
Abstract
Intertidal endobenthic bivalves are often dislodged from sediments by hydrodynamic forces. As a result, they encounter the dangers of predation and desiccation, which are generally harsh near the sediment surface. To cope with such dangers, the bivalves possibly possess: (1) a strong body to endure predation and desiccation stress, (2) quick mobility to avoid the stresses, or (3) a high growth rate for attaining a size refuge. The present study examined which of these modes are adopted by the subtropical cobbled-shore Venus clams Gafrarium tumidum (Röding, 1798) and Ruditapes variegatus (Sowerby, 1852), revealing the following interspecific differences. (1) G. tumidum survived better than R. variegatus did in harsh experimental conditions, namely: the experimental cages exposed to predation and desiccation on a cobbled shore; a laboratory aquarium with a predatory crab Scylla serrata; and ovens with high temperatures (27°C and 34°C). (2) R. variegatus was more mobile than G. tumidum was, digging into the sediment on a cobbled shore more rapidly at both high and low tides. (3) The two species with shell lengths 20–30 mm showed similar growth rates (median: −0.2 to 44.5 μm day−1) in seasonal mark–recapture surveys over 2 years. Overall, to cope with the dangers of predation and desiccation G. tumidum appears to have a strong body, while R. variegatus displays rapid mobility, and neither species seems to attain a size refuge through rapid growth. Such species-specific modes are discussed in relation to the interspecific differences found in shell morphology.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
In the intertidal zone, endobenthic bivalves are relatively well protected from the stress of predation and desiccation compared to epibenthic bivalves. Predators are generally rarer under the sediment surface (e.g. polychaete: Kurihara et al. 2000; Seitz et al. 2001) than above the surface (gastropod, octopus, crab, starfish, fish, bird: Jangoux 1982; Rodrigues 1986; Eggleston et al. 1992; Hilgerloh 1997; Hines et al. 1997; Leonard et al. 1999; Seitz et al. 2001; for general information see Vermeij 1978; Commito and Ambrose 1985). Epibenthic predators often fail to attack prey hidden in the sediment (Blundon and Kennedy 1982b; Zwarts and Wanink 1989; Zaklan and Ydenberg 1997; Seitz et al. 2001). Desiccation is weaker in the sediment than on the sediment surface due to moisture and shading.
Even endobenthic bivalves, however, encounter intense predation and desiccation in the intertidal zone, when dislodged from the sediment by hydrodynamic forces. Such dislodgment has been reported from soft-bottom shores for the bivalves Mya arenaria (Matthiessen 1960; Emerson and Grant 1991), Gemma gemma (Commito et al. 1995) and Macomona liliana (Turner et al. 1997). Dislodgment presumably also occurs for many other bivalves, since many soft-bottom shores are disturbed by wave action and rainfall (Woodin 1978; Grant 1981; Berg and Alatalo 1985; Savidge and Taghon 1988; Kurihara 2002).
To adapt to such dangers, intertidal endobenthic bivalves may have evolved at least three modes. First, bivalves possibly possess morphologically and/or physiologically strong bodies. For example, the shells of some bivalves are thick enough to ward off predators and can store sufficient water to avoid desiccation (Vermeij 1973, 1978). The second mode involves being highly mobile to avoid stresses. This may be adopted by bivalves that can dig into the sediment quickly after a disturbance (Sakurai et al. 1996; Sakurai and Seto 1999). The third mode involves quick growth by which bivalves may rapidly exceed body sizes that are vulnerable to predation (Paine 1976; Vermeij 1978; Commito 1982; Boulding 1984; Eggleston 1990) and desiccation (Ricciardi et al. 1995). That is, bivalves that grow rapidly to exceed the vulnerable body size can avoid such types of stress quickly (Seed and Brown 1978).
Which of the three modes is adopted by intertidal endobenthic bivalves has been studied on some temperate and boreal shores (Seed and Brown 1978; Commito 1982; Boulding 1984; Zwarts and Wanink 1989), but rarely on subtropical shores. Compared to temperate and boreal shores, predation and desiccation appear to be more intense on subtropical shores (Vermeij 1978, 1987; Brown and Lomolino 1998). This is because, on lower latitude shores, predators have stronger weapons (e.g. crabs with large chelae and fishes with powerful jaws; Vermeij 1978, 1987); shellfish experimentally exposed to predators are more severely attacked (Zipser and Vermeij 1978; Bertness et al. 1981); sunlight is stronger; and wave splash is generally less intense, because wave action on shores is weakened by offshore coral reefs (Kurihara 2002). Therefore, to adapt to such harsh conditions, endobenthic bivalves on subtropical shores might have evolved an extremely strong body, quick mobility, or a high growth rate during their early life stages.
