
Abstract
The effect of body posture on the human soleus H-reflex via electrical stimulation of the tibial nerve at the popliteal fossa was studied. All parameters that may influence the reflex were controlled stringently. H-reflexes were elicited in three different body postures while keeping the level of background muscle activation to a minimum. The H-reflex curve relative to the M-wave curve did not change significantly in any of the body postures. However, the maximal H-reflex amplitude significantly increased in the prone position compared with the sitting (p = 0.02) and standing positions (p = 0.01). The background level of electrical activity of the soleus muscle did not significantly change during varying body postures. Together, these findings indicate that the effectiveness of the spindle primary afferent synapse on the soleus motor neuron pool changes significantly in prone position as compared to sitting and standing positions. Given that we have controlled the confounding factors excluding the head position relative to the gravity and the receptors that may be differentially activated at varying body postures such as the proprioceptors, it is concluded that the tonic activity from these receptors may presynaptically interfere with the effectiveness of the spindle primary afferent synapses on the soleus motor neurons.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
H-reflex has been used as a tool to study the effectiveness of the spindle primary afferent (Ia fibers) synapse on motoneurons under many circumstances (reviewed in Pierrot-Deseilligny and Mazevet 2000; Knikou 2008; Chen and Zhou 2011) including varying body postures, static and dynamic conditions (Schieppati 1987; Shimba et al. 2010). The results on the effect of body posture on the H-reflex varied considerably even though similar H-reflex protocols were used in these studies.
While some experiments showed an increase in the H-reflex, others indicated either a decrease or no change when the position of the body changed from laying position to sitting or standing positions. For example, Knikou and Rymer (2003) studied the H-reflex on a tilt table and found that the reflex increased by 140–180% in any of the positions compared with the supine position. Similar results were reported by others (Aiello et al. 1983).
Using similar methodologies, however, several reports claimed that the H-reflex decreased during standing compared with the laying position (Hayashi et al. 1992; Koceja et al. 1993). More interestingly, Shimba et al. (2010) found that the H-reflex size reduced in the standing position compared with the laying position while the opposite was the case for the stretch reflex. This variance is reasonable since the presynaptic effect is different between the stretch and H-reflexes (Morita et al. 1998). It is worth noting that the H-reflex amplitude can also be limited by the activation of the tendon organ afferents (Ib fibers) during mixed nerve stimulation (Pierrot-Deseilligny and Burke 2012).
There are other studies where it has been claimed that the static body positions do not alter the H-reflex response significantly (Trimble 1998). Therefore, we have three different results regarding the effect of the static body position on the H-reflex.
Even though the above studies used tibial nerve stimulation and recorded the H-reflex from the leg muscles, it is possible that such dramatically varying findings can be due to the following reasons: (1) not controlling the level of background muscle activity preceding the stimulus; (2) not normalizing the level of other receptor activity such as the receptors in the soles of the feet during standing or sitting; and (3) not normalizing the reflex findings to the stimulus intensity.
The aim of this study therefore was to examine the changes in the H-reflex in varying body postures using a standardized approach, which attempts to minimize the above mentioned possible confounding variables. The principle hypothesis was that the effect of the body posture on the H-reflex could be demonstrated only when all parameters that may influence the reflex are controlled stringently. In this approach, the only uncontrolled variable was the changes in body posture.
Methods
A total of 14 informed volunteer men aged 25.4 ± 3.4 participated in the study. The subjects were all healthy with no medical condition affecting the sensorimotor system. The participants gave written informed consents, which conformed to the Declaration of Helsinki and the study was approved by the Northern B Health and Disability Ethics Committee of New Zealand (14/NTB/113/AM03).
Experimental procedure
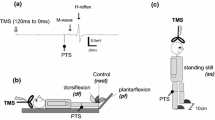
Each subject had their H-reflex and M-wave recruitment curves recorded in their right soleus muscle three times. This was done in three different body postures: while the subject standing on their left leg with the right leg loosely hanging to the side; sitting on a high chair without feet touching to the ground; and lying prone with the subjects’ feet over the edge of the bench. In all postures, the aim was to prevent any activity on the soleus muscle and any involvement of the skin receptors on the sole of the right foot. The order of the different positions was randomized, and the subjects were instructed to relax their muscles while they received electrical stimulations. Subjects were instructed to keep the head position steady throughout the experiment to avoid neck reflexes from confounding the current study (Traccis et al. 1987).
