
Abstract
Effects of food availability and season on the free amino acid (FAA) and total protein content of the copepod Calanus finmarchicus females were investigated in two mesocosm experiments on the Norwegian west coast in spring and autumn. Starved C. finmarchicus females showed no change in total FAA content, but the FAA pool composition changed drastically. During the first 10 days of starvation the protein content showed a moderate decline (<2 μg ind−1); however, during the following 21 days the total content was drastically reduced, from 63 to 9 μg ind−1. This supports the notion of a sequential catabolism of endogenous nutrients during starvation. In females at high food concentrations, the body protein content increased during spring, but decreased during autumn. The FAA pool composition of females differed between spring and autumn in 14 of the 18 FAA investigated. Reduced body protein content and increased proportion of essential free amino acid were observed during starvation. Similar changes were observed in females sampled at the end of the mesocosm experiments in the autumn. The results suggest that mature C. finmarchicus females are in a negative protein balance during autumn, despite high food concentrations, contributing to a lower fitness than in females maturing during the spring.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Little is known about environmental effects on the protein and free amino acid (FAA) content in common marine zooplankton such as calanoid copepods. In general, the balance between supply and biosynthesis on the one hand, and osmotic equilibrium and catabolism on the other hand, determines the level of the FAA pool in marine organisms (Yancey et al. 1982). In certain circumstances, energy requirements can be met by catabolism of endogenous proteins, but so far no storage protein has been found, although it has been hypothesised that myofibrillar protein serves such a role in fishes (Mommsen 2001). The non-selective use of endogenous protein for energy will be detrimental to the animal, in the long run. Because mobilised proteins may include enzymes and other vital organic molecules, their degradation will have far-reaching effects on metabolic functions. Intracellular FAA plays a key role in the osmoregulation of planktonic organisms (Schoffeniels 1976; Yancey et al. 1982), where increased ambient salinity results in enhanced FAA content (Fyhn 1976; Goolish and Burton 1988; Dalla Via 1989; Fyhn et al. 1993; Helland et al. 2000). Guisande et al. (2000) showed that the female body amino acid composition was unaffected by the diet composition. However, to our knowledge there has been no study in which the FAA and protein content in common zooplankton such as Calanus finmarchicus has been examined in relation to the combined effects of diet and season.
In Norwegian waters and shelf seas, the life history of C. finmarchicus is considered to vary between one and three generations per year, where the number of generations decreases from low to high latitudes. A two-generation pattern is found around the British Isles, Faeroe Islands and even up to the Western Coast of Norway (Østvedt 1955; Wiborg 1955) and Lofoten is often referred to as the northernmost area where this two-generation cycle occurs (Tande 1991). C. finmarchicus is known to complete one or several annual generations, but the major reproduction period is confined to spring (e.g. Hirche 1996a; Båmstedt 2000, and references therein). It is also known that the developmental stage of overwintering may vary with latitude, and that feeding is depressed during the prolonged overwintering period (e.g. Hirche 1996b; Båmstedt 2000; Pasternak et al. 2001). Females reproducing in spring represent the major contributors to population recruitment, while females reproducing in autumn only contribute to a small fraction of the next generation (Båmstedt 2000). Theoretically, offspring from the first group would have good chances of developing, whereas offspring from the second group are less likely to develop to adulthood, due to inferior nutritional and environmental conditions. In the present study we compare the FAA and protein contents in C. finmarchicus females during spring and autumn, and during starvation in the autumn.
Materials and methods
The mesocosms
Two series of seawater enclosure experiments were conducted at the Marine Biological Field Station at Espegrend, University of Bergen, Norway between 17 April 1998 and 12 May 1998 (hereafter, spring), and between 11 August 1998 and 29 August 1998 (hereafter, autumn). The nutrients within the mesocosms were manipulated (Table 1) in order to produce contrasting feeding environments of natural plankton dominated by diatoms and flagellates, respectively (compare results from previous enclosure experiments, e.g. Egge and Aksnes 1992; Egge 1993; Egge and Jacobsen 1997).
The mesocosms were 27 m3 (2 m diameter, 9.25 m deep) and made of transparent polyethylene with a penetration allowing 90% photosynthetically-active radiation . They were filled in situ by pumping water from 5 m depth. In order to stratify the water column, the mesocosms were topped up with about 0.6 m3 freshwater, and the upper 4 m (12.5 m3) of the water column was mixed and kept homogenous with air-lifts throughout the experiment. This created a salinity difference between upper and lower layers of 0.19±0.08 g l−1 in the spring, and 1.35±0.07 g l−1 (where values are mean±SD) in autumn. The upper 4 m were well mixed. This has been shown in previous mesocosm experiments with the same circulation system (Williams and Egge 1998), and was again confirmed by completely linear depth profiles of the salinity, temperature and fluorescence in the mixed layer 24 h after the onset of the circulation (unpublished data). Nutrients were added to the upper layers as shown in Table 1. The original names of the different mesocosms are used (Table 1) in order to facilitate comparisons with other related studies (Svensen et al. 2002; Helland et al. 2003a). For further description of the mesocosm experimental design see Svensen et al. (2001, 2002); a general description of the mesocosm facility is also available at http://www.ifm.uib.no/LSF/inst2.html.
