
Abstract
Understanding the fundamental characteristics of prosthetic movement control is imperative in improving prosthesis design and training. This study quantified how using an upper limb prosthesis affected performance during goal-directed reaching tasks. Nine prosthesis users with unilateral transradial limb absence and nine healthy controls completed a series of goal-directed reaching movements with different goals: one spatial and three temporal with different goal frequencies. We quantified end-point accuracy, smoothness, and peak speed for the spatial task and temporal accuracy, horizontal distance, and speed for the temporal task. For the temporal task, we also used a goal-equivalent manifold (GEM) approach to decompose variability in movement distance and speed into those perpendicular and tangential to the GEM. Detrended fluctuation analysis (DFA) quantified the temporal persistence of each time series. For the spatial task, movements made with prostheses were less smooth, had larger end-point errors, and had slower peak speed compared to those with control limbs (p < 0.041). For the temporal task, movements made with prostheses and intact limbs of prosthesis users and control limbs were similar in distance and speed and had similar timing errors (p > 0.138). Timing errors, distance, speed, and GEM deviations were corrected similarly between prosthetic limbs and control limbs (p > 0.091). The mean and variability of distance, speed, and perpendicular deviations decreased with increased goal frequency (p < 0.001). Our results suggest that prosthesis users have a sufficient internal model to successfully complete ballistic movements but are unable to accurately complete movements requiring substantial feedback.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Goal-directed movements such as reaching with a spatial goal or a time constraint are fundamental components of many activities of daily living. For individuals with upper limb absence, these activities are often accomplished with a body-powered or myoelectric prosthesis. These prostheses are controlled either mechanically (body-powered) or electrically (myoelectric) to open and close the end effector. While a prosthesis aims to replace the function of an anatomical limb, upper limb prosthesis users expressed low device satisfaction driven by the ‘lack of function’ of their prosthesis (Smail et al. 2020). Prosthesis users also reported difficulty accomplishing simple activities of daily living (Jang et al. 2011). Due to the differences in properties between a prosthesis and an anatomical limb, it is reasonable to assume that prosthetic movements are fundamentally different from anatomical movements. If and how these movements differ from each other remain unclear, however.
Although end-point accuracy is an important component of goal-directed reaching, only a few studies have quantified movement accuracy in upper limb prosthesis users (Doeringer and Hogan 1995; Schabowsky et al. 2008; Metzger et al. 2010). In these studies, upper limb prosthesis users completed reaching movements toward spatial targets displayed on a screen before or continuously during the reach. Participants were able to successfully reach targets both with (Metzger et al. 2010) and without visual feedback during the task (Doeringer and Hogan 1995; Metzger et al. 2010), suggesting they have some degree of proprioceptive feedback (Doeringer and Hogan 1995) and an appropriate internal model of the prosthesis (Metzger et al. 2010). However, when transhumeral prosthesis users were given spatial targets with a speed constraint (i.e., dynamic tracking), they were no longer able to match the performance of their intact limb (Doeringer and Hogan 1995). While previous studies allude to the possibility of differences in the way prosthetic and anatomical movements are controlled, the discrepancy between studies suggests that these differences are likely task dependent.
In addition to end-point accuracy, it is important to understand how individuals achieve a given task. Anatomical limbs exhibit equifinality, meaning that an infinite number of possible motor solutions are available to successfully complete a task (Bernstein 1967; Cusumano and Cesari 2006; Latash et al. 2007). For goal-directed arm movements, equifinality of the anatomical limbs allows movement flexibility, from which different movement strategies can be used to achieve the same goal. For example, individuals can use multiple shoulder, elbow, and wrist positions to achieve the same goal position or multiple end-point speed and distance combinations can achieve the same goal time. Healthy individuals without limb absence can exploit motor redundancy to maintain task accuracy in the presence of muscle fatigue (Gates and Dingwell 2008; Singh et al. 2010; Ranganathan et al. 2013; Cowley et al. 2014) and pain (Chehrehrazi et al. 2017; Cowley and Gates 2018). It is unclear whether prosthesis users can take advantage of this redundancy to the same extent healthy individuals without limb loss can. Prior work suggests that prosthesis users employ alternate movement strategies compared to individuals without limb absence to complete a task with the same accuracy. For example, similarly accurate movements with prostheses were completed with lower peak speed (Doeringer and Hogan 1995; Metzger et al. 2010) and greater number of sub-movements (Doeringer and Hogan 1995) compared to anatomical movements. This is significant as a greater number of sub-movements is associated with a less smooth movement and higher metabolic costs (Nishii and Taniai 2009).