The present study examined which of these three modes is adopted by the Venus clams Gafrarium tumidum (Röding, 1798) and Ruditapes variegatus (Sowerby, 1852), on a subtropical cobbled shore. To compare the strength of their bodies, morphological measurements and experiments manipulating predation and desiccation stress were carried out. To compare their mobility, the digging behavior of the clams was observed. To compare their growth rates, mark–recapture surveys were conducted.
Materials and methods
Ruditapes variegatus and Gafrarium tumidum
The two clams inhabit subtropical cobbled shores around Japan (Kurihara et al. 2000). In preliminary sampling with quadrats, their densities peaked at 30–50 cm below the mean sea level (MSL) in the present study site. At the peak, R. variegatus and G. tumidum densities were approximately 30 and 10 m−2, respectively. The two clams are often consumed by the local people.
Study site
The study site was a cobbled shore (24°30′N; 124°17′E) located in Ibaruma Bay, Ishigaki Island, Okinawa. The shore extended 20 m vertically and 200 m horizontally. The shore was hard to approach and undisturbed by beachcombers. It consisted of sandy sediment and cobbles with a mean ±SD diameter of 4.6±2.5 cm (N=411). The sandy sediment often moved due to storm or terrestrial run-off (Kurihara 2002), as reported in studies of other cobbled shores (Berg and Alatalo 1985). Gastropods (buccinids, muricids and naticids), crabs (portunids and xanthids), fishes and possibly polychaetes (Kurihara et al. 2000; Kurihara, personal observation) were the main predators of Venus clams at the study site. Since most predators occur on or above the sediment surface, predation pressure appears to be highest near the sediment surface (for predation by the above-mentioned animals on Venus clams see Rodrigues 1986; Robert and Parra 1991; Saito et al. 1999).
The shore was protected by a complicated shoreline and an offshore coral reef. This leads to weak wave action (maximum wave height measured at low tide=7 cm), resulting in limited sea spray. The shore was directly exposed to sunlight. Between July 1999 and April 2001, temperatures measured nearly every month were higher at the sediment surface (mean±SD: 32.2±2.8°C during August–October; 21.0±1.1°C during January–March) than in the sandy sediment at 10 cm depth (29.2±2.3°C and 20.7±0.4°C) and in the air 1 m above the sediment (27.2±1.6°C and 19.8±1.5°C). Considering such conditions, desiccation appears to be very severe near the sediment surface.
Morphological measurements
To compare morphology between G. tumidum and R. variegatus, 30 individuals of each species were haphazardly collected near the study site in November 2001. In the laboratory, the shell length, height and width of each clam were measured with vernier calipers (±0.01 mm). After being dried at 60°C for 24 h, the individuals were weighed ("total dry weight" hereafter). They were then divided into flesh and shells, which were weighed separately ("flesh dry weight" and "shell dry weight"). The ratio of shell dry weight to total dry weight was calculated ("shell weight proportion"). After this, the thickness of one shell of each individual was measured near the umbo with vernier calipers ("shell thickness"). Finally, the weight of both shells of each individual was measured in water ("underwater shell weight"). Underwater shell weight is equivalent to [(shell dry weight) minus (shell volume)] according to Archimedes' principle. Using this relationship, underwater shell weight was subtracted from shell dry weight to estimate shell volume. Morphological characteristics that were not correctly measured on occasion due to instrument failure were omitted from the analyses.
Shell weight proportion, which was the only measurement not significantly correlated with shell length (see "Results"), was compared between species by a Mann–Whitney test (Zar 1999). The other measurements were compared by analyses of covariance (ANCOVAs), with shell length being a covariate (Zar 1999). In total, data from the same individuals were tested six times (i.e. one Mann–Whitney test and five ANCOVAs). Thus, to avoid excessive type-I errors, the significance level was lowered from the widely used 0.05 to 0.0085 (Dunn–Sidák procedure; Underwood 1997). Before these six tests were performed, 20 individuals with a shell length of 23–35 mm were re-sampled from each species. This procedure was necessary because the sample included extremely small R. variegatus (minimum: 19.4 mm) and large G. tumidum (maximum: 44.5 mm), which led to extreme, and possibly erroneous, extrapolations in ANCOVAs.