Recording
Bipolar surface electrodes (20 mm Blue Sensor Ag/AgCl, AMBU A/S, Denmark) were placed on the right soleus muscle (SOL) 2 cm below the lateral gastrocnemius muscle and 2 cm apart (bipolar configuration). A ground electrode was placed on the malleolus at the ankle of the same leg. The surface electromyography (SEMG) signals were amplified and recorded using a CED Power 1401 MK 2 data acquisition board at 5 kHz and band-pass filtered at 20–1000 Hz.
Electrical stimulation
The H-Reflex and M-wave of the soleus muscle were elicited by stimulating the tibial nerve. The electrical stimulations were delivered via an isolated stimulator (Digitimer DS7AH, UK). The stimulating electrodes (PALs platinum rectangular electrode, 75 × 100 mm; Axelgaard Man) at the anode end and a custom made silver disc with a diameter of 10 mm at the cathode. The anode was placed proximal to the patella and the cathode was placed on the tibial nerve in the popliteal fossa both on the right leg. The exact position of the cathode and the intensity of the stimulus needed to elicit a response was manipulated until the greatest H-reflex response with a minimum stimulus intensity was achieved.
Establishment of H-reflex and M-wave recruitment curves
With the cathode set on the optimum position for the H-reflex response, the intensity needed to achieve an M-max was found at each of the three different body positions. The M-max was found by progressively increasing the stimulation intensity by 5 mA and delivering three stimulations at each intensity. When the peak-to-peak amplitude of the M-wave did not change over three consecutive trials, the current intensity of the previous stimulation was considered as the 100% stimulus intensity for the M-max.
To construct the H and M recruitment curves, the stimulus intensity that achieved M-max was divided into 16 equal steps between zero and M-max. Each of the 16 levels of intensities were delivered three times at a varying time intervals between 2 and 3 s in a random order, a total of 48 randomized stimulations for each of the three positions. After the delivery of 48 stimuli lasting for two minutes, subject rested for five minutes and changed his/her body posture for the next 48 stimuli. Using this approach the stimulus intensity was also normalized according with the intensity that induced M-max (method of Brinkworth et al. 2007).
Data analysis
For the evoked potentials, peak-to-peak amplitude of the H-reflex and M-wave was computed offline from the unrectified SEMG signals. To reduce inter-subject variability, H- and M-waves were normalized to the corresponding maximal M-wave (M-max), thus the H-max/M-max ratio was computed. For each recruitment curve, the current intensity at H-max and M-max was identified.
The ascending part of recruitment curve was fitted by a general least squares model, as described by (Brinkworth et al. 2007). From the curve fit analysis, the following parameters were analyzed: current intensity at H-reflex threshold, current intensity at 50% of the H-max; and the slope of the ascending limb of the recruitment curve at 50% of the H-max (Hslope). The dependent measures were; threshold of H-reflex, M-wave at 50%, M-wave slope, H-reflex area under curve normalized to M-max and H-max.
Surface EMG background level
Average rectified value (ARV) of the SOL SEMG was estimated from each sweep for an epoch of 450 ms preceding the stimulation and then integrated, after all of the 48 sweeps were computed the result was averaged.
Statistical analysis
All changes were evaluated using one-way ANOVA with factor condition (Prone, sitting, and standing). Post hoc pairwise comparison was done using Tukey’s HSD tests if required. Statistical significance was set at P < 0.05 for all comparisons.
Results
The H/M curves were obtained in 14 healthy informed volunteers. Peak-to-peak values of the H-reflex and M-wave were calculated for each of the 16 stimulus intensities and for each of the three body positions. Stimulus–response curves were constructed that normalizes the amplitude of the H-reflex to the maximal M-wave and also normalizes the stimulus intensity to the current that induces maximal M-wave. With these ‘double’ normalization procedures (see Methods for the details) M-wave curves for all body positions superimposed, hence allowing any shifts in the H-reflex curves to be distinguished clearly. This unique approach also allows the results of different subjects to be pooled so that average response from a group of individuals to be illustrated.
