
Abstract
This paper examines up to third-order geometric properties of wrist path and the first-order property of wrist trajectory (wrist speed) for spatial pointing movements. Previous studies report conflicting data regarding the time invariance of wrist-path shape, and most analyses are limited to the second-order geometric property (straightness, or strictly speaking, curvature). Subjects performed point-to-point reaching movements between targets whose locations ensured that the wrist paths spanned a range of lengths and lay in various portions of the arm’s spatial workspace. Movement kinematics were recorded using electromagnetic sensors located on the subject’s arm segments and thorax. Analysis revealed that wrist paths tend to lie in planes and to curve more as movement speed decreases. The orientation of the wrist-path plane depends on the reaching task but does not vary significantly with movement speed. The planarity of wrist paths indicates that the paths have close to zero torsion—a third-order geometric property. Wrist-speed profiles showed multiple peaks for sufficiently slow and long lasting movements, indicating deviation from the well-known, bell-shaped profile. These kinematic findings are discussed in light of various motor control theories.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Bernstein (1967) first described the redundancy problem that the human central nervous system (CNS) must resolve for movement control. For arm movement, the three-degree-of-freedom task of positioning the wrist in space is redundant in that it requires control of (at least) four joint variables—three characterizing shoulder movement and one characterizing elbow movement. The optimal control view of motor control holds that the CNS resolves motor redundancy by optimizing some measure of movement cost (Todorov 2004), and several kinematic measures such as minimum-jerk (Flash and Hogan 1985) and dynamic criteria such as muscle fatigue (Prilutsky and Zatsiorsky 2002) have been experimentally verified. Observed consistencies or invariant attributes of movement are often critical in the identification of these potential cost functions. For example, the concept of minimizing the net jerk of the hand trajectory directly arose from the desire to explain straight-line wrist paths in horizontal reaching (Flash and Hogan 1985). Identifying invariant movement characteristics is therefore important, and this paper focuses on identifying geometric invariant characteristics in spatial reaching motion.
A host of motion studies have identified a number of invariant characteristics in human reaching. Wrist-path shape in pointing movements has been shown to be independent of movement speed in horizontal-plane (Morasso 1981; Soechting et al. 1995) and vertical-plane (Atkeson and Hollerbach 1985) studies. Paths were straight in the horizontal plane and curved in the vertical plane. The tangential speed of the hand has a single, bell-shaped curve regardless of its magnitude (Atkeson and Hollerbach 1985; Flash and Hogan 1985; Morasso 1981; Soechting and Lacquaniti 1981). In handwriting and drawing, tangential velocity is proportional to the radius of curvature of the trajectory such that the angular velocity takes on several distinct, constant values during the movement and the corresponding movement segmentation is independent of the total duration—the so-called two-thirds power law (Lacquaniti et al. 1983; Viviani and Terzuolo 1982). Piano playing is another skilled task in which the set of ratios between interstroke intervals is independent of the duration (Soechting et al. 1996). Human subjects proved to be incapable of adapting to robot-applied force fields that depended explicitly on time rather than motion variables (Conditt and Mussa-Ivaldi 1999), but successfully generalized adaptation to motion-variable-dependent fields across different movements to the same regions of the fields (Conditt et al. 1997).
The above evidence for invariant kinematic patterns in planar motion is substantial and mostly convergent. A notable exception is the work of Papaxanthis et al. (2003), which reports a speed effect on wrist-path shape during sagittal-plane reaching. For spatial reaching, however, the evidence is not convergent. Schaal and Sternad (2001) demonstrated systematic violations of the two-thirds power law for unconstrained, rhythmic, spatial arm movements. Adamovich et al. (1999), Nishikawa et al. (1999), and van der Well et al. (2010) found wrist-path shape in spatial reaching to be speed independent, whereas Breteler et al. (1998) report the opposite.
The invariant bell-shaped wrist-speed profile is a robust characteristic of arm movement that has been observed for both planar and spatial movement, but, similar to path geometry, exceptions can be found. Multi-modal wrist-speed characteristics close to movement termination is a well-studied phenomenon (Dounskaia et al. 2005; Meyer et al. 1988), and similar deviations have been observed for timed slow movements (Isenberg and Conrad 1994) and untimed movements for a rather limited set of spatial movements (Messier et al. 2003).