Caging experiments
Two runs of caging experiments were conducted. The experiments examined whether G. tumidum survive better than R. variegatus do in cages exposed to both predation and desiccation. Each run used lidless cages (37 cm length×25 cm width×9 cm height; made of 0.5 cm×0.5 cm resin mesh), set randomly at 40 cm below MSL and at a mean distance of 70 cm apart. The open side of cages faced upward. Half the cages were chosen randomly, and sediment from the study site was added. These cages were covered with a 0.2 cm×0.2 cm black mesh ("netted cage"). The other half of the cages were not netted and contained no sediments ("no-net cage"). Clams collected near the study site were placed into each cage. Clams were more safely protected from predation and desiccation in the netted than in the no-net cages due to the net and sediments. In run 1, two netted and two no-net cages were prepared. Ten marked G. tumidum were put into one cage of each type, and ten marked R. variegatus were placed into the remaining cages on 16 April 2002. Ten days later the survivors were counted by checking whether they closed their shells together tightly. In a preliminary experiment, it had been confirmed that clams could not emigrate from the cages. In run 2, ten netted and ten no-net cages were prepared. Six marked individuals of either G. tumidum or R. variegatus (mean±SD shell length=30.1±3.5 mm and 31.4±2.3 mm, respectively) were put into each cage on 26 April 2002 in a similar manner to run 1. Then, 13 days later, the survivors were counted. Counts were compared between cage types for each species, using a Mann–Whitney test (Zar 1999).
At the end of each run, predators at each cage were identified at low tide. During run 2, for some cages, maximum temperatures were measured by two maximum thermometers (Nihon Keiryoki Kogyo, Tokyo), and downwelling light intensity by four micro data-recorder systems for light ("MDSL"; Alec Electronics, Kobe). Half of these recorders were randomly set in netted cages, while the remaining half were placed in no-net cages. The MDSL underestimated irradiance due to fouling on the sensor and thus assessed only relative intensity of irradiance for each cage. Because a netted cage holding a MDSL with G. tumidum was broken by an animal, the data on irradiance and survival in this cage were excluded from analyses.
Predation experiments
Laboratory experiments were conducted to examine whether G. tumidum is more resistant to predation than R. variegatus is. Crabs possibly preying upon clams were caught near the study site in December 2002. They consisted of four Scylla serrata, four Thalamita sima and one Eriocheir japonicus, with carapace widths of 66–145 mm. Each crab was put into a separate aquarium (38 cm length×26 cm width×24 cm height, but 51 cm length×36 cm width×30 cm height for the largest crab) with running seawater (mean±SD: 23.6±1.0°C; N=31). Ten individuals of each clam species taken from the study site were placed in each aquarium. Within each aquarium the shell lengths of G. tumidum (mean±SD: 30.2±4.4 mm) and R. variegatus (30.0±4.7 mm) were not significantly different (t-tests: t=0.074 to 0.83, P=0.417 to 0.941). None of the aquaria held sediment, so the clams were directly exposed to the crabs. After 3 days of exposure, eaten clams were counted. The ratio of the numbers of individuals eaten versus not eaten was compared between G. tumidum and R. variegatus for each aquarium by a Fisher exact test (Zar 1999).
Desiccation experiments
Two runs of laboratory experiments were conducted to examine whether G. tumidum is more resistant to desiccation than R. variegatus is. For each run ten individuals of each clam species were collected from the study site in December 2002. They were brought into a drying oven in the laboratory. The oven was regulated to 34°C for the first run and 27°C for the second. These values approximated the annual maximum and mean temperatures, respectively, on the sediment surface in the study site. In the oven, clams were put on a saucer with no sediment, so that they were directly exposed to the dry air. After 44 h of exposure, individuals whose valves were gaping were counted. Such gaping appears to indicate damage to clams, since nine out of ten gaping clams died 1 day after being placed in an aquarium with running seawater after the experiment, whereas only one out of ten non-gaping clams died. For each run the ratio of gaping and non-gaping individuals was compared between G. tumidum and R. variegatus by a Fisher exact test.
Observations of digging behavior
Four runs of observations were made to determine whether R. variegatus digs into the sediment faster than G. tumidum does. Runs 1 and 2 were conducted at low tide on 3 November 2001 and 8 February 2002, respectively (air temperatures recorded at http://www.data.kishou.go.jp/index90.htm: 23.3°C and 19.1°C). Runs 3 and 4 were conducted at high tide on 12 and 15 December 2002, respectively (19.6°C and 23.3°C). Weather conditions during run 3 were exceptionally harsh, with heavy rain and strong wave action. For each run, 10–20 individuals of each species were collected near the study site. These individuals were placed onto the sandy sediment among cobbles at 40 cm below MSL. Individuals that extended their foot to dig into the sediment were then counted intermittently over 15 min. The count just after each run was analyzed by a Fisher exact test, which compared the ratio of the numbers of digging/non-digging individuals between R. variegatus and G. tumidum.