Following limb amputation, individuals typically experience cortical reorganization, or the invasion of the adjacent areas of the somatosensory map onto the hand territory (Gunduz et al. 2020). While the extent of this reorganization may not always be complete (Makin et al. 2013, 2015), amputation and changes in somatotopic reorganization may impact movement planning and control, altering the way individuals exploit motor redundancy and influencing their movement strategy. One study demonstrated that prosthesis users were able to achieve similarly smooth and straight trajectories as healthy individuals when completing a ballistic reaching task (Schabowsky et al. 2008). However, when presented with a velocity-dependent force field during reaching, prosthesis users exhibited greater movement trajectory error and variability than healthy individuals during the late phase of adaptation to the force field (Schabowsky et al. 2008). Therefore, we may speculate that prosthesis users have difficulty incorporating novel cues to update their internal model, possibly due to post-amputation neural plasticity. In particular, previous research discovered atypical bilateral activation of posterior parietal regions during unilateral raising and rotating task using prosthetic limbs, instead of predictable activation of contralateral motor areas (Williams et al. 2016). Since another study reported an association between larger learning errors and the disruption of posterior parietal cortex in healthy individuals (Della-Maggiore et al. 2004), it is possible that the cortical reorganization following amputation impacts movement planning in prosthesis users. This theory is further supported by the fact that while prosthesis users were able to perform with similar accuracy as healthy individuals in a reaching task without visual feedback, they employed decreased movement speed to maintain their accuracy (Metzger et al. 2010). The compensation of speed with accuracy demonstrates an uncertainty of the internal model of the prosthetic limb, which is essential in movement planning. While it is clear that neural plasticity following amputation has implications in prosthetic movement control, how this impacts strategy has not been sufficiently explored.
The goal of this paper was to quantify movement accuracy and strategies during repetitive goal-directed reaching movements with spatial or temporal goals. The goal of the spatial task was to reach toward a specific location without regard to how long it took to complete the movement. In contrast, the goal of the temporal task was to time the reach with a metronome set to one of three frequencies, without regard to movement location. Based on prior literature on spatial reaching, we expected that prosthetic and anatomical movements would exhibit similar spatial accuracy, but would employ different movement strategies. For the temporal task, we employed a Goal-Equivalent Manifold (GEM) (Cusumano and Cesari 2006) approach that maps body state variables (e.g., distance and speed) to a defined goal variable (e.g., time). Each body state variable combination that maintains the defined goal is a possible motor solution to the task. This approach helps elucidate the strategy employed by individuals (i.e., which body state variable combinations are used to complete the task?) and the variability associated with the strategy (i.e., how wide is the range of employed motor solutions?). Similar to our hypothesis regarding spatial reaching, we hypothesized that anatomical and prosthetic limbs would exhibit similar timing accuracy but different movement strategies. We also hypothesized that the differences in strategies would be larger as the task became more difficult (i.e., faster goal frequency).
Methods
Participants
Nine prosthesis users and nine age- and sex-matched healthy controls without limb absence participated in the study (Table 1). Prosthesis users were included if they had a unilateral transradial limb absence, were over the age of 18, and used an upper limb prosthesis for at least four months. Potential participants from both groups were excluded if they had a history of neurological disorders or visual, balance, or hearing impairments that would limit their ability to perform the study protocol. The study protocol was approved by the University of Michigan Medical School IRB and all participants provided their written informed consent prior to participation. All control participants were right hand dominant, as determined by a modified version of the Edinburgh Inventory (Oldfield 1971). Participants with limb absence used a body-powered prosthesis (n = 3), a myoelectric prosthesis (n = 3), or both (n = 3).
Experimental protocol
All participants performed two sets of constrained, planar goal-directed reaching movements using a custom-built planar reaching device. The device consisted of a two-link robotic arm, based on the design described in (Casadio et al. 2006), and an adjustable, straight-backed chair. In the first set of movements, participants reached toward spatial targets without concern for timing (‘spatial task’), while they attempted to match timing without concern for position in the second set (‘temporal task’). Prosthesis users did not actuate their terminal device during the task. This study design was chosen to understand how the internal model developed during prosthesis users’ daily lives impacts movement accuracy and strategy during reaching with differing explicit goals. For each task, participants first performed a minimum of 20 practice trials or practiced until they felt comfortable with the task. The two-dimensional positions of the handle were collected using integrated encoders in two motors (Emerson Industrial Automation, St. Louis, MO) that were used as inputs for the custom software (MATLAB, MathWorks, MA). Sampling frequency was dependent on the runtime of the custom software and varied between 50 and 80 Hz.
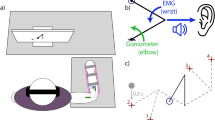
During the spatial task, participants were instructed to match the handle position to a target location shown on a computer screen. The radius of the on-screen target was 10 mm, while the radius of the on-screen handle representation was 9 mm. For each trial, the participants received two seconds to reach towards the target with the handle and hold before the target moved to a new location on the computer screen (Fig. 1). While the explicit goal of the spatial task was defined by the spatial accuracy of the handle, a time constraint was given to prompt the next target. This time was chosen based on the average completion time during pilot testing. The location of the target oscillated between two locations that were 20 cm apart horizontally.