One possible explanation for the conflicting results regarding which motion characteristics actually are invariant with movement speed is that the pointing movements in previous spatial arm-motion studies (Adamovich et al. 1999; Breteler et al. 1998; Nishikawa et al. 1999; van der Well et al. 2010) encompass only limited portions of the workspace [see Ambike (2011) for details]. The present paper explores this potential explanation by seeking regularities in the output-space kinematics of spatial arm movements that encompass a large portion of the subject’s output space (see “Results” section for details). The target locations used in this work ensure that (a) the wrist paths are longer than in previous studies, (b) both initial and final target locations are changed across tasks, (c) movements involve different changes in wrist elevation so that the influence of gravity differs across tasks, and (d) movements require significantly different shoulder- and elbow-joint excursions (Ambike and Schmiedeler 2013).
Thus, the objectives of this research were to ascertain for spatial reaching motion (a) which, if any, geometric properties of the wrist path are invariant with movement speed and (b) whether the shape of the wrist-speed profile is invariant with movement speed. While not a comprehensive search for regularities, this work goes beyond the common practice in prior work to examine just up to the second-order geometric property of wrist-path curvature. Here, the third-order path property, torsion, is also analyzed.
Methods
Subjects and tasks
Nine subjects, five females and four males, with no history of physical or neurological disorders participated in the study. Eight subjects were right-handed, and one was ambidextrous. Subjects were between 20 and 33 years of age [mean = 23.22 years, standard deviation (SD) = 3.9 years] and naive to the purpose of the experiment. All subjects gave their informed consent prior to inclusion in the study. This research received approval from the appropriate Institutional Review Board.
The subjects sat in and were strapped to a chair with a band passing over the chest and under the arms to minimize movement of the thorax. Eight colored tennis balls serving as targets were arranged around the subject on stands made of PVC pipe, as shown in Fig. 1. Table 1 specifies the locations of the targets relative to the subject’s right shoulder (median values).

Location of targets relative to subject’s right shoulder. Tennis balls covered with colored paper serve as targets mounted on PVC pipe frames. Dotted lines indicate spatial locations of targets (color figure online)
The following eight pointing tasks, consisting of the subject pointing from an initial target to a final target with the right arm, were used in the study.
- Task 1::
-
\(T1 \rightarrow T8\), short contralateral movement, decreasing elevation.
- Task 2::
-
\(T3 \rightarrow T6\), long ipsilateral movement, increasing elevation.
- Task 3::
-
\(T1 \rightarrow T5\), short ipsilateral movement, horizontal.
- Task 4::
-
\(T2 \rightarrow T7\), long contralateral movement, decreasing elevation.
- Task 5::
-
\(T2 \rightarrow T6\), long contralateral movement, increasing elevation.
- Task 6::
-
\(T2 \rightarrow T5\), long contralateral movement, horizontal.
- Task 7::
-
\(T4 \rightarrow T7\), long contralateral movement, horizontal.
- Task 8::
-
\(T3 \rightarrow T7\), short ipsilateral movement, horizontal.
Tasks 1 through 6 are depictedFootnote 1 in Fig. 2. All eight tasks required movement away from the body and full extension of the elbow. Each began with the subject in the neutral position with the hands resting on the knees. The following instructions were given to the subject. The experimenter will name the initial target. Point to the initial target with your forefinger close to the target and remain in that position. The experimenter will then call out a speed cue, followed by the final target. The speed cue will be either ‘slow,’ ‘normal,’ or ‘fast.’ Interpret the speed cue in a consistent fashion for various tasks. Move your head to locate the position of the final target (if necessary) and then point to the final target at the appropriate speed. Move the entire arm, i.e., the shoulder and the elbow, such that the elbow is maximally extended in the final position. Remain in the final position until the experimenter says the word ‘neutral.’ Return to the neutral position and wait for the specification of the next task. Keep the wrist rigid, and focus more on producing smooth movements rather than the accuracy of the final position of the finger.