On the same days as runs 1 and 2, observations under natural conditions were made to examine whether R. variegatus lay on the sediment surface less frequently than G. tumidum did. For each species, living individuals whose whole body was exposed to the air were counted along a transect of approximately 200 m length at 40 cm below MSL. Whether individuals were alive or dead was determined by examining whether they closed their shells firmly.
Surveys of growth patterns
To examine whether R. variegatus grows faster than G. tumidum does, eight sets of mark–recapture surveys (four seasons×two years) were conducted between July 1999 and May 2001. For each set different individuals were collected, released and recaptured (Table 1). Both species were randomly collected from near the study site, and 86–90 individuals of each were numbered with paint in the laboratory. Their shell lengths were measured with slide calipers (±0.01 mm). For each species, individuals were divided into six groups of 14–15 individuals. Among the six groups, shell lengths were not significantly different (one-way analyses of variance; df 1= 5 to 8; df 2=84 to 126; F=0.011 to 0.098, P>0.992). Each of the 12 groups (six groups×two species) was released into 1 of 12 plots of 0.5×0.5 m2 that had been placed within a 50×1 m2 area at 40 cm below MSL. For the fourth set, three additional groups were prepared for each species (i.e. six additional groups in total), and each group was released into one of six additional plots. The positions of all plots were randomly determined, and no two plots were placed in the same position during the study period. Because plots were marked with a lone stake, marked individuals could emigrate freely. From 38 to 55 days after release, marked individuals were recaptured by sieving the sediment in each plot through 6 mm×6 mm mesh. Shell lengths of clams recaptured alive were measured again. Increase in shell length between release and recapture was divided by the number of days to estimate growth rate.
Growth rates of both species were generally smaller for larger individuals (see "Results"). Thus, growth rates were compared between species within each of 20–30 mm and 30–40 mm shell length classes. Comparisons were made only visually. None of the widely used statistic tests (e.g. ANOVA, ANCOVA) was performed, due to excessive violation of the assumptions of such tests (i.e. balanced data and homoscedasticity; Zar 1999) (P<0.05 in Cochran's tests).
Results
Morphology
Most of the morphological measurements were significantly larger in Gafrarium tumidum than in Ruditapes variegatus (Fig. 1). Shell height, shell thickness, log shell volume and log flesh dry weight significantly differed, not regarding slopes (ANCOVAs with shell length being the covariate, modified by a Dunn–Sidák procedure; P=0.015 to 0.890; see Table 2 for df 1, df 2 and F-ratios), but regarding elevation (P≤0.001). All elevations were higher for G. tumidum than for R. variegatus, except for log flesh dry weight. Shell width significantly differed regarding slope (ANCOVA; P=0.003), which made interspecific comparison of elevation difficult (Zar 1999). In the range of shell lengths examined, however, G. tumidum had a higher elevation (Fig. 1). Shell weight proportion was not significantly correlated with shell length for either species (Spearman rank correlation tests; P=0.104 to 0.203) and was significantly higher for G. tumidum (Mann–Whitney test; P=0.001).

Ruditapes variegatus and Gafrarium tumidum. Morphological measurements and regression lines for R. variegatus (filled diamonds and solid lines) and G. tumidum (open circles and dotted lines), together with regression equations and correlation coefficients (r 2). Note that no regression lines are presented for shell weight proportion because this measurement was not significantly correlated with shell length (P>0.05), as shown by the low Spearman rank correlation coefficients (ρ)
Survival in caging experiments
Environments greatly differed between cage types, at least during run 2. Daytime irradiance was more intense in the no-net cages (mean±SD: 479.6±283.2 μE m−2 s−1; N=2026) than in the netted cage (300.6±341.9 μE m−2 s−1; N=1013). The maximum temperature was higher in the no-net cage (36.0°C) than in the netted cage (30.4°C). Predators (muricid gastropods and portunid crabs) were found only around no-net cages.
During both runs 1 and 2, all G. tumidum except for one missing individual survived, whereas two to six R. variegatus died in the no-net cage (Table 3). As a result, only for R. variegatus, the median numbers of surviving individuals were lower in the no-net than in the netted cages. After each run shells of marked R. variegatus that had died were found in the vicinity of the no-net cages. Some consisted of a pair of uninjured valves with a hinge, while others were broken into pieces.
Survival in predation experiments
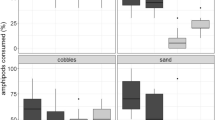
Of the nine predatory crabs, only the largest and secondly largest individuals (both Scylla serrata) foraged on clams (Table 4). Of the two S. serrata, the larger crab foraged on the same number of G. tumidum and R. variegatus, while the smaller crab ate a significantly larger number of R. variegatus (Fisher exact test; P=0.011).