Participants performed goal-directed repetitive reaches using a two-link robotic arm whose position was displayed on a computer screen. a In the spatial task, participants reached towards a circular target with the goal of matching the center of the target with the handle as closely as possible. b In the temporal task, participants were instructed to match a change in horizontal direction with the time of a metronome
During the temporal task, the participants were instructed to move the handle horizontally across the computer screen between two large shaded regions that were 20-cm in width and 20-cm apart in time with a metronome (Fig. 1). While the explicit goal of the temporal task was defined by movement timing only, we asked the participants to make excursions between two wide spatial regions to discourage participants from making very small movements or not moving at all. The participants completed 300 goal-directed reaches with each limb at each of three frequencies: 1, 1.5, and 2 Hz. These goal frequencies were chosen since the average natural frequency of upper extremity movements of healthy individuals is approximately 1 Hz (Gates and Dingwell 2008). The order of testing was consistent across participants as intact (dominant) limb followed by the prosthetic (non-dominant) limb at 1.5 Hz, 1 Hz, and 2 Hz.
Data analysis
The handle and target positions were interpolated to 100 Hz using a ‘cubic spline’ and ‘previous’ interpolant, respectively, in Matlab (Mathworks, Natick, MA, USA) to have uniform sampling frequency across all trials. Handle data were then filtered using a 5th order low-pass Butterworth filter with a cutoff frequency of 15 Hz.
For each movement of the spatial task, we quantified end-point errors, peak speed, and number of sub-movements during the active reaching phase. End-point error was the average distance between the centers of the target and handle during the last 10% (200-ms) of each movement. Peak speed was defined by the maximum movement speed of each trial. The active reaching phase was defined by any duration of the trial where movement speed was greater than 5 cm/s. We quantified the number of zero-crossing of acceleration during the reaching phase as sub-movements. We visually inspected the speed profile of each movement and excluded the analysis of spatial error on trials where participants started reaching to the next target before it was displayed (2.2% of total trials). For the majority of the participants, the speed profile of the first movement visually differed from all other movements and was, therefore, excluded from analysis.
For the temporal task, we quantified the timing error as the time difference between each peak excursion to its nearest metronome signal (Fig. 1). Negative timing errors indicated that the person reached the target before the metronome (i.e., anticipatory), while positive errors indicated that the person reached the target after the metronome (i.e., responsive). We non-dimensionalized horizontal distance and speed of each reach (\(D\) and \(S\), respectively) by rescaling the distance traveled by \(\frac{1}{H}\) where \(H\) is the height of each participant and the speed by \(\frac{1}{H*f}\) where \(f\) is the frequency of the metronome. The non-dimensional distance, \(D\), was equal to the non-dimensional speed, \(S\), at every location where the goal, \(f\), was reached (Gates and Dingwell 2008). The goal of the temporal task was to match the movements with the timing of the metronome signal. All [\(D\), \(S\)] combinations where \(D\)/\(S\) =\(T\) achieve the task goal (\(T\); movement time) and define the GEM for the task. Variation in \(D\) and \(S\) were decomposed into deviations perpendicular (\({\delta }_{P}\); goal-relevant) and tangential to the GEM (\({\delta }_{T})\). The relative variability of \({\delta }_{P}\) and \({\delta }_{T}\) indicates whether the movement strategy employed affected body parameters (\(D\), \(S\)), goal parameters (\(T\)), or both.
We quantified how individuals controlled their movements using detrended fluctuation analysis (DFA). This method quantifies the temporal persistence in a time series (Peng et al. 1993, 1995). Using the approach described in (Gates and Dingwell 2008), we calculated a scaling exponent,\(\alpha\), which indicates what type of correlations are present in a time series. When the \(\alpha\) < 0.5, the time series exhibits “anti-persistent” correlations, which is an actively controlled process with deviations in one direction likely to be followed by deviations in the opposite direction. When \(\alpha\) = 0.5, the time series is considered to be completely uncorrelated and therefore exhibit characteristics of white noise. When \(0.5<\alpha \le 1.0\), the time series exhibits “persistent” correlations where deviations in one direction are likely to be followed by deviations in the same direction (i.e., not immediately corrected). We calculated an \(\alpha\) value for each time series for the temporal task (timing errors, \(S, D\), \({\delta }_{P}\), \({\delta }_{T}\)) only, as the spatial error time series was non-continuous due to excluded trials. The quantification and comparison of \(\alpha\) value between different groups can help elucidate the level of control exerted over each specific outcome.