This is exactly what is shown in the first figure. Figure 1a shows the pooled results for all 14 experiments. We also illustrate results from an individual subject for comparison (Fig. 1b).

The M-wave and H-reflex curves for the experiment. The traces are the curves depicting average changes in all three positions (Standing, Sitting, and Prone) for a all subjects; b one subject
Figure 2 illustrates the changes in the H-reflex (area under the curve and threshold) and M-wave parameters. There were no significant differences between the parameters for most of the conditions: H-reflex area under the curve (p = 0.21); threshold for eliciting H-reflex (p = 0.67); M-wave at 50% (p = 0.22); M-wave slope (p = 0.34) and average rectified value (ARV) of the prestimulus background surface EMG (p = 0.11).

Effect of the positions (Standing, Sitting and Prone) on the H-reflex parameters (the normalized H-reflex, the H-reflex threshold, the M-wave s50, and the M-wave slope)
The only exception for this rule was the size of the maximal H-reflex response (H-max). Normalized H-max in prone position was 0.49 ± 0.23 as compared with sitting 0.32 ± 0.17 and standing 0.30 ± 0.14 positions. Hence, H-max was significantly larger in the prone posture compared with both sitting (p = 0.02) and standing (p = 0.01) postures (Fig. 3).

Effect of the position (Standing, Sitting and Prone) on the normalized H-max values. Normalized H-max value in the prone position was significantly higher than the values in sitting (p = 0.02) and standing (p = 0.01) positions
Discussion
The aim of the current study was to examine any potential posture related changes in the effectiveness of the spindle primary afferent synapse on the motor neurons of the soleus muscle. To achieve this aim, standardized H-reflex methodology was utilized. H-reflexes were induced in three different body positions; prone, sitting and standing. While inducing the H-reflex, three different standardizations were applied to obtain reliable results; amplitude normalization to M-max, stimulus normalization to the stimulus intensity that generated M-max, and background EMG level normalization preceding the stimulus.
The results indicated that the efficacy of spindle primary afferent synapse on the soleus motor neuron pool changes with a change in the body posture, by becoming significantly stronger in the prone position. However, we must stress that the H-reflex results were only significant for the H-max values. The body posture did not significantly alter the values for the H-reflex area under the curve and the threshold for eliciting H-reflex.
We speculate that this discrepancy may be due to the distribution of presynaptic inhibition (PSI) on different sized motor neurons. It is possible that the PSI originating from the vestibular apparatus and other posture activated proprioceptors on the spindle primary afferent synapses is differentially distributed and is stronger on the larger motor neurons compared with the smaller motor neurons. We reason this speculation to the following rationale:
-
1.
We know that the spindle primary afferent synapses inject twice as much current into the smaller motor neurons compared with the larger ones (Heckman and Binder 1988). This differential distribution strengthens the size principle of recruitment and makes sure that under the circumstances where muscle spindles are contributing to the recruitment of motor units the small size motor units will be recruited earlier.
-
2.
This differential distribution of spindle primary afferent synapses has been suggested to be compensated for by a weaker PSI originating from the Ia fibers on the spindle primary afferent synapses on larger motor units compared with the smaller ones (Aimonetti et al. 2000).
-
3.
Although we have no direct evidence on the distribution of PSI originating from the vestibular apparatus / proprioceptors on the spindle primary synapse, it is possible that they may be also be differentially distributed but this time to be more effective on the larger motor units. If the PSI from these posture activated receptors are distributed to be more effective in the larger motor units, their release during lying would generate larger H-reflex responses from the larger motor units, hence increasing the size of the H-max.
The literature on this subject is vastly variable, as three different findings have been reported. While some research groups claimed that the H-reflex in the laying position was weaker than the sitting or standing positions (Knikou and Rymer 2003; Aiello et al. 1983; Chan and Kearney 1982), others suggested that the H-reflex in the laying position was stronger than when it is elicited in other positions (Shimba et al. 2010; Hayashi et al. 1992; Koceja et al. 1993; Fox and Koceja 2017). There is another group of researchers, however, who found no change in the H-reflex in different body postures (e.g., Trimble 1998). The likely factors that may be responsible for such varying results are discussed below. The current findings in the light of such confounding factors are discussed.