Salinity and temperature profiles (0–8 m) were obtained with a Sea Cat Profiler (Sea-Bird Electronics, Bellevue, Wash., USA). Samples for nutrients, chlorophyll (Chl) a, particulate organic carbon (POC) and nitrogen (PON) and protists were taken using a Ruttner water bottle (at 2 m and 6 m). This was done at day one, and every third day until termination of the enclosure experiments. Sampling of water from the mesocosms for determination of FAA and protein in the particulate organic matter (POM) was done by filtration (Nalgene suction filter system) on pre-weighed glass fibre filters (Whatman GF/F) or 90 μm plankton net filters after filtering through a 200 μm net in order to remove mesozooplankton. The filter was transferred to pre-weighed Nunc cryo tubes, and the samples were frozen on dry ice then stored at −80°C until later lyophilisation. Females of C. finmarchicus were collected from the mesocosms by vertical tows from 9 m to the surface using a 30 cm diameter 90 μm-mesh net. Females were individually identified using a dissecting microscope and the prosome length measured to the nearest 0.02 mm. Only adult females of stage CVI were used in the experiments. For biochemical analyses, individual females were transferred to Nunc cryo tubes and frozen on dry ice. Egg diameter measurements and visual inspection of the females, showed egg diameters (mean±SD) of 145±6 μm (n=804) in spring and 150±6 μm (n=63) in autumn. This is in agreement with the expected diameter of eggs from C. finmarchicus, in contrast to the occasionally co-occurring sibling species Calanus helgolandicus (Marshall and Orr 1955), and confirmed our taxonomical determination of C. finmarchicus based on morphology (Jaschnov 1972; Frost 1974; Svensen and Tande 1999, and references therein).
Starvation experiment
Females of C. finmarchicus were caught in the fjord surrounding the mesocosms, with short (less than 20 m) oblique tows (1 m diameter net, 500 μm mesh, 14 l non-filtering codend). The content of the codend was immediately diluted with surface water (avoiding animals on the net), brought to a walk-in cold room with dim light at 6.0±1.0°C (mean±SD). After dead and injured animals had settled for 30 min, actively swimming and intact females were sorted under a dissecting microscope, and randomly assigned to six groups of eight females each. Only adult females of stage CVI were used in the experiments. Each group was transferred to a 500 ml beaker containing 0.2 μm-filtered seawater. The beakers had an inner chamber of acrylic plastic with a false bottom of plankton net (500 μm mesh) in order to separate the females from faecal pellets and facilitate water renewal (every second day). Sampling was conducted six times (days 1, 4, 10, 18, 31, and 39), and after measurement of prosome length, individual females were transferred to Nunc cryo tubes and immediately frozen on dry ice.
Analyses of samples
Nitrate, phosphate and silicate concentrations were determined on fresh samples using a Skalar SANplus segmented flow analyser. Chl a samples were filtered onto Sartorius filters (0.45 μm) and analysed on a Turner Designs Model 10-AU Fluorometer according to Parsons et al. (1984). POC and PON samples were filtered onto pre-combusted Whatman GF/F glass-fibre filters and analysed on a Leeman Lab CEC 440 elemental analyser after removal of carbonate with fumes of concentrated HCl. Samples for analysis of protein and FAA were extracted in 6% tri-chloro-acetic acid (TCA) for 24 hours, and centrifuged for 10 min at 5,000 g. The supernatant was used for analysis of FAA according to Helland et al. (2000), while the precipitated material was used for determination of protein by the method of Lowry et al. (1951). After centrifugation the precipitate was rinsed once with 6% TCA, to remove any residual FAA, then solubilised in 1.0 M NaOH and further diluted to 0.5 M before analysis. The term essential free amino acids (EAA) is used to denote the same amino acids as identified by Guillaume (1997) with the addition of tyrosine considered as semi-essential (Guillaume 1997). Phytoplankton samples were preserved with a glutardialdehyde-lugol mix (35‰ final v/v) (Rousseau et al. 1990). Microzooplankton samples were fixed in acid Lugol's solution (final concentration 1%). Samples were settled, counted and sized (using an ocular micrometer) as described in Nejstgaard et al (2001a). Cell volume was converted into C according to the equations by Putt and Stoecker (1989) for aloricate ciliates, and according to Menden-Deuer and Lessard (2000) for all other protist plankton, as described in Nejstgaard et al. (2001a).
Statistics
Changes in FAA during starvation of C. finmarchicus was tested by analysis of variance (ANOVA) followed by Tukey's multiple comparison test for unequal n when appropriate. For testing the interrelationship between FAA composition of POM and C. finmarchicus females, linear regression was employed. The FAA compositions were sorted so that each amino acid in POM was tested against the same amino acid in C. finmarchicus females. This method for testing of interrelationships has also been employed by Tulli and Tibaldi (1997) and Aragão et al. (2001). Since arginine is a major EAA compared to the others, we tested the impact of this amino acid on linear regressions by performing them with and without arginine. This did not change the conclusions drawn from the data, and we therefore used all EAA in the tests of interrelationship. Differences between spring and autumn samples in protein, FAA and EAA versus prosome length, and the ratio of FAA to protein, were tested using the Mann-Whitney U-test (Zar 1996). Variations in salinity, temperature, POC, PON, carbon to nitrogen ratio, Chl a, protist C content, and C. finmarchicus females' protein and FAA content and composition were tested within and between seasons with one way ANOVAs, followed by Tukey's test, as above (Zar 1996). A 95% significance level was employed throughout.