Two views of each pointing task, one from behind subject and one overhead (color figure online)
Each subject performed 220 pointing motions. Of these, the first 20 involved all tasks performed at all three speeds to acquaint the subject with the procedure. The actual experiment was comprised of the next 200 motions presented in pseudorandom order such that Tasks 1 through 6 were repeated ten times at each of the three speeds and Tasks 7 and 8 in combination were repeated a total of 20 times. To minimize fatigue, breaks of approximately 5 min were enforced after the 80th, 130th, 170th, and 200th motion. None of the subjects reported feeling fatigued during the experiment.
Data recording
Electromagnetic sensors (Flock-of-Birds, Ascension Technology Corporation) were taped onto the subject’s right wrist, right upper arm, right scapula, and spinous process of the seventh cervical vertebra. Sensor locations were related to various body-segment locations via a digitization protocol recommended by the International Society of Biomechanics (Wua et al. 2005). The X, Y, and Z position coordinates were sampled at 100 Hz and low-pass filtered using a fourth-order, zero-lag, Butterworth filter with a cutoff frequency of 1.5 Hz. The wrist position and velocity were computed in The MotionMonitor software (Innovative Sports Training, Inc.). To isolate the smoothest portion of the motion, movement onsets and end points were identified as the points in time when 10 % of the peak wrist speed attained during the particular trial was first reached (onset), and next reached (end point). All analyses were confined between these two points.
Data analysis
Wrist-path length
The mean and SD of the linear distances traveled by the wrist from the initial to the final target position were computed for each task. Data were pooled across subjects and speeds.
Average speed
To determine whether the speed commands elicited movements of distinct speeds, the average speed of each movement, defined as the wrist-path arc length divided by the movement time, was computed.
Wrist-path planarity
Figure 3 shows a hypothetical but typical wrist path that resembles those observed in this study. Wrist paths appear as nonlinear curves that tend to lie in ‘wrist planes’ that have variable orientations relative to the horizontal plane. Therefore, principal component analysis (PCA) was used to fit planes to the 3D wrist-position data. The PCA approach to obtain a linear regression model is appropriate because the wrist-position coordinates cannot be naturally categorized into predictor and response variables. The eigenvectors of the covariance matrix of the data set comprised of the wrist positions for a pointing task are the principal components (PCs) for that task. The first PC points roughly in the direction of the pointing movement, the second in the direction of the greatest deviation from the direction of the pointing movement, and the third in the ‘out-of-plane’ direction. The first two PCs define the plane of movement, termed the wrist plane, which is also the best-fit plane in a least-squares sense.

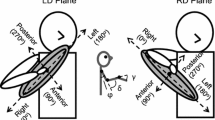
The plane of normals is viewed along the movement-direction vector and from the wrist-path end point. The bidirectional green line in the plane of normals represents the two choices for the planarized normal to characterize the wrist-path plane (color figure online)
The eigenvalues of the covariance matrix describe the amount of variance explained by the corresponding PC. Therefore, the sum of the two largest eigenvalues obtained from the PCA serves as a goodness-of-fit metric. A value of one indicates planar data, and it decreases for nonplanar data. Another measure of the fit is the ratio of the mean absolute distance of data from the fit plane to the linear distance between the end points of the path (Breteler et al. 1998). This metric will be zero for planar data, and it increases for nonplanar data. Both metrics were computed.
Wrist-path linearity
The spatial wrist paths were projected onto the corresponding wrist planes, and a scale-invariant linearity metric was applied to quantify the global curvature of the planarized wrist paths. The linearity metric, introduced by Atkeson and Hollerbach (1985) and employed in other studies (Adamovich et al. 1999; Nishikawa et al. 1999), is the ratio of the maximum normal distance between a wrist path and a straight line joining the initial and final points on the path to the length of that line. The metric value is zero for a straight-line wrist path and increases with deviation from a straight line. Note that here ‘linearity’ and ‘curvature’ refer to opposite trends describing the same second-order property of the path.