Most of the work was performed on the lower leg muscles, as it is easier to elicit the H-reflex in these muscles. Therefore, the chosen muscle is not the problem for these varying results. Furthermore, most researchers used similar stimulating and recording techniques, and hence these should also not be responsible for the vastly different results found.
Level of prestimulus background muscle activity
Since it is well established that the level of background electrical activity of a muscle influences the size of the reflex response recorded (Miles and Türker 1986; Türker 1988; Stein et al. 2007) most researchers seriously considered this issue while studying the H-reflex (Kyung-Min et al. 2013). To achieve this, some researchers controlled the level of background muscle activity using various feedback methods (Kyung-Min et al. 2013; Cattagni et al. 2014). Others normalized the reflex response to similar EMG levels (Mynark and Koceja 1997). Still others used leg devices such as orthosis that reduced the increase in the EMG level in the standing position (Shimba et al. 2010). These EMG normalizations were done “in order to remove the effects of background muscle activity from the obtained H-reflex” Kawaishi and Domen (2016).
However, despite these warnings many researchers have taken little consideration regarding the background level of muscle activation into account. Even though it is known that the background EMG level of the posture muscles does change during standing compared with laying (Mynark and Koceja 1997) and it becomes about 10% of the MVC activity levels (Billot et al. 2010), some researchers only instructed the subjects to “...relax throughout the experiment” (Shimba et al. 2010) or stated that “No spontaneous EMG activity of the tibialis anterior, medialis gastrocnemius or soleus muscles was present” (Knikou and Rymer 2003) when studying the H-reflex under various body positions. It is therefore possible that muscle activation levels were not as carefully controlled as could be. In the current experiments, the level of background activity of the muscle was minimized using a special protocol (Methods) to overcome the postsynaptic influences on the effectiveness of spindle primary afferent synapse on the motor neuron.
Receptor activity from the soles of the feet
During upright standing, cutaneous mechanoreceptors from the soles of the feet are expected to be activated as they respond to the mechanical stimulation on them (Trulsson 2001; Strzalkowski et al. 2015). When activated, these receptors form multisynaptic reflex connections with the motor neurons of the lower leg muscles (Aniss et al. 1992) and also influence the discharge pattern of the spindle afferents from the these muscles (Aniss et al. 1990). Therefore, it is expected that in the standing and sitting positions where the mechanical pressure from a part or all of the weight of a person stimulates the receptors in the soles of the feet, and that this would influence the synapses on the motor neurons that supply the muscles of the lower leg.
During the current study this was carefully controlled for. The weight of the subject was not applied on the right foot at all so that any synaptic/presynaptic effect that may originate from the tonic activation of mechanoreceptors in the sole of the foot was prevented.
Normalizing the reflex findings to the stimulus intensity
Researchers were aware of the changes in the stimulating electrode against the nerve when the body posture was changed and attempted to normalize the stimulus intensity to a M-wave value that induces a percentage of the M-max amplitude (Fox and Koceja 2017; Knikou and Rymer 2003). This approach is justifiable since it insured that each stimulus activates a percentage of the motor axons, and hence any change in relative position between the stimulating electrodes and the nerve could be accounted for.
Besides using a stimulus intensity that insures delivery of similar stimulating currents to the nerve, in most of the previous studies on the H-reflex modulation with body posture the H-reflex amplitude was normalized to the maximal M-wave; and thus H/M curves were established (Mynark 2005; Cattagni et al. 2014). However, in most of the previous studies in this area of research such a normalization of the H/M curves to the stimulus intensity was not performed.
In 2007, Brinkworth and colleagues established a methodology where the normalization can be done not only for the M-max amplitude but for the stimulus intensity that induces the M-max (Brinkworth et al. 2007). This standardization procedure insures that the M-wave curves belonging to different experiments/subjects are superimposed. This procedure allows any shift on the H-reflex curve to be clearly illustrated. It is then possible to pinpoint any changes in the threshold for activation or the effectiveness of the Ia synapse on the motor neuron pool via examining the shifts in the H-reflex curve relative to the M-wave curve.