Results
Starvation
During the first 10 days of starvation of female C. finmarchicus, the protein content declined moderately (1.7 μg ind-1); however, at the end of the experiment, the total content was drastically reduced, from about 63 to about 9 μg ind-1 (Fig. 1). Seven females were analysed for protein content at day 8 of starvation and the data coefficients of variation were 27% and 23% when the protein was calculated as μg protein female−1 and μg protein mm prosome−1, respectively.

Content (nmol female−1) of total free amino acids (FAA) where values are mean±SD (n=17) and of protein (μg female−1) where values are mean±SD (n=25) in Calanus finmarchicus females during 39 days of starvation. A best fit was made to the protein content dataset. For points marked with an asterisk, n=2
The total individual FAA content fluctuated during the starvation experiment (Fig. 1), and there was no significant trend in FAA content during starvation (linear regression, r 2=0.02). Calculation of FAA content relative to prosome length mimicked the results of individual content, and the coefficient of variation was similar (24.7% and 25.7%, respectively). The composition of the FAA pool changed throughout the starvation experiment, where EAA accounted for 36% and 52% of total FAA at start and end, respectively. The changes in FAA composition during starvation were calculated and the eight FAA that changed most, between 7 and 24 nmol female−1, are presented in Fig. 2. Of these eight FAA, three were EAA and five were non-essential amino acids (NEAA) and together they contributed 80% and 60% of total FAA at start and end of the experiment, respectively.

Changes (nmol female−1) in FAA content in Calanus finmarchicus females from start and during 39 days of starvation; values are means for n=17. The eight amino acids that changed most are presented
Mesocosm hydrography and plankton content
The average salinity and temperature was not different between the mesocosms within each experiment (ANOVA, P<0.05). However, the average temperature in the spring was 7.4±0.6°C while it was 14.1±1.0°C in autumn. The salinity also differed between spring (31.8±0.1 g l−1) and autumn (30.1±0.4 g l−1) (ANOVA, P<0.05). There was no significant difference in abundance of particulate material between silicate and non-silicate treatments within the seasons (ANOVA, P=0.13, Table 2). However, the diatom abundance tended to be lower, but not significantly, in all non-silicate treatments (mesocosms Z1, Z2, Z3, 1 and 4), compared to the silicate-fertilised mesocosms (ANOVA, P=0.06, Table 2). The autumn mesocosms (1-S4) showed significantly higher concentrations of large flagellates, POC and carbon to nitrogen ratio (ANOVA, P≤0.01) but not ciliates (ANOVA, P=0.07), while the small flagellates were more abundant in the spring mesocosms (ANOVA, P<0.0001, Table 2). The Chl a concentrations did not differ between spring and autumn (t-test, P=0.06), while the abundance of flagellates <10 μm was higher in the spring (t-test, P<10−7, Table 2). The large flagellates were dominated by Gyrodinium spp. in both seasons, large forms of Apedinella spinifera in the spring and Ceratium spp. in the autumn. The smaller flagellates were dominated by single celled A. spinifera and Phaeocystis pouchetii in the spring and unidentified species between 3 and 5 μm diameter in the autumn. Ciliates were mostly aloricate species (Strombidium and Strombilidium spp.) dominated by the 30–90 μm size range.
FAA in the POM in the mesocosms
At the end of the spring experiment, the EAA in the POM ranged from 19 to 34% of total FAA, while the range was 17–41% in the autumn (Table 3). The dominating EAA varied between the different POM fractions in the mesocosms. Valine, leucine and threonine were highest in most cases, and arginine dominated the EAA pool in six cases, while histidine, methionine and tyrosine accounted only for a minor fraction of the pool (trace to 4.3%, Table 3). The dominating NEAA of the POM also differed between seasons. In spring, most of the 0.45–90 μm fractions of the POM had high proportion of glutamate, alanine and aspartate, while most of the 90–200 μm fraction showed high glycine, serine, proline and taurine concentrations. In the autumn, glycine was the major NEAA except for the POM of mesocosm S4 where glutamate and proline dominated. Apparently, depending upon the selected diet, the female may show large differences in the intake of the various FAA.