Statistical analysis
The primary dependent measures for the spatial task were the mean magnitude and variability (standard deviation) of end-point errors and the mean and variability of the number of sub-movements and peak speed. The primary dependent measures for the temporal task were the mean, variability (standard deviation), and \(\alpha\) of \({E}_{t}\), \(D\),\(S\), \({\delta }_{P},\) and \({\delta }_{T}\). We first tested for differences between right and left limbs of controls using a paired t-test (2-tailed, significance level of 0.05). There were no significant differences for any comparison except mean and variability of \({\delta }_{P}\). In all cases where this difference was non-significant, we combined the data from right and left limbs of control participants into a ‘control’ limb for subsequent analyses.
For each dependent measure of the spatial task, we tested for differences between limb type (control, intact, prosthesis) using a linear mixed model where limb type was a fixed factor and participants was a random factor. For each dependent measure of the temporal task, we tested for difference between limbs and frequencies using a linear mixed model where limb type (control, intact, prosthetic), goal frequency (1, 1.5, 2 Hz), and interaction of limb type and goal frequency were fixed factors and participants was a random factor. Significant interactions were explored using post-hoc pairwise comparison of estimated marginal means with a Sidak correction. All statistical analyses were performed using SPSS (version 24, IBM Corp, Armonk, NY) with α = 0.05. Effect sizes of significant pairwise comparisons of limb type were calculated (Hedge’s g) and were considered small (g \(\ge\) 0.2), medium (g \(\ge\) 0.5), or large (g \(\ge\) 0.8) (Cohen 2013).
Results
One prosthesis user who used both body-powered and myoelectric prostheses, S08, did not return to complete the temporal task with his myoelectric device. Due to technical difficulties, S01’s intact limb data for the 2 Hz goal frequency was excluded from the analysis.
Result of the spatial task
There was a significant main effect of limb on mean and variability of end-point errors (p \(\le\) 0.005), the mean number of sub-movements (p = 0.001), and mean peak speed (p = 0.036) (Figs. 2 and 3). Prosthetic movements were completed with larger mean and variability of end-point errors (p < 0.005, g \(\ge\) 1.41), greater number of sub-movements (p = 0.001, g = 1.71), and slower peak speed (p = 0.041, g = 1.14) compared to movements completed by control limbs. Movements completed by the intact limb of prosthesis users also had greater end-point error variability (p = 0.019, g = 1.49) and a greater number of sub-movements (p = 0.037, g = 1.44).

Excursion trajectory of the a spatial task completed with prosthetic (blue) and intact limb (green) and the b temporal task completed with a prosthetic limb with a goal frequency of 1 Hz (red), 1.5 Hz (light blue), and 2 Hz (purple) completed by a representative participant. Spatial task goal is illustrated as a circle and the boundaries of the grey shaded region of the temporal task are illustrated on either side of the computer screen

Mean and variability of end-point errors, number of sub-movements, and peak speed between control limbs (grey) and intact (green) and prosthetic limbs (blue) of prosthesis users with standard error bars. Individual points represent individual limbs. For prosthetic limbs, blue and red points represent limbs of individuals with acquired and congenital limb absence, respectively. Statistically significant pairwise comparisons are denoted by * (p < 0.05)
Results of the timing task
There was a significant main effect of goal frequency on mean and variability of distance (\(D\)) and speed (\(S\)) (p < 0.001; Figs. 4 and 5). The mean and variability of \(D\) and \(S\) generally decreased with increasing goal frequency, though this was not significant for all pairwise comparisons (Appendix 1). There were no other significant effects of limb (p \(\ge\) 0.116) nor any interactions effects (p \(\ge\) 0.765) for the mean or variability \(D\) and S. While there was a significant main effect of limb on \(\alpha\) of \(S\) (p = 0.035), none of the post-hoc pairwise comparisons were significant (p \(\ge\) 0.071). There were no other significant main effects or interaction effects \(\alpha\) of \(D\) and S (p \(\ge\) 0.114).