Heat coma in desiccation experiments
At the end of both runs of the desiccation experiments, no G. tumidum but five R. variegatus fell into heat coma, showing a significant interspecific difference (Fisher exact test: P=0.033 for each run).
Digging behavior
During runs 1, 2 and 4, the numbers of R. variegatus digging into the sandy sediment gradually increased, while the numbers of digging G. tumidum remained low (Table 5; P≤0.011 in Fisher exact tests at the end of each run). During run 3, although there was no such significant difference (P>0.999), a greater number of R. variegatus dug into the sediment. Under natural conditions, only G. tumidum individuals were found to lie on the sediment surface: seven and nine individuals for runs 1 and 2, respectively.
Growth patterns
In all but the first survey, the median recapture rate of R. variegatus (20.0–36.7%) did not exceed that of G. tumidum (30.0–86.7%; Table 1). Small R. variegatus, with shell lengths of 20–30 mm, showed no clear difference in growth rate from small G. tumidum (Fig. 2). That is, the median growth rate of small R. variegatus was greater by >1.0 μm day−1 than that of small G. tumidum for the October 1999, April 2000 and October 2000 surveys, while the former was smaller by >1.0 μm day−1 for the July 1999, July 2000 and April 2001 surveys. For the remaining four surveys, the median growth rates of small clams did not much great interspecific difference. Large R. variegatus, with shell lengths of 30–40 mm, grew more slowly than large G. tumidum did. That is, only during the October 2000 survey was the median growth rate of large R. variegatus greater than that of large G. tumidum by >1.0 μm day−1, while for four other surveys (July 1999, April 2000, July 2000 and April 2001) the latter was greater by >1.0 μm day−1.

Ruditapes variegatus and Gafrarium tumidum. Growth rates of mark–recaptured clams with medians for shell-length classes of 20–30 mm and 30–40 mm for R. variegatus (filled diamonds and solid lines) and G. tumidum (open circles and dotted lines)
Growth rates varied not only between species, but also among seasons and between years. Generally, growth rates were the lowest in January in each year and in the second year for each season. However, the causes of such temporal fluctuations are beyond the scope of this study and will be discussed elsewhere.
Discussion and conclusions
Resistance of Gafrarium tumidum to predation and desiccation
Ruditapes variegatus is likely to be more susceptible to predation than G. tumidum is. This is suggested by the caging experiments, in which only R. variegatus survived less often under high predation pressure (i.e. no-net cage) than under low predation pressure (i.e. netted cage). Higher predation pressure in the no-net cages is reinforced by the observations that predators and pieces of shell from dead R. variegatus, probably broken by crabs and/or fishes, occurred only around the no-net cages. The vulnerability of R. variegatus to predation is also suggested by the predation experiments, in which R. variegatus were eaten by the single crab more often than G. tumidum were. Although only the medium-body-sized crab showed such prey selectivity in the predation experiments, R. variegatus is still considered to be more vulnerable to predation. The result may mean that medium-body-sized crabs can crush R. variegatus more easily than G. tumidum, while large crabs can readily crush both clam species, and small crabs cannot crush either. This is supported by the result that the largest crab ate the same number of both clam species and the third largest to the smallest crabs ate none. Such size-specific predation by crabs on clams has been observed previously (Hughes and Elner 1979; Arnold 1984; Boulding 1984).
The resistance of G. tumidum to predation is attributable to three morphological characteristics. First, its shells are globose (i.e. large in width and height), which may be more difficult for crusher-type predators to handle and crush with chela and jaw. This is suggested by the experiments of Blundon and Kennedy (1982a), in which a globose-shelled clam species was more immune to predation by blue crab than a flat-shelled species was. The second advantageous characteristic of G. tumidum is the thickness of its shells, which may make crushing and drilling more difficult for predators. This is in part suggested by the experiments of Boulding (1984), in which thicker-shelled clam species resisted a greater number of crushing behaviors of crabs. The third favorable characteristic of G. tumidum is the relatively limited amount of flesh in the shell. This increases the cost/benefit ratio (i.e. ratio of energy consumed/gained) for predators and possibly lowers the probability of predation (Charnov 1976).
In addition to predation, G. tumidum is likely to be more immune to desiccation than R. variegatus is, as indicated by the caging and desiccation experiments. The caging experiments showed that only R. variegatus died in significantly larger numbers in the no-net cages than in the netted cages. In the no-net cages desiccation may be more severe due to the stronger irradiance and higher temperature inside. In the desiccation experiments R. variegatus showed heat coma significantly more frequently than G. tumidum did.