Copepod protein content
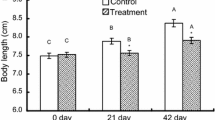
No difference in protein content was found between females sampled at the start of the experiment in spring and in autumn, nor was any difference found between females given the different POM within a season (ANOVA, between start of seasons P=0.16, within spring P=0.15 and within autumn P=0.57). However, after 21 days in the spring the protein content was higher than at the start (ANOVA, n=28, P<0.0002) and after 18 days in the autumn it was lower (ANOVA, n=22, P<0.002). The protein content after 21 days in spring was higher than after 18 days in autumn (ANOVA, n= 50, P<0.00001). The copepods had longer prosomes in the spring than in the autumn, 2.53±0.18 mm ind−1 and 2.41±0.11 mm ind−1, respectively (t-test, n=51, P<0.01). Also when protein content (μg ind−1) was standardised for prosome length (mm ind−1, Fig. 3), females had significantly higher protein content in the spring than in autumn (U-test, n=51, P<0.001, Fig. 3).

Protein content (μg ind−1) versus prosome length (mm) in Calanus finmarchicus females after 21 and 18 days, respectively, in mesocosms during the spring and autumn of 1998
Copepod FAA content
Values of FAA and EAA versus prosome length were higher for females sampled in the autumn compared to spring (Fig. 4) and differences were found for values of EAA versus prosome length (U-test, n=38, P <0.05), but not for total FAA versus prosome length. Values of FAA versus protein at the start of the experiments were slightly higher in autumn than in spring, although not significantly. In contrast, at the end of the autumn experiment higher values were found for FAA versus protein (U-test, n=38, P<0.001). No within-season differences were found between the start and the end for the FAA pool profile (% of total) comparing females fed the different POM (Table 4, P >0.05). However, the composition of amino acids differed between spring and autumn samples for 14 of the 18 amino acids investigated (Table 4, ANOVA, n=41, P<0.05).

Content (nmol ind−1) of total FAA and essential free amino acids (EAA) versus prosome length (mm) in Calanus finmarchicus females after 21 and 18 days, respectively, in mesocosms during the spring and autumn of 1998
Relationship between FAA in POM and copepods
Effects of food suspensions and seasons on the amino acid composition of females in the different food environments were further investigated by performing a series of linear regressions (Table 5). During spring the food suspensions from the mesocosms were fractionated. In mesocosms Z3, SZ1 and SZ3, there was an interrelationship between the EAA composition of the 0.45–90 μm fraction of the POM and that of the females. Similarly, the 90–200 μm fraction showed an interrelationship in EAA composition of females with all POM except that in mesocosm Z3. In the autumn, the EAA composition of the POM from mesocosms S1, S4, and 4 and their respective females showed an interrelationship. An interrelationship between POM and females' NEAA composition was only found for the POM of mesocosm SZ2 (spring), and mesocosms S1 and 4 (autumn).
Discussion
Starvation
During the first 10 days of starvation C. finmarchicus showed no significant change in total FAA, and the protein showed only a small decline. However, during the reminder of the experiment the total protein content declined dramatically, from 63 to 9 μg ind−1. This fast reduction which started at day ten and had mostly ended at day 31, is equivalent to a protein loss of 2.6 μg ind−1 day-1. Cowey and Corner (1963) studied the losses of amino acids by adult female Calanus during starvation experiments conducted in winter and in summer. They show that the average dry weight of individuals in winter (at 7°C) was 109 μg at the start and 84 μg after 14 days starvation, and in summer (at 12°C), 200 μg at the start and 123 μg after 10 days of starvation. Their calculated loss of total amino acids is 1.5 and 4.1 μg ind−1 day-1 in winter and summer, respectively. Thus, the results of Cowey and Corner (1963) are comparable to the results of our starvation experiments performed in autumn at 6°C.
Taking a molecular weight of protein amino acid of 110 Da and an average nitrogen content of 1.3 mol N mol−1 amino acid (Rønnestad et al. 1992), the 2.57 μg ind−1 day-1 protein reduction corresponds to a reduction of 30.3 nmol N day−1. Ikeda et al. (2001) presented an ammonia excretion of 0.013 µg N ind−1 h−1 for C. finmarchicus females, giving a total excretion loss of 22.3 nmol N ind−1 day−1. Calculating the ammonia excretion of Ikeda et al. (2001), using a Q10 of 2.0 (Torres et al. 1994) gives an excretion loss of 33.6 nmol N ind−1 day−1 (6°C). The protein reduction found in the present paper is slightly lower than that of Ikeda et al. (2001), but their value may also include N from other N containing compounds that were not measured in the present study (e.g. nucleic acids).
The fact that reduction of protein lagged in the present study may reflect a sequential biochemical breakdown of the animal. Such a sequential breakdown is reported by Mayzaud (1976) for starving C. finmarchicus, where a repetitive switch between a primarily protein catabolism to a primarily lipidic catabolism was observed. Initially, during starvation the lipid store is preferably used as energy for the basal metabolism (Båmstedt and Holt 1978), and later, when this store is exhausted, the hydrolysis of proteins and breakdown of structural lipids may commence. Helland et al. (2003b) found no lag in protein reduction during starvation of Temora longicornis, a copepod with negligible wax ester content (Fraser et al. 1989). C. finmarchicus females contain a considerable amount of storage lipids in the form of wax esters (Kattner and Krause 1987; Fraser et al. 1989; Hygum et al. 2000). Wax esters formation is a rapid process limited by the amount of free fatty alcohols (Sargent et al. 1976), and the resulting energy-rich wax ester reserves are mobilised during unfavourable food conditions (Lee et al. 1970; Lee 1974). With regard to the fatty acid/alcohol composition, it appears that the shorter-chain wax esters are preferably catabolised, and the wax esters with the highest energy content are stored longer (Kattner and Krause 1987). Furthermore, Mayzaud (1976) found that C. finmarchicus catabolised their lipids preferentially, both on an absolute and relative basis. The sparing of protein that is proposed in the present paper and the sequential breakdown that was found by Mayzaud (1976) may conserve body proteins. Such a sparing of proteins will enhance the resistance of the animal to starvation, and is of fundamental importance since use of endogenous protein for energy will ultimately be destructive to the animal.