Mean, variability, and \(\alpha\) of a non-dimensional distance (\(D\)), non-dimensional speed (\(S\)), and b timing errors between control limbs (grey) and intact (green) and prosthetic limbs (blue) of prosthesis users across three different goal frequencies—1, 1.5, and 2 Hz with standard error bars. Individual points represent individual limbs. For prosthetic limbs, blue and red points represent limbs of individuals with acquired ad congenital limb absence, respectively. Statistically significant pairwise comparisons of limb and frequency effect are denoted by * and ‡, respectively (p < 0.05). Outcomes with all significant pairwise comparisons are noted on the y-axis label

Normalized horizontal reach distance (\(D\)) and speed (\(S\)) of each excursion are shown for each goal frequency of a single prosthesis user along the GEM (dashed line) for prosthetic (blue) and intact (intact) limbs and the right limb of their age- and sex-matched control. Mean operating points of prosthetic (blue circle), intact (green diamond), and control (grey square) limbs across all participants for each goal frequency were illustrated. Variability of tangential (\({\delta }_{T}\)) and perpendicular deviation (\({\delta }_{P}\)) of the GEM for each limb are illustrated as ellipses with their respective color
There was a significant main effect of frequency on the mean timing errors (p < 0.001). Error switched from positive values (i.e., responsive) for lower goal frequencies (1 Hz, 1.5 Hz) to negative values (i.e., anticipatory) for the higher goal frequency (2 Hz) (Fig. 5). Main effect of limb (p = 0.138) and interaction effect (p = 0.758) were not significant. There were no effects of limb or frequencies on \(\alpha\) of timing errors (p \(\ge\) 0.054).
Goal-equivalent manifold (GEM) results
Participants aligned their movements with the GEM such that deviations perpendicular (\({\delta }_{P})\) to the GEM were much smaller than those tangential (\({\delta }_{T})\) to the GEM (p < 0.001; Figs. 5 and 6). There were significant main effects of goal frequency (p < 0.001) on the variability of\({\delta }_{P}\), but no limb type (p = 0.052) or frequency by limb type interaction (p = 0.518). Variability of \({\delta }_{P}\) decreased (p < 0.001) with increased goal frequency while \(\alpha\) of \({\delta }_{P}\) (p = 0.006) increased. There were no other significant main effects nor interaction effects on the \(\alpha\) of \({\delta }_{T}\) or \({\delta }_{P}\) (p \(\ge\) 0.114).

Mean, variability, and \(\alpha\) of tangential deviation (\({\delta }_{T}\)) and perpendicular deviation (\({\delta }_{P}\)) of the GEM between control limbs (grey) and intact (green) and prosthetic limbs (blue) of prosthesis users across different goal frequencies with standard error bars. Individual points represent individual limbs. For prosthetic limbs, blue and red points represent limbs of individuals with acquired and congenital limb absence, respectively. Statistically significant pairwise comparisons of limb and frequency effect are denoted by * and ‡, respectively (p < 0.05). Outcomes with all significant pairwise comparisons are noted on the y-axis label
Discussion
Upper limb prosthesis users self-report difficulty completing activities of daily living. To date, little is known about how individuals with limb absence complete goal-directed reaching, which is a fundamental component of many activities of daily living. In this study, we characterized movement accuracy, strategy, and control of repetitive reaches toward spatial and temporal targets. During the spatial task, prosthetic movements were completed with lower smoothness, peak speed, and end-point accuracy. There were no differences in movement accuracy, strategy, or control between limbs for the temporal task, however. Overall, the results suggest that individuals are capable of achieving similar outcomes with their prosthetic limb to their anatomical limb, depending on the task goal.
The temporal task was designed to allow movement variability and investigate whether movement strategy differed between prosthetic and anatomical movements. As such, instead of having a target that required many fine adjustments, multiple solutions to the task existed as long as movements had consistent timing signaled via a metronome. Therefore, the task predominantly required familiarity with internal model and feedforward control. While there are several theories surrounding human motor control, one popular principle pertains to feedforward control, or the ability to plan movements through an internal model. Given the delay in incorporating feedback cues from the environment, feedforward control is essential in executing fast and coordinated movements, such as the temporal task that involves ballistic movements. The successful development of an internal model depends on an individual’s familiarity with the mechanical property of the body that they are controlling (i.e., prosthesis and anatomical limbs), which can be established over time and spontaneously (Kim 2019). Our results from the temporal task suggest that prosthesis users were able to develop an adequate internal model. Similarly, another study interpreted their findings that prosthesis users completed movements with accuracy similar to anatomical movements as indicative of an appropriately updated internal model (Metzger et al. 2010). In our study, prosthetic and anatomical movement were altered as goal frequency increased by switching from ‘responding’ to ‘anticipating’ the signal but maintaining a consistent goal-relevant deviation. To maintain this performance, participants made faster corrections of goal-relevant deviation as the goal frequency increased (i.e., α decreased). Not only did the participants switch the way they responded to the metronome as goal frequency increased, the mean timing error also decreased with frequency. Since the timing error was calculated as the time distance between each peak excursion to its nearest metronome signal, the maximum obtainable timing error for a given frequency was half of the period. Moreover, decreases in the magnitude of mean timing error with increasing goal frequency may be indicative of a less consistent response type (i.e., alternating between responding and anticipating) within limb types. Therefore, complementing comparison of mean accuracy with the GEM analysis that dissects employed movement strategy and its relation to overall performance can lead to a better understanding of how prosthetic movements fundamentally differ from anatomical movements.