It is unclear whether such resistance of G. tumidum to desiccation is due to morphology. The globosity and thickness of G. tumidum shell may be simultaneously advantageous and disadvantageous. The globosity decreases the ratio of shell surface area to body volume. On the one hand, this characteristic lowers the amount of gain of heat per unit volume of G. tumidum body (Porter and Gates 1969), which prevents a sharp rise in body temperature. On the other hand, shell globosity lowers the amount of heat loss, which hinders rapid cooling. The thickness of G. tumidum shells may also add to the uncertainty of whether the shell is desiccation resistant, because the thick shell may slow heat conduction between the inside and outside. Therefore, characteristics of G. tumidum other than morphology, such as possible physiological mechanisms, should be examined to explain why G. tumidum is more immune to desiccation than R. variegatus is.
Avoidance of R. variegatus from predation and desiccation
R. variegatus is more likely than G. tumidum to remain in the sediment. This is indicated from the observations that, although R. variegatus were more abundant than G. tumidum under the sediment surface (see "Materials and methods"), only G. tumidum lay on the sediment surface. The ability of R. variegatus to remain in the sediment is attributable to its quick digging behavior. This is suggested by the laboratory experiments of Sakurai and Seto (1999), in which a congeneric clam R. philippinarum was able to dig into sandy sediment quickly enough to stabilize its position when the sediment was artificially deposited and eroded. In the present study, although R. variegatus did not dig quickly in one observation at high tide, this exceptional result appears to be due to the strong wave action and low salinity during this observation. The interferences with digging behavior by these physical factors have been previously reported for R. philippinarum (Sakurai et al. 1996; Sakurai and Seto 1999).
By remaining in the sediment, R. variegatus may be able to shelter from desiccation and predation. Desiccation is milder in the sediments than on the sediment surface (see "Study site"). Predators are less abundant and diverse in the sediment (polychaete) than on and above the surface at the study site (e.g. crabs, fishes, gastropods and octopi) (Kurihara et al. 2000; Kurihara, personal observation). Predation pressure has been reported to be lower in the sediment than near the surface in many regions of the world (Blundon and Kennedy 1982b; Haddon et al. 1987; Zwarts and Wanink 1989). Staying in the sediment under cobbled shores, such as the present study site, may be particularly effective to avoid predation, since obstacles such as cobbles can protect clams from predatory attacks (Arnold 1984; Sponaugle and Lawton 1990). Overall, the ability of R. variegatus to remain in the sediment may offset its weaknesses against desiccation and predation.
The ability of R. variegatus to move quickly is attributable to the shell morphology. Its shell is less dense, which allows good mobility. Its shell is relatively flat, which facilitates movement into narrow interstices between cobbles. Further, even when dislodged from sediment by abiotic disturbances (e.g. Matthiessen 1960; Commito et al. 1995; Turner et al. 1997), the distance between the shell ventral margin and sediment surface is relatively short because of the flatness of the shell; as such, R. variegatus can touch the sediment surface effortlessly with its foot and dig quickly into the sediment.
Similarity in growth pattern between G. tumidum and R. variegatus
The weaknesses of R. variegatus against predation and desiccation are unlikely to be offset by "escaping in size" (sensu Paine 1976; see also Seed and Brown 1978; Eggleston 1990). This is indicated by the mark–recapture surveys. Small R. variegatus did not grow faster than small G. tumidum did.
It is, however, possible that R. variegatus overcomes its weaknesses by fast sexual maturity. That is, R. variegatus might increase the chances of producing offspring before death due to predation or desiccation, as reported for the bivalve Macoma balthica (Commito 1982). According to Ota and Tokeshi (2000), the oldest cohort of R. variegatus has a shell length mode of 20 mm, which suggests that R. variegatus can sexually mature before attaining this size. In contrast, Baron (1992) reported that G. tumidum can sexually mature after attained this shell length. Thus, it is possible that R. variegatus sexually matures faster than G. tumidum does, even if their growth rates are similar.
Conclusions
G. tumidum may overcome predation and desiccation through greater endurance, while R. variegatus may avoid stress through greater mobility. This interspecific difference is attributable in part to species-specific morphology. Yet, the difference may also arise from physiological mechanisms, which should be further investigated. In addition, characteristics other than endurance and mobility (e.g. the age at which they sexually mature) should also be compared between the species in the future.