Typically, in marine copepods the FAA pool amounts to 15–30% of the proteinic amino acid content (Helland et al. 2003b), 5 to 9% of dry mass (Jeffries 1969; Jeffries and Alzara 1970) and 1.6% of wet mass (Naess et al. 1995). The EAA make up 20% and 45% of the total FAA pool (Jeffries and Alzara 1970; Fyhn et al. 1993; Naess et al. 1995; Helland et al. 2003b). In the present study, there was an increase in the proportion of EAA and a coupled decrease in NEAA in the FAA pool. We propose that a simple dilution model can explain this shift. Hydrolysis of proteins during starvation will release amino acids in the same EAA/NEAA ratio as in the proteins. In the present study no increase was found in the total FAA content, and, due to stable salinity in rearing, none was expected. By this mechanism, the catabolism equals the hydrolysis unless FAA is excreted. As long as the ratio of EAA is higher in the protein than in the free pool, a simple dilution explains the increase in EAA versus NEAA in the free pool. An ideal increase and decrease in all individual EAA and NEAA in the free pool is not to be expected since the protein's amino acid composition differs from that of the free pool. In addition, a sparing of some essential amino acids is likely, due to differing rates of catabolism of the various amino acids.
Mesocosms
The C. finmarchicus females sampled at the end of the spring experiment showed a high protein, low FAA, and low EAA content and longer prosomes, compared to females sampled in the autumn. The prosome length results are in accordance with Båmstedt (2000), who found that females sampled in the upper water column (0–50 m) in Skagerrak coastal waters had longer prosomes in March–April and shorter in July–August. In the present study, it is likely that the females sampled at the end of the spring experiment entered the mesocosms at stage CV and successively developed to stage CVI during the experimental period. Hygum et al. (2000) found an increase in growth rates from stage CI to adults in C. finmarchicus, and that the copepod stages had a larger body size and an increased amount of carbon, nitrogen and lipids when the authors enhanced food resources by nutrient addition. Therefore, the present results suggest that spring females had a better food availability and/or a better nutrient retention than autumn females.
In the autumn the copepods were exposed to higher concentrations of larger prey such as ciliates, larger flagellates and diatoms, compared to the more abundant smaller flagellates in the spring mesocosms (Table 2). Grazing experiments performed with Calanus spp. in parallel with the spring experiment (Nejstgaard et al. 2001b), and a number of previous mesocosm experiments at this facility (Nejstgaard et al. 1994, 1997, 2001a) have shown a clear preference for larger prey such as ciliates and other microzooplankton up to several hundreds of μm size. This is in accordance with the literature (see reviews by Kleppel 1993; Hansen et al. 1997; Levinsen et al. 2000). While small flagellates and diatoms generally were not selected as food, diatoms often made up the bulk of the diet when abundant in the food suspension (Nejstgaard 1997, 2001a). Considering that the Chl a concentrations were always above 2.5 μg l−1, the potential prey C was above 200 μg l−1 in all suspensions except for mesocosm S4 (Table 2), and that the concentration of preferred copepod prey (larger flagellates, diatoms and ciliates, cf. Nejstgaard et al. 2001a) were at least as high in autumn as in spring, food limitation seems unlikely (cf. e.g. Ohman and Runge 1994; Båmstedt et al. 1999; Campbell et al. 2001). We therefore assume that the copepods did not experience a lower concentration of generally preferred food during the autumn experiment.
The FAA pool of the various food suspensions was compared with that of the females in order to test whether diet FAA pool composition influences female FAA pool composition (Table 5). Comparing the pool EAA compositions, a smaller interrelationship was found for the 0.45–90 μm fractions than for the 90–200 μm fractions (Table 5). This could be expected, since the 0.45–90 μm fractions contain mostly algae and protozoa, while the larger fractions contain mostly metazoa, which are expected to have a composition more similar to that of the females, because of their higher taxonomical and trophic position (Klein Breteler et al. 1990, 1999). The amino acid taurine is a degradation product of sulphur-amino acid metabolism in animals; it is not present in phytoplankton and detritus (Cowey and Corner 1963). Taurine, therefore, may be an index of the relative amounts of plant and animal components of the POM. We neither found a difference between the FAA composition of females at the start or end of each mesocosm experiment, nor between females offered the various POM within each experiment, even though there were large differences in FAA composition of the various POM both within seasons and between seasons (Table 3). We therefore conclude that diet FAA pool does not alter the female copepod FAA pool composition. Even though the POM differed in FAA composition within season, the FAA composition in the females was similar, so a regulation of female FAA composition seems likely. Such a homeostasis in amino acids composition has previously been shown for other marine copepods, controlling the proteinic pool (Guisande et al. 1999) and the free pool (Helland et al. 2003b).