Movements across all limb types in both the prosthesis user and control participants had similar mean distance, speed, and goal-irrelevant deviation during the temporal task (Fig. 4). Contrary to our hypothesis that a faster goal frequency would result in greater differences between limb types, using a prosthesis did not impact the way individuals adapted to a more challenging task. The movement patterns of different limbs also exhibited a similar curved trajectory (Fig. 5). This movement pattern is likely the path of least resistance in the two-link robotic arm, rather than a fundamental characteristic of this type of movement. However, it is noteworthy that all participants, regardless of limb type, utilized this path. As the goal frequency increased, the participants made shorter and slower reaches with decreased variability in their distance and speed. Our results suggest that both prosthesis users and control participants were able to respond to changes in goal frequency by changing their movement strategies in a way that preserved the overall performance. Therefore, we may speculate that participants were able to use sufficiently developed internal models to complete the task, regardless of their limb type.
While prosthesis use did not impact movement accuracy and strategy of ballistic movements such as in the temporal task, it did impact movement quality and accuracy during the spatial task. The spatial task required individuals to incorporate both movement planning, or feedforward control, and movement adjustments, or feedback control (Metzger et al. 2010). While individuals without limb absence can incorporate both visual and proprioceptive feedback during the task, prosthesis users likely have to rely heavily on visual feedback. Visual feedback is slower to incorporate into motor control compared to other feedback modalities (Bongers et al. 2012) and the synthesis of proprioception and vision is integral in movement control (Graziano 1999; Sober and Sabes 2005, Kuchenbecker et al. 2007). Moreover, the force resistance from the prosthetic socket during movements involved in the task may have introduced additional challenges (Metzger et al. 2010).
The aforementioned challenges associated with prosthetic movement control suggest that prosthetic movements would yield lower end-point accuracy compared to anatomical movements. However, prior work reported similar end-point accuracy between prosthetic and anatomical movements (Schabowsky et al. 2008; Metzger et al. 2010). In one of the previous studies (Metzger et al. 2010), prosthetic movements had a lower peak velocity than anatomical limbs, suggesting that prosthesis users were able to achieve the same level of accuracy by reducing their movement speed. Similarly, in our study, prosthetic movements were completed with lower peak speed compared to anatomical movements of control participants. Contrary to previous work, prosthetic movements in this study were also completed with lower end-point accuracy despite implementing a strategy that involved lower peak speed. This may be explained by the fact that our task imposed a reach duration limit of two seconds. This duration was chosen based on pilot testing of healthy individuals and a few individuals with upper limb absence, and may not have been sufficient for all participants to complete the task with high accuracy. In other words, it is possible that participants would have accomplished the task with similar end-point accuracy regardless of the limb type if they were given more time, although it is unclear whether longer reach duration limit would have yielded even slower peak speed in prosthetic movements. Our results indicate that prostheses users are limited in their ability to achieve fast and accurate movements.
Prosthetic movements also had lower movement smoothness compared to anatomical movements during the spatial task, which is consistent with previous reports of lower movement quality in prosthesis users during reach and grasp movements (Bouwsema et al. 2010) and activities of daily living (Cowley et al. 2017; Engdahl and Gates 2021). While all prior work found that there is a lack of smoothness in grasping, results from this study suggest that prosthetic movements exhibit lower movement smoothness even in tasks that require no activation of the end-effector. Collectively, the lower smoothness in prosthetic reach and grasp movements indicate that prosthesis users make metabolically inefficient movements.
It is interesting to note that anatomical movements completed by the contralateral intact limb of prosthesis users exhibited greater end-point error variability and slower peak speed compared to anatomical movements completed by control participants. This suggests that changes in movement control from prosthesis use also impact how movements are completed by the contralateral anatomical limb. Our findings are consistent with results from another study that observed abnormal movement trajectory of the intact arm in prosthesis users during a similar reaching task (Metzger et al. 2010). Metzger et al. partially attributed their findings to the order of testing where the intact limb was tested after the prosthetic limb. Given that the prosthetic and intact limbs have different mechanical properties, intact limb performance would have been impacted if individuals used an internal model established for their prosthesis to complete the task with their intact limb. While the protocol of the spatial task similarly involved testing the two limb types in sequence, the order was reversed such that the intact limb was always tested first. Therefore, the likely explanation for this phenomenon is that prosthesis use changed the overall movement control of the body such that intact limb movements were also affected. Previous studies have noted that during bilateral tasks, unilateral prosthesis users complete movements such that their intact limb movement patterns match that of their prosthetic limb (Engdahl and Gates 2021). Our results further suggest that the difference in movement control and the resulting movement pattern seen in contralateral intact limbs of prosthesis users may also persist in unilateral tasks not involving prosthesis.