References
Arnold WS (1984) The effects of prey size, predator size and sediment composition on the rate of predation of the blue crab, Callinectes sapidus Rathbun, on the hard clam, Mercenaria mercenaria (Linné). J Exp Mar Biol Ecol 80:207–219
Baron J (1992) Reproductive cycles of the bivalve molluscs Atactodea striata (Gmelin), Gafrarium tumidum Röding and Anadara scapha (L.) in New Caledonia. Aust J Mar Freshw Res 43:393–402
Berg CJ, Alatalo P (1985) Biology of the tropical bivalve Asaphis deflorata (Linné, 1758). Bull Mar Sci 37:827–838
Bertness MD, Garrity SD, Levings SC (1981) Predation pressure and gastropod foraging: a tropical–temperate comparison. Evolution 35:995–1007
Blundon JA, Kennedy VS (1982a) Mechanical and behavioral aspects of blue crab, Callinectes sapidus (Rathbun), predation on Chesapeake Bay bivalves. J Exp Mar Biol Ecol 65:47–65
Blundon JA, Kennedy VS (1982b) Refuges for infaunal bivalves from blue crab, Callinectes sapidus (Rathbun), predation in Chesapeake Bay. J Exp Mar Biol Ecol 65:67–81
Boulding EG (1984) Crab-resistant features of shells of burrowing bivalves: decreasing vulnerability by increasing handling time. J Exp Mar Biol Ecol 76:201–223
Brown JH, Lomolino MV (1998) Biogeography. Sinauer, Sunderland, Mass.
Charnov EL (1976) Optimal foraging: attack strategy of a mantid. Am Nat 110:141–151
Commito JA (1982) Effects of Lunatia heros predation on the population dynamics of Mya arenaria and Macoma balthica in Maine, USA. Mar Biol 69:187–193
Commito JA, Ambrose WG (1985) Multiple trophic levels in soft-bottom communities. Mar Ecol Prog Ser 26:283–289
Commito JA, Currier CA, Kane LR, Reinsel KA, Ulm IM (1995) Dispersal dynamics of the bivalve Gemma gemma in a patchy environment. Ecol Monogr 65:1–20
Eggleston DB (1990) Functional responses of blue crabs Callinectes sapidus Rathbun feeding on juvenile oysters Crassostrea virginica (Gmelin): effects of predator sex and size, and prey size. J Exp Mar Biol Ecol 143:73–90
Eggleston DB, Lipcius RN, Hines AH (1992) Density-dependent predation by blue crabs upon infaunal clam species with contrasting distribution and abundance patterns. Mar Ecol Prog Ser 85:55–68
Emerson CW, Grant J (1991) The control of soft-shell clam (Mya arenaria) recruitment on intertidal sandflats by bedload sediment transport. Limnol Oceanogr 36:1288–1300
Grant J (1981) Sediment transport and disturbance on an intertidal sandflat: infaunal distribution and recolonization. Mar Ecol Prog Ser 6:249–255
Haddon M, Wear RG, Packer HA (1987) Depth and density of burial by the bivalve Paphies ventricosa as refuges from predation by the crab Ovalipes catharus. Mar Biol 94:25–30
Hilgerloh G (1997) Predation by birds on blue mussel Mytilus edulis beds of the tidal flats of Spiekeroog (southern North Sea). Mar Ecol Prog Ser 146:61–72
Hines AH, Whitlatch RB, Thrush SF, Hewitt JE, Cummings VJ, Dayton PK, Legendre P (1997) Nonlinear foraging response of a large marine predator to benthic prey: eagle ray pits and bivalves in a New Zealand sandflat. J Exp Mar Biol Ecol 216:191–210
Hughes RN, Elner RW (1979) Tactics of a predator, Carcinus maenas, and morphological responses of the prey, Nucella lapillus. J Anim Ecol 48:65–78
Jangoux M (1982) Food and feeding mechanisms: Asteroidea. In: Jangoux M, Lawrence JM (eds) Echinoderm nutrition. Balkema, Rotterdam, pp 117–159
Kurihara T (2002) Spatial and temporal fluctuation in the density of the intertidal limpet, Patelloida striata Quoy & Gaimard, on subtropical cobbled shores. J Moll Stud 68:83–90
Kurihara T, Takada Y, Kosuge T, Kobayashi M, Katoh M, Mito K (2000) Species composition of epifauna and infauna on intertidal boulder shores at Ishigaki Island in subtropical Japan. Bull Seikai Natl Fish Res Inst 78:31–47
Leonard GH, Bertness MD, Yund PO (1999) Crab predation, waterborne cues, and inducible defenses in the blue mussel, Mytilus edulis. Ecology 80:1–14
Matthiessen GC (1960) Intertidal zonation in populations of Mya arenaria. Limnol Oceanogr 5:381–388
Ota N, Tokeshi M (2000) Population analysis of Ruditapes variegatus (Sowerby) (Bivalvia: Veneridae) on an intertidal boulder shore. Venus Jpn J Malacol 59:29–36
Paine RT (1976) Size-limited predation: an observational and experimental approach with the Mytilus–Pisaster interaction. Ecology 57:858–873