The EAA fraction of the total FAA pool in C. finmarchicus females was clearly lower in spring (25%), compared to in autumn (40%). This difference between spring and autumn females was also seen after plotting FAA and EAA content versus prosome length, showing higher and more varying amounts in autumn than in spring females. It is known that the FAA content increases with increasing salinity, and that the amino acids that increase are mainly the NEAA (Fyhn 1976; Fyhn et al. 1993; Helland et al. 2000). In the present study however, the FAA and EAA content was highest in the autumn when the mesocosm salinity was lower, although only by 5%. In addition, the body protein content of the females was lowest during the autumn. A reduced content of protein and an increased EAA composition was also found in the starvation experiment, suggesting that the females sampled from the mesocosms at the end of the autumn experiment were in a negative nitrogen balance. The reduction in protein was probably due to inadequate intake of amino acids, combined with use of nitrogenous compounds for energy. Further studies, including body water and lipid contents, should be performed to reveal the physiological processes involved in the found differences in the pool of FAA of C. finmarchicus females in spring and autumn.
Conclusions
This is the first study of C. finmarchicus females in which the FAA and protein content has been examined in relation to the combined effects of diet and season. Starvation resulted in lower body protein content and increased proportion of essential amino acids in the FAA pool. Our finding of a lag in protein reduction during starvation supports the notion of a sequential catabolism of endogenous nutrients in C. finmarchicus females. Furthermore, the lower protein and higher EAA content in autumn than in spring females indicates a nitrogen limitation by the diet. The degree of interrelationship in EAA and NEAA between diets and female in the present study are not an adequate indicator for nutritional quality of POM per se. It may, however, indicate similarity between prey and predator, and whether the POM has a predominantly animal or algae FAA composition.
References
Aragão C, Conceição LEC, Fyhn HJ, Dinis MT (2001) Amino acid profiles of sea bream (Sparus aurata) and sole (Solea senegalesisi) during first feeding: are currently used diets imbalanced in amino acids? In: Hendry CI, Van Stappen G, Wille M, Sorgeloos P (eds) Special publication number 30. European Aquaculture Society, Oostende
Båmstedt U (2000) Life cycle, seasonal vertical distribution and feeding of Calanus finmarchicus in Skagerak coastal water. Mar Biol 137:279–289
Båmstedt U, Holt MR (1978) Experimental studies on the deep-water pelagic community of Korsfjorden, western Norway. Sarsia 63:225–236
Båmstedt U, Nejstgaard JC, Solberg PT (1999) Utilisation of small-sized food algae by Calanus finmarchicus (Copepoda, Calanoida) and the significance of feeding history. Sarsia 84:19–38
Campbell RG, Runge JA, Durbin EG (2001) Evidence for food limitation of Calanus finmarchicus production rates on the southern flank of Georges Bank during April 1997. Deep-Sea Res II 48:531–549
Cowey CB, Corner EDS (1963) On the nutrition and metabolism of zooplankton. II. The relationship between the marine copepod Calanus helgolandicus and particulate material in Plymouth Sea water, in terms of amino acid composition. J Mar Biol Assoc UK 43:495–511
Dalla Via GJ (1989) The effect of salinity on free amino acids in the prawn Palaemon elegans (Rathke). Arch Hydrobiol 115:125–135
Egge JK (1993) Nutrient control of phytoplankton growth: effects of macronutrient composition (N, P, Si) on species succession. Ph.D. thesis, University of Bergen
Egge JK, Aksnes DL (1992) Silicate as regulating nutrient in phytoplankton competition. Mar Ecol Prog Ser 83:281–289
Egge JK, Jacobsen A (1997) Influence of silicate on particulate carbon production in phytoplankton. Mar Ecol Prog Ser 147:219–230
Fraser AJ, Sargent JR, Gamble JC (1989) Lipid class and fatty acid composition of Calanus finmarchicus (Gunnerus), Pseudocalanus sp. and Temora longicornis Muller from a nutrient enriched seawater enclosure. J Exp Mar Biol Ecol 130:81–92
Frost BW (1974) Calanus marshallae, a new species of calanoid copepod closely allied to the sibling species C. finmarchicus and C. glacialis. Mar Biol 26:77–79
Fyhn HJ (1976) Holeuryhalinity and its mechanisms in a cirriped crustacean, Balanus improvisus. Comp Biochem Physiol 53A:19–30