There are several limitations to be considered while interpreting results from this study. First, prosthesis users who participated in this study were relatively heterogeneous, with a mix of individuals who had congenital or acquired limb absence and with varying levels of prosthesis experience and proficiency. In particular, the prosthetic type varied among prosthesis users, some using a body-powered device with a harnessing system over their shoulders and others using a myoelectric device with a socket suspension over their epicondyles. While suspension mechanism can influence the interaction between the residual limb and their prosthesis, the prosthesis users completed the reaching tasks with their at-home devices with which they were able to develop the internal model. Nevertheless, it is possible that the varying duration of prosthetic ownership and daily use time across individuals influenced their movement strategy. It is also plausible that the difference in typical suspension mechanisms between body-powered and myoelectric prosthesis inherently resulted in different movement strategies among prosthesis users. However, a relatively small sample size (n = 9) limits our ability to generalize our findings to prosthetic movements more broadly. Moreover, the discontinuity of time series from excluding trials during the spatial task did not allow us to quantify the level of persistence of errors in the spatial task. Evaluating the level of persistence in time series across different types of tasks would be useful in understanding movement control of prosthesis users with different task constraints. Tasks chosen in this study were completed without consideration of any wrist mobility, since the majority of prosthesis users today do not use a device with an active wrist joint. However, it is important to acknowledge that the addition of wrist movement in goal-directed reaching increases the degrees of freedom with which these movements can be completed, which may have implications on the available motor solutions. Future work may evaluate how the addition of an active wrist joint impacts movement strategy and control in prosthesis users.
In conclusion, prosthesis users completed the spatial task with reduced movement accuracy, peak speed, and smoothness, but completed the temporal task with similar accuracy and movement strategy compared to individuals without limb absence. Prosthesis users exerted similar movement control to perform comparably to individuals without limb absence during the temporal task. Results from our study indicate that prosthesis users are able to use their feedforward control to preserve performance during a repetitive reaching task, even with increased task difficulty. However, the reduced accuracy and altered movement strategy seen in the spatial task suggest that exclusive reliance on visual feedback may make tasks requiring small corrections or precise movement near a target more difficult with a prosthesis. Our study also highlighted several fundamental differences that exist between prosthetic and anatomical movements. Specifically, we demonstrated that prosthesis users may be able to complete ballistic tasks with a sufficient internal model, but completing tasks that require corrections by incorporating visual and proprioceptive feedback remains a challenge.
Data availability
The data that support the findings of this study are available from the corresponding author, DG, upon reasonable request.
References
Bernstein N (1967) The coordination and regulation of movement. Pergamon, New York
Bongers RM, Kyberd PJ, Bouwsema H, Kenney L, Plettenburg D, Van der Sluis CK (2012) Bernstein’s level of construction of movements applied to upper limb prosthetics. J Prosthet Orthot 24(2):67–76
Bouwsema H, van der Sluis CK, Bongers RM (2010) Movement characteristics of upper extremity prostheses during basic goal-directed tasks. Clin Biomech 25(6):523–529
Casadio M, Sanguineti V, Morasso PG, Arrichiello V (2006) Braccio di Ferro: a new haptic workstation for neuromotor rehabilitation. Technol Health Care 14(3):123–142
Chehrehrazi M, Sanjari MA, Mokhtarinia HR, Jamshidi AA, Maroufi N, Parnianpour M (2017) Goal equivalent manifold analysis of task performance in non-specific LBP and healthy subjects during repetitive trunk movement: Effect of load, velocity, symmetry. Hum Mov Sci 51:72–81
Cohen J (2013) Statistical power analysis for the behavioral sciences. Academic, New York
Cowley JC, Gates DH (2018) Influence of remote pain on movement control and muscle endurance during repetitive movements. Exp Brain Res 236(8):2309–2319
Cowley JC, Dingwell JB, Gates DH (2014) Effects of local and widespread muscle fatigue on movement timing. Exp Brain Res 232(12):3939–3948
Cowley J, Resnik L, Wilken J, Smurr Walters L, Gates D (2017) Movement quality of conventional prostheses and the DEKA Arm during everyday tasks. Prosthet Orthot Int 41(1):33–40