Porter WP, Gates DM (1969) Thermodynamic equilibria of animals with environment. Ecol Monogr 39:227–244
Ricciardi A, Serrouya R, Whoriskey FG (1995) Aerial exposure tolerance of zebra and quagga mussels (Bivalvia: Dreissenidae): implications for overload dispersal. Can J Fish Aquat Sci 52:470–477
Robert R, Parra R (1991) Experimental study of predation by the gilthead bream Sparus aurata and the gray triggerfish Balistes capriscus on the Manila clam Ruditapes philippinarum (in French with English abstract). Aquat Living Resour 4:181–189
Rodrigues CL (1986) Predation of the naticid gastropod Neverita didyma (Röding) on the bivalve Ruditapes philippinarum (Adams & Reeve): evidence for a preference linked functional response. Publ Amakusa Mar Biol Lab Kyushu Univ 8:125–141
Saito H, Imabayashi H, Kawai K (1999) Growth of the bivalve-feeder Halla okudai (Polychaeta: Lysaretidae) under wild and rearing conditions, in relation to species and abundance of prey organisms. Fish Sci (Tokyo) 65:230–234
Sakurai I, Seto M (1999) Behavioral characteristics of the juvenile Japanese littleneck clam Ruditapes philippinarum in response to sand erosion and deposition (in Japanese with English abstract). Sci Rep Hokkaido Fish Exp Stn 54:41–46
Sakurai I, Seto M, Nakao S (1996) Effects of water temperature, salinity and substrata on burrowing behaviors of the three bivalves, Pseudocardium sachalinensis, Mactra chinensis, and Ruditapes philippinarum (in Japanese with English abstract). Nippon Suisan Gakkaishi 62:878–885
Savidge WB, Taghon GL (1988) The influence of passive advection on the colonization of two types of disturbance on an intertidal sandflat. J Exp Mar Biol Ecol 115:137–155
Seed R, Brown RA (1978) Growth as a strategy for survival in two marine bivalves, Cerastoderma edule and Modiolus modiolus. J Anim Ecol 47:283–292
Seitz RD, Lipcius RN, Hines AH, Eggleston DB (2001) Density-dependent predation, habitat variation, and the persistence of marine bivalve prey. Ecology 82:2435–2451
Sponaugle S, Lawton P (1990) Portunid crab predation on juvenile hard clams: effects of substrate type and prey density. Mar Ecol Prog Ser 67:43–53
Turner SJ, Grant J, Pridmore RD, Hewitt JE, Wilkinson MR, Hume TM, Morrisey DJ (1997) Bedload and water-column transport and colonization processes by post-settlement benthic macrofauna: does infaunal density matter? J Exp Mar Biol Ecol 216:51–75
Underwood AJ (1997) Experiments in ecology. Cambridge University Press, Cambridge, UK
Vermeij GJ (1973) Morphological patterns in high-intertidal gastropods: adaptive strategies and their limitations. Mar Biol 20:319–346
Vermeij GJ (1978) Biogeography and adaptation. Harvard University Press, Cambridge, Mass.
Vermeij GJ (1987) Evolution and escalation: an ecological history of life. Princeton University Press, Princeton, N.J.
Woodin SA (1978) Refuges, disturbance, and community structure: a marine soft-bottom example. Ecology 59:274–284
Zaklan SD, Ydenberg R (1997) The body size-burial depth relationship in the infaunal clam Mya arenaria. J Exp Mar Biol Ecol 215:1–17
Zar JH (1999) Biostatistical analysis, 4th edn. Prentice Hall, Engelwood Cliffs, N.J.
Zipser E, Vermeij GJ (1978) Crushing behavior of tropical and temperate crabs. J Exp Mar Biol Ecol 31:155–172
Zwarts L, Wanink J (1989) Siphon size and burying depth in deposit- and suspension-feeding benthic bivalves. Mar Biol 100:227–240
Acknowledgements
I am obliged to Drs. Y. Shimadzu, Y. Takada, S. Wada and an anonymous referee for their constructive comments regarding this manuscript. I thank Mrs. Y. Hosokawa for her help with the measurement of samples. I am indebted to the staff of the Ishigaki Tropical Station for their advice and assistance during the study. The present study was supported in part by the Fisheries Research Center Seeds Research Program. The experiments in the present study comply with the current laws of Japan.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by T. Ikeda, Hakodate
Rights and permissions
About this article
Cite this article
Kurihara, T. Adaptations of subtropical Venus clams to predation and desiccation: endurance of Gafrarium tumidum and avoidance of Ruditapes variegatus . Marine Biology 143, 1117–1125 (2003). https://doi.org/10.1007/s00227-003-1158-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00227-003-1158-9