Fyhn HJ, Finn RN, Helland S, Rønnestad I, Lømsland ER (1993) Nutritional value of phyto- and zooplankton as live food for marine fish larvae. In: Reinertsen H, Jørgensen L, Tvinnereim K (eds) Proceedings of the first international conference on fish farming technology. Balkema, Rotterdam, pp 121–126
Goolish EM, Burton R (1988) Exposure to fluctuating salinity enhances free amino acid accumulation in Tigriopus californicus (Copepoda). J Comp Physiol B 158:99–105
Guisande C, Maneiro I, Riveiro I (1999) Homeostasis in the essential amino acid composition of the marine copepod Euterpina acutifrons. Limnol Oceanogr 44:691–696
Guisande C, Riveiro I, Maneiro I (2000) Comparisons among the amino acid composition of females, eggs and food to determine the relative importance of food quantity and food quality to copepod reproduction. Mar Ecol Prog Ser 202:135–142
Guillaume J (1997) Protein and amino acids. In: D´Abramo LR, Conklin DE, Akiyama DM (eds) Crustacean nutrition. Advances in world aquaculture, vol 6. World Aquaculture Society, Louisiana State University, Baton Rouge
Hansen PJ, Bjørnsen PK, Hansen BW (1997) Zooplankton grazing and growth: scaling within the 2–2,000-μm body size range. Limnol Oceanogr 42:687–704
Helland S, Triantaphyllidis GV, Fyhn HJ, Evjen MS, Lavens P, Sorgeloos P (2000) Modulation of the free amino acid pool and protein content in populations of the brine shrimp Artemia spp. Mar Biol 137:1005–1016
Helland S, Nejstgaard JC, Humlen R, Fyhn HJ, Båmstedt U (2003a) Effects of season and maternal food on Calanus finmarchicus reproduction, with emphasis on free amino acids. Mar Biol 142:1141–1151
Helland S, Terjesen BF, Berg L (2003b) Free amino acid and protein content in the planktonic copepod Temora longicornis compared to Artemia franciscana. Aquaculture 215:213–228
Hirche H-J (1996a) The reproductive biology of the marine copepod, Calanus finmarchicus—a review. Ophelia 44:111–128
Hirche H-J (1996b) Diapause in the marine copepod, Calanus finmarchicus—a review. Ophelia 44:129–143
Hygum BH, Rey C, Hansen BW, Tande K (2000) Importance of food quality to structural growth rate and neutral lipid reserves accumulated in Calanus finmarchicus. Mar Biol 136:1057–1073
Ikeda A, Kanno Y, Ozaki K, Shinada A (2001) Metabolic rates of epipelagic marine copepods as a function of body mass and temperature. Mar Biol 139:587–596
Jaschnov WA (1972) On the systematic status of Calanus glacialis, Calanus finmarchicus and Calanus helgolandicus. Crustaceana 22:279–284
Jeffries HP (1969) Seasonal composition of temperate plankton communities: free amino acids. Limnol Oceanogr 14:41–52
Jeffries HP, Alzara L (1970) Dominance-diversity relationships of the free amino acids in coastal zooplankton. Comp Biochem Physiol 37:215–223
Kattner, G, Krause M (1987) Changes in lipids during the development of Calanus finmarchicus from copepodid I to adult. Mar Biol 96:511–518
Klein Breteler WCM, Schogt N, Gonzales SR (1990) On the role of food quality in grazing and development of life stages, and genetic change of body size during cultivation of pelagic copepods. J Exp Mar Biol Ecol 135:177–189
Klein Breteler WCM, Schogt N, Baas M, Schouten S, Kraay GW (1999) Trophic upgrading of food quality by protozoans enhancing copepod growth: role of essential lipids. Mar Biol 135:191–198
Kleppel GS (1993) On the diets of calanoid copepods. Mar Ecol Prog Ser 99:183–195
Lee RF (1974) Lipid composition of the copepod Calanus hyperboreus from the Arctic Ocean. Changes with depth and season. Mar Biol 26:313–318
Lee RF, Neventzel JC, Paffenhöfer GA, Benson AA (1970) The metabolism of wax esters and other lipids by the marine copepod Calanus helgolandicus. J Lipid Res 11:237–240
Levinsen H, Turner JT, Nielsen TG, Hansen BW (2000) On the trophic coupling between protists and copepods in arctic marine ecosystems. Mar Ecol Prog Ser 204:65–77
Lowry OH, Rosebrough NJ, Lewis Farr A, Randall RJ (1951) Protein measurement with the folin phenol reagent. J Biol Chem 193:265–275
Marshall SM, Orr AP (1955) The biology of a marine copepod Calanus finmarchicus Gunnerus. Oliver and Boyd, Edinburgh
Mayzaud P (1976) Respiration and nitrogen excretion of zooplankton. IV. The influence of starvation on the metabolism and the biochemical composition of some species. Mar Biol 37:47–58
Menden-Deuer S, Lessard EJ (2000) Carbon to volume relationships for dinoflagellates, diatoms, and other protist plankton. Limnol Oceanogr 45:569–579
Mommsen TP (2001) Paradigms of growth in fish. Comp Biochem Physiol 129:207–219
Nejstgaard JC, Witte HJ, Wal P van der, Jacobsen A (1994) Copepod grazing during a mesocosm study of an Emiliania huxley (Prymnesiophyceae) bloom. Sarsia 79:369–377