Orientation of the wrist planes
Each wrist plane is characterized by a (unit) vector normal to it given by the third PC of the corresponding wrist-path data. Ideally, for all repetitions of a task, the normals would lie in a single plane called the plane of normals. This plane in turn is characterized by the unique movement-direction vector that points along the line joining the initial and final points on the wrist path, as shown in Fig. 3. However, there is some variability in the orientation of the individual movement-direction vectors of all repetitions of a task. The average of all movement-direction vectors across all subjects and speeds serves as the movement-direction vector representing the particular task, as shown in Fig. 4, and defines the plane of normals. All individual wrist-plane normals are projected onto the plane of normals and are called the planarized normals. The planarized normals for a given task are chosen such that the variability in the angles between these normals is minimized. The choice is between a normal and its negative, both of which characterize the same wrist plane, as shown in Fig. 3. The gravity vector is also projected onto the plane of normals. Then, the normal angle is the angle from the projected gravity vector to a planarized normal in the plane of normals, measured positive counterclockwise when viewed from the wrist-path end point, and it quantifies the relative orientation of the wrist plane. The projected gravity vector is a consistent datum for measuring the normal angles so that they can be compared across tasks. Figure 4 illustrates this procedure for all repetitions of one task.

Movement-direction vectors (blue dashed lines) and planarized normals (green solid lines) for Task 5, all subjects and speeds. The black thick line is the average of the movement-direction vectors and represents the movement direction for the task. It also defines the plane of normals onto which the wrist-plane normals are projected. The red solid line is the gravity vector, acting downward, and the red dotted line is its projection onto the plane of normals (color figure online)
Wrist-speed profile
All wrist-speed profiles were normalized for peak wrist speed and movement time, and the average of all 60 normalized wrist-speed profiles of the same speed was constructed as the reference profile for that speed. The deviation of each of the 60 wrist-speed profiles from the reference profile was calculated with the similarity metric developed by Atkeson and Hollerbach (1985). Let A and B be the areas under the reference profile and an experimental profile, respectively. Then, A∪ B is the total area contained beneath both curves, and A ∩ B is the area common to both curves. The similarity metric is
The value of w is zero for identical profiles and increases as the profiles become more dissimilar.
A local maximum in the wrist-speed curve was considered a peak in the profile. This definition was used to determine the number of peaks in the wrist-speed profiles.
Statistical analysis
The linearity metric was subjected to a two-way repeated-measures ANOVA (TASK × SPEED), and the average speed and similarity metric were subjected to one-way repeated-measures ANOVAs with SPEED as a factor. Data were pooled across subjects. Post hoc analysis was conducted when necessary using pairwise comparisons with Bonferroni corrections. All statistics were performed with SPSS statistical software using an α-level of 0.05. Mauchly’s sphericity tests were performed to verify the validity of using repeated-measures ANOVA. The Greenhouse–Geisser adjustment to the degrees of freedom was applied whenever departure from sphericity was observed. The linearity metric was also regressed against the movement time.
The normal angles are drawn from a semicircular distribution since the normal vector and its negative define the same plane. To apply circular statistical methods, they are multiplied by a factor of 2 (Jones 1968). The Rayleigh test for circular uniformity was used to ensure that the (doubled) normal angles belonged to a unimodal distribution. The main effects of SPEED and TASK on the (doubled) normal angles were investigated by using the Watson–Williams procedure for multisample testing (Zar 1999).
Results
Wrist-path length
The linear distances traveled by the wrist for Tasks 1 through 6 are (mean ± SD) 0.49 (±0.06), 0.92 (±0.07), 0.39 (±0.05), 0.85 (±0.08), 0.77 (±0.07), and 0.83 (±0.09) m, respectively.
Average speed
The speed commands elicited movements with distinct average wrist speeds.
Significant effect of SPEED was observed on the average speed [F (2,16) = 147.7, p < 0.01]. Post hoc tests revealed that average speeds for all levels were significantly different from each other. The mean (± SD) values were 0.29 ± 0.2, 0.48 ± 0.3, and 0.72 ± 0.3 m/s for the slow-, normal-, and fast-speed movements, respectively.
Wrist-path planarity
Wrist paths for spatial pointing movements tend to lie in planes.
The goodness of fit obtained from PCA was greater than 0.996, and the maximum mean deviation from the fit plane was less than 2.5 % of the linear distance between the path end points for all tasks and subjects.
Wrist-path linearity
Wrist paths for a spatial pointing task tend to curve more for slower and longer-lasting movements. Therefore, wrist paths are not speed invariant, in general.