In this study, all these standardization methods were used, including establishing H-reflex / M-wave recruitment curves (Brinkworth et al. 2007). Using this method, a curve was fitted on the raw values of the H-reflex and the M-wave. Since the M-wave was normalized to its maximum size on the ordinate and the stimulus intensity that induced 100% M-wave on the axis, M-wave curves superimpose on top of each other when varying body posture experiments were performed. In this way, it is possible to determine any change in the position and area under the curve of the H-reflex that would indicate changes in effectiveness of the Ia synapse on the homonymous motor neurons.
Vestibular and other tonically active receptor systems and their influence on the effectiveness of spindle primary afferent synapses on soleus motor neurons
Since changing the body posture also involves changes in the activity of the vestibular apparatus it is difficult to separate their various effects. To study the effect of vestibular input on the excitability of motor neurons that innervate lower leg muscles Dakin et al. (2016) stimulated the vestibular apparatus and showed that it influenced the discharge activity of them. In particular, galvanic stimulation affected the motor neurons that innervate the medial gastrocnemius muscles more than the motor neurons that innervate the soleus muscle.
Lowrey and Bent (2009) studied the effect of galvanic stimulation of the vestibular apparatus on the H-reflex and found that such a stimulus increased the reflex by as much as 20%. Scarpini et al. (1991) stimulated the vestibular apparatus by a movement of a rotating chair and illustrated that the H-reflex increased in relation with the velocity and acceleration, and concluded that the changes described in motoneuron excitability could represent a vestibulospinal reflex response originating from the horizontal semicircular canals. Therefore, it is clear that the vestibular apparatus does influence both the excitability of motor neurons and effectiveness of the spindle primary afferent synapse on these motor neurons.
Since the current experiments controlled for all the above mentioned parameters except the tonic input from many receptor systems including the vestibular apparatus and the receptors that may be differentially activated at varying body postures such as the cutaneous receptors, it is concluded that the tonic input from these receptors may presynaptically interfere with the effectiveness of the spindle primary afferent synapses on the soleus motor neurons.
Limitations of the study
We used three different body postures that may affect the tonic discharges from not only the vestibular apparatus but also from the cutaneous, joint and muscle receptors. Although our protocol limited the electrical activity of right leg muscles, discharges from the proprioceptors especially from the left leg (during one-leg standing) and discharges from cutaneous receptors (during prone and sitting positions) may also presynaptically affect the effectiveness of the spindle primary afferent synapse on the motor neurons.
Conclusion
To resolve the conflicting results in the literature regarding the effect of static body position on the H-reflex, the standardized method put forward by Brinkworth et al. (2007) was utilized in this study. Other confounding factors were also controlled for, such as the prestimulus background activation of the muscle and the natural stimulation of the mechanoreceptors in the soles of the feet. The results indicated that the efficacy of spindle primary afferent synapse on the soleus motor neuron pool does change with the change in the body posture becoming significantly stronger in the prone position. We speculate that this change can be due to the differential distribution of PSI from the vestibular apparatus and the proprioceptors on spindle primary synapse. If the PSI from these posture activated receptors are distributed to be stronger on larger motor units, its release in lying posture would generate larger H-reflex responses from the large motor units, hence increasing the size of the H-max.
