gansbeke D, Vincx M (1999) Nematode communities from the North Sea: environmental controls on species diversity and vertical distribution within the sediment. J Mar Biol Assoc UK 79:253–264
Tsurumi M, de Graaf RC, Tunnicliffe V (2003) Distributional and biological aspects of copepods at hydrothermal vents on the Juan de Fuca Ridge, north-east Pacific ocean. J Mar Biol Assoc 83:469–477
Tunnicliffe V, Desbruyères D, Jollivet D, Laubier L (1993) Systematic and ecological characteristics of Paralvinella sulfincola Desbruyères and Laubier, a new polychaete (family Alvinellidae) from northeast Pacific Hydrothermal vents. Can J Zool 71:286–297
Van Dover CL, Fry B (1989) Stable isotopic compositions of hydrothermal vent organisms. Mar Biol 102:257–263
Van Dover CL, Fry B (1994) Microorganisms as food resources at deep-sea hydrothermal vents. Limnol Oceanogr 39:51–57
Vanaverbeke J, Arbizu PM, Dahms HU, Schminke HK (1997) The metazoan meiobenthos along a depth gradient in the Arctic Laptev Sea with special attention to nematode communities. Polar Biol 18:391–401
Vander Zanden MJ, Rasmussen JB (1999) Primary consumer δ13C and δ15N and the trophic position of aquatic consumers. Ecology 80:1395–1404
Vanhove S, Arntz W, Vincx M (1999) Comparative study of the nematode communities on the southeastern Weddell Sea shelf and slope (Antarctica). Mar Ecol Prog Ser 181:237–256
Vanhove S, Vermeeren H, Vanreusel A (2004) Meiofauna towards the South Sandwich Trench (750–6300 m), focus on nematodes. Deep Sea Res II 51:1665–1687
Vanreusel A, Van den Bossche I, Thiermann F (1997) Free-living marine nematodes from hydrothermal sediments: similarities with communities from diverse reduced habitats. Mar Ecol Prog Ser 157:207–218
Acknowledgments
We would like to thank the ROPOS crew and the Captain and crew of the R/V Thomas G. Thompson for assistance during sampling and C. Lalande and R. Léveillé for collecting samples. Thanks to M. Morineaux for help with sorting and preparing samples for stable isotope analysis and to A. Adamowicz, L. Cournoyer and J. McKay for performing the stable isotope analysis. Also, many thanks to J. Marty for valuable comments on the manuscript. This study was funded by an NSERC Canada Collaborative Research Opportunities grant to the CanRidge project and an NSERC Discovery grant to SKJ. This study was performed in Canada and is in compliance with current laws.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by R.J. Thompson.
Rights and permissions
About this article
Cite this article
Limén, H., Levesque, C. & Kim Juniper, S. POM in macro-/meiofaunal food webs associated with three flow regimes at deep-sea hydrothermal vents on Axial Volcano, Juan de Fuca Ridge. Mar Biol 153, 129–139 (2007). https://doi.org/10.1007/s00227-007-0790-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00227-007-0790-1