Nejstgaard JC, Gismervik I, Solberg PT (1997) Feeding and reproduction by Calanus finmachicus, and microzooplankton grazing during mesocosm blooms of diatoms and the coccolitophore Emiliania huxleyi. Mar Ecol Prog Ser 147:197–217
Nejstgaard J, Naustvoll L-J, Sazhin A (2001a) Correcting for underestimation of microzooplankton grazing in bottle incubation experiments with mesozooplankton. Mar Ecol Prog Ser 221:59–75
Nejstgaard J, Hygym B, Naustvoll L-J, Båmstedt U (2001b) Zooplankton growth, diet and reproductive success compared in simultaneous diatom- and flagellate- microzooplankton-dominated plankton blooms. Mar Ecol Prog Ser 221:77–91
Naess T, Germain-Henry M, Naas KE (1995) First feeding of Atlantic halibut (Hippoglossus hippoglossus) using different combinations of Artemia and wild zooplankton. Aquaculture 130:235–250
Ohman MD, Runge JA (1994) Sustained fecundity when phytoplankton resources are in short supply: omnivory by Calanus finmarchicus in the Gulf of St.Lawrence. Limnol Oceanogr 39:21–36
Østvedt O-J (1955) Zooplankton investigations from Weather Ship M in the Norwegian Sea, 1948–49. Hvalråd Skr 40:1–93
Parsons TR, Maita Y, Lalli CM (1984) A manual of chemical and biological methods for seawater analysis. Pergamon Press, Oxford
Pasternak A, Arashkevich E, Tande K, Falkenhaug T (2001) Seasonal changes in feeding, gonad development and lipid stores in Calanus finmarchicus and C. hyperboreus from Malangen, northern Norway. Mar Biol 138:1141–1152
Putt M, Stoecker DK (1989) An experimentally determined carbon: volume ratio for marine "oligotrichous" ciliates from estuarine and coastal waters. Limnol Oceanogr 34:1097–1103
Rønnestad I, Finn NR, Groot EP, Fyhn HJ (1992) Utilization of free amino acids related to energy metabolism of developing eggs and larvae of lemon sole Microstomus kitt reared in the laboratory. Mar Ecol Prog Ser 88:195–205
Rousseau V, Mathot S, Lancelot C (1990) Calculating carbon biomass of Phaeocystis sp. from microscopic observations. Mar Biol 107:305–314
Sargent JR, Lee RF, Neventzel JC (1976) Marine waxes. In: PE Kolattukdy (ed) Chemistry and biochemistry of natural waxes. Elsevier, Amsterdam, pp 50–91
Schoffeniels E (1976) Adaptations with respect to salinity. Biochem Soc Symp 41:179–204
Svensen C, Tande K (1999) Sex change and female dimorphism in Calanus finmarchicus. Mar Ecol Prog Ser 176:93–102
Svensen C, Egge J, Stiansen J (2001) Can silicate and turbulence regulate the vertical flux of biogenic matter? A mesocosm study. Mar Ecol Prog Ser 217:67–80
Svensen C, Nejstgaard JC, Egge JK, Wassmann P (2002) Pulsing versus constant supply of nutrients (N, P and Si): effect on phytoplankton community, mesozooplankton grazing and vertical flux of biogenic matter. Sci Mar 66:189–203
Tande KS (1991) Calanus in north Norwegian fjords and in the Barents Sea. Polar Res 10:389–407
Torres JJ, Aarset AV, Donnelly J, Hopkins TL, Lancraft TM, Ainley DG (1994) Metabolism of Antarctic micronektonic Crustacea as a function of depth of occurrence and season. Mar Ecol Prog Ser 113:207–219
Tulli F, Tibaldi E (1997) Changes in amino acids and essential fatty acids during early larval rearing of dentex. Aquaculture 5:229–236
Wiborg KF (1955) Investigations on zooplankton in coastal and offshore waters of western and northwestern Norway with special reference to the copepods. Fisk Dir Skr Ser Havunders 11:1–246
Williams P le B, Egge JK (1998) The management and behaviour of the mesocosm. Estuar Coast Shelf Sci 46:3–14
Yancey PH, Clark ME, Hand SC, Bowlus RD, Somero GN (1982) Living with water stress: evolution of osmolyte systems. Science 217:1214–1222
Zar JH (1996) Biostatistical analysis, 3rd edn. Prentice-Hall, London
Acknowledgements
We acknowledge the experimental and field assistance of Rikard Humlen and the analytical assistance of Maria Sula Evjen, Monica Martinussen and Aud Larsen. This work was a part of the MARICULT project NAPP and was supported by grants from the Research Council of Norway. These experiments comply with the current animal experimental laws of Norway, the country in which the present work was conducted.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by L. Hagerman, Helsingør
Rights and permissions
About this article
Cite this article
Helland, S., Nejstgaard, J.C., Fyhn, H.J. et al. Effects of starvation, season, and diet on the free amino acid and protein content of Calanus finmarchicus females. Marine Biology 143, 297–306 (2003). https://doi.org/10.1007/s00227-003-1092-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00227-003-1092-x