References
Aimonetti JM, Vedel J-P, Schmied A, Pagni S (2000) Distribution of presynaptic inhibition on type-identified motoneurones in the extensor carpi radialis pool in man. J Physiol 522:125—135
Aniss AM, Diener HC, Hore J, Burke D, Gandevia SC (1990) Reflex activation of muscle spindles in human pretibial muscles during standing. J Neurophysiol 64:671–679
Aniss AM, Gandevia SC, Burke D (1992) Reflex responses in active muscles elicited by stimulation of low-threshold afferents from the human foot. J Neurophysiol 67;1375–1384
Billot M, Simoneau EM, Van Hoecke J, Martin A (2010) Age-related relative increases in electromyography activity and torque according to the maximal capacity during upright standing. Eur J Appl Physiol 109:669–680
Brinkworth RSA, Tuncer M, Tucker K, Jaberzadeh S, Türker KS (2007) Standardization of H-reflex analyses. J Neurosci Meth 162:1–7
Cattagni T, Martin A, Scaglioni G (2014) Is spinal excitability of the triceps surae mainly affected by muscle activity or body position? J Neurophysiol 111:2525–2532
Dakin CJ, Heroux ME, Luu BL, Inglis JM, Blouin J-S (2016) Vestibular contribution to balance control in the medial gastrocnemius and soleus. J Neurophysiol 115:1289–1297
Fox A, Koceja D (2017) Static otolithic drive alters presynaptic inhibition in soleus motor pool. J Electromyogr Kinesiol 32:37–43
Heckman CJ, Binder MD (1988) Analysis of steady-state effective synaptic currents generated by homonymous Ia afferent fibers in motoneurons of the cat. J Neurophysiol 60:1946–1966
Knikou M (2008) The H-reflex as a probe: pathways and pitfalls. J Neurosci Meth 171:1–12
Knikou M, Rymer WZ (2003) Static and dynamic changes in body orientation modulate spinal reflex excitability in humans. Exp Brain Res 152:466–475
Kyung-Min K, Hart JM, Hertel J (2013) Influence of body position on fibularis longus and soleus Hoffmann reflexes. Gait Posture 37:138–140
Lowrey CR, Bent LR (2009) Modulation of the soleus H-reflex following galvanic vestibular stimulation and cutaneous stimulation in prone human subjects. Muscle Nerve 40:213–220
Miles TS, Türker KS (1986) Does reflex inhibition of motor units follow the “Size Principle”? Exp Brain Res 62:443–445
Morita H, Petersen N, Christensen LO, Sinkjaer T, Nielsen J (1998) Sensitivity of H-reflexes and stretch reflexes to presynaptic inhibition in humans. J Neurophysiol 80:610–620
Mynark RG (2005) Reliability of the soleus H-reflex from supine to standing in young and elderly. Clin Neurophysiol 116:1400–1404
Scarpini C, Mazzocchio R, Mondelli M, Nuti D, Rossi A (1991) Changes in alpha motoneuron excitability of the soleus muscle in relation to vestibular stimulation assessed by angular acceleration in man. ORL J Otorhinolaryngol Relat Spec 53:100–105
Schieppati M (1987) The Hoffmann reflex: a means of assessing spinal reflex excitability and its descending control in man. Prog Neurobiol 28:345–376
Shimba S, Kawashima N, Ohta Y, Yamamoto S-I, Nakazawa K (2010) Enhanced stretch reflex excitability in the soleus muscle during passive standing posture in humans. J Electromyogr Kinesiol 20:406–412
Stein RB, Estabrooks KL, m McGie S, Roth MJ, Jones KE (2007) Quantifying the effects of voluntary contraction and inter-stimulus interval on the human soleus H-reflex. Exp Brain Res 182:309–319
Strzalkowski ND, Mildren RL, Bent LR (2015) Thresholds of cutaneous afferents related to perceptual threshold across the human foot sole. J Neurophysiol 114:2144–2151
Traccis S, Rosati G, Patraskakis S, Bissakou M, Sau GF, Aiello I (1987) Influences of neck receptors on soleus motoneuron excitability in man. Exp Neurol 95:76–84
Trimble MH (1998) Postural modulation of the segmental reflex: effect of body tilt and postural sway. Int J Neurosci 95:85–100
Trulsson M (2001) Mechanoreceptive afferents in the human sural nerve. Exp Brain Res 137:111–116
Türker KS (1988) A method for standardization of silent period measurements in human masseter muscle. J Oral Rehab 15:91–101
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Cecen, S., Niazi, I.K., Nedergaard, R.W. et al. Posture modulates the sensitivity of the H-reflex. Exp Brain Res 236, 829–835 (2018). https://doi.org/10.1007/s00221-018-5182-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-018-5182-x