Cusumano JP, Cesari P (2006) Body-goal variability mapping in an aiming task. Biol Cybern 94:367–379
Della-Maggiore V, Malfait N, Ostry DJ, Paus T (2004) Stimulation of the posterior parietal cortex interferes with arm trajectory adjustments during the learning of new dynamics. J Neurosci 24(44):9971–9976
Doeringer JA, Hogan N (1995) Performance of above elbow body-powered prosthesis in visually guided unconstrained motion tasks. IEEE Trans Biomed Eng 42:621–631
Engdahl SM, Gates DH (2021) Differences in quality of movements made with body-powered and myoelectric prostheses during activities of daily living. Clin Biomech 84:105311
Gates DH, Dingwell JB (2008) The effects of neuromuscular fatigue on task performance during repetitive goal-directed movements. Exp Brain Res 187(4):573–585
Graziano MSA (1999) Where is my arm? The relative role of vision and proprioception in the neural representation of limb position. Proc Natl Acad Sci USA 96:10418–10421
Gunduz ME, Pinto CB, Saleh Velez FG, Duarte D, Pacheco-Barrios K, Lopes F, Fregni F (2020) Motor cortex reorganization in limb amputation: a systematic review of TMS motor mapping studies. Front Neurosci 14:314
Jang CH, Yang HS, Yang HE, Lee SY, Kwon JW, Yun BD, Choi JY, Kim SN, Jeong HW (2011) A survey on activities of daily living and occupations of upper extremity amputees. Ann Rehabil Med 35(6):907–921
Kim D (2019) A computational scheme for internal models not requiring precise system parameters. PLoS ONE 14(2):e0210616
Kuchenbecker K, Gurari N, Okamura A (2007) Effects of visual and proprioceptive motion feedback on human control of targeted movement. In: IEEE 10th international conference on rehabilitation robotics. Noordwijk, The Netherlands, pp 513–524
Latash ML, Scholz JP, Schoner G (2007) Toward a new theory of motor synergies. Mot Control 11(3):276–308
Makin TR, Scholz J, Filippini N, Henderson Slater D, Tracey I, Johansen-Berg H (2013) Phantom pain is associated with preserved structure and function in the former hand area. Nat Commun 4:1570
Makin TR, Scholz J, Henderson Slater D, Johansen-Berg H, Tracey I (2015) Reassessing cortical reorganization in the primary sensorimotor cortex following arm amputation. Brain 138(Pt 8):2140–2146
Metzger AJ, Dromerick AW, Schabowsky CN, Holley RJ, Monroe B, Lum PS (2010) Feedforward control strategies of subjects with transradial amputation in planar reaching. J Rehabil Res Dev 47(3):201–211
Nishii J, Taniai Y (2009) Evaluation of trajectory planning models for arm-reaching movements based on energy cost. Neural Comput 21:2634–2647
Oldfield RC (1971) The assessment annd aalysis of handedness: the Edinburgh inventory. Neuropsychologia 9:97–113
Peng CK, Mietus J, Hausdorff JM, Havlin S, Stanley HE, Goldberger AL (1993) Long-range anticorrelations and non-Gaussian behavior of the heartbeat. Phys Rev Lett 70(9):1343–1346
Peng CK, Havlin S, Stanley HE, Goldberger AL (1995) Quantification of scaling exponents and crossover phenomena in nonstationary heartbeat time series. Chaos 5(1):82–87
Ranganathan R, Adewuyi A, Mussa-Ivaldi FA (2013) Learning to be lazy: exploiting redundancy in a novel task to minimize movement-related effort. J Neurosci 33(7):2754–2760
Schabowsky CN, Dromerick AW, Holley RJ, Monroe B, Lum PS (2008) Trans-radial upper extremity amputees are capable of adapting to a novel dynamic environment. Exp Brain Res 188(4):589–601
Singh T, Varadhan SK, Zatsiorsky VM, Latash ML (2010) Fatigue and motor redundancy: adaptive increase in finger force variance in multi-finger tasks. J Neurophysiol 103(6):2990–3000
Smail LC, Neal C, Wilkins C, Packham TL (2020) Comfort and function remain key factors in upper limb prosthetic abandonment: findings of a scoping review. Disabil Rehabil Assist Technol 6:821–830
Sober SJ, Sabes PN (2005) Flexible strategies for sensory integration during motor planning. Nat Neurosci 8(4):490–497
Williams L, Pirouz N, Mizelle JC, Cusack W, Kistenberg R, Wheaton LA (2016) Remodeling of cortical activity for motor control following upper limb loss. Clin Neurophysiol 127(9):3128–3134
Acknowledgements
The authors would like to thank Kelsey Ebbs for assisting with the data collection. Partial support for this research was provided by the Congressionally Directed Medical Research Programs [Award Number W81XWH-16-1-0648].
Funding
Congressionally Directed Medical Research Programs (Grant no. W81XWH-16-1-0648).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors report no conflicts of interest with this work.
Additional information
Communicated by Bill J Yates.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Lee, C., Gonzalez, M.A., Kang, J. et al. Temporal and spatial goal-directed reaching in upper limb prosthesis users. Exp Brain Res 240, 3011–3021 (2022). https://doi.org/10.1007/s00221-022-06476-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-022-06476-7