The two-way ANOVA revealed significant effects of SPEED [F (2,16) = 35.27, p < 0.01) and TASK [F (5,40) = 23.53, p < 0.01]. Post hoc analysis revealed that faster speeds always produced significantly lower linearity metric values, and TASK effect was not significant for the pairs Task 1–Task 3, Task 2–Task 4, Task 2–Task 5, Task 2–Task 6, Task 4–Task 6, and Task 5–Task 6. The unbalanced effects of TASK and SPEED on the metric resulted in significant interaction between the factors [F (10,80) = 6.48, p < 0.01].
The linearity metric was regressed against the movement time. The error bar plots of the confidence intervals on the residuals (not included) showed a random distribution about a zero mean, validating the normality assumption for all 54 cases. Of these, 46 cases showed significant, negative correlation between linearity metric and movement time. This indicates that the wrist paths tend to get more curved as movements become slower and longer.
Orientation of the wrist-path plane
The orientation of the wrist-path plane is task dependent. However, it does not change with movement speed.
The maximum and mean (±SD) angular rotation for a normal during the projection onto the plane of normals were 26° and 4.2 (±3.9)°, respectively, for all subjects and tasks. These angles provide a sense of the variability in the movement direction. The loss of information associated with this projection is negligible.
The Rayleigh test confirmed that all samples were drawn from unimodal circular distributions. The Watson–Williams test failed to reveal a significant effect of SPEED [F (2,159) = 0.8, p = 0.45] and revealed a significant effect of TASK on the normal angles [F (5,156) = 22.03, p < 0.01]. The circular mean and SD of the (doubled) normal angles for slow-, normal- and fast-speed motions were −0.83 ± 0.9, −0.72 ± 0.9, and −0.5 ± 0.9 radians, respectively. These values were 0.40 ± 1.1, −0.91 ± 0.5, −0.43 ± 1.2, −0.90 ± 0.8, −0.72 ± 0.7, −0.73 ± 0.7 radians for tasks 1–6, respectively.
Wrist-speed profile
The variability in the shape of the normalized wrist-speed profile reduces as movement speed increases. The wrist-speed profile starts to deviate from the unimodal bell shape for low movement speed and long movement duration.
The metric w was significantly affected by SPEED [F (2,16) = 41.6, p < 0.01]. Post hoc tests revealed that the profiles became more uniform as movement speed increased. The metric values for slow movements were significantly different from the values for the other two speeds. The mean ± SD for w were 0.17 ± 0.1, 0.1 ± 0.09, and 0.09 ± 0.07 for the slow-, normal-, and fast-speed movements, respectively.
Figure 5 shows two kinds of wrist-speed profiles observed in this study. Normalized time and wrist speed are plotted on the x and y axes, respectively. All profiles for fast movements are unimodal and resemble the profile shape reported in the literature. Almost 50 % of the profiles for slow movements, and some profiles for normal-speed movements, show multiple peaks.

Individual plots of normalized wrist speed against normalized time. The plots demonstrate examples of two qualitatively different profiles of wrist speed, unimodal (a) and multimodal (b). The blue, red, and green curves represent fast-, normal-, and slow-speed movements, respectively. Subject JR (color figure online)
Examples of multimodal wrist-speed profiles were found for all subjects and for all tasks, suggesting that the phenomenon may be task independent. All subjects together performed 540 movements at each speed. Of these, 251 slow-speed movements, 39 normal-speed movements, and zero fast-speed movements were multimodal. Except for Task 3, the number of repetitions with multimodal profiles tends to be roughly independent of the task. The number also varies with subject and depends on the subject’s interpretation of ‘normal’ and ‘slow’ for performing those movements. Typically, slower movements that last longer are more likely to display multimodal wrist-speed profiles (Ambike 2011).
Figure 6 plots average movement speed against the movement time for all tasks and subjects, wherein tests that exhibit unimodal profiles are indicated as crosses and those with multimodal profiles are indicated as circles. Only unimodal profiles lie to the left and above the overlaid rectangle, and only multimodal profiles lie to the right of it. Both unimodal and multimodal wrist-speed profiles appear within the rectangle, so it represents a transition zone in the speed-movement duration space wherein the unimodal bell shape of the wrist-speed profile begins to break down. A rough characterization of the transition zone is as follows:
-
Movement duration between 1.5 and 3.7 s.
-
Average wrist speed less than ≈0.5 m/s.

Average wrist speed versus movement time for all tests and subjects. Crosses indicate tests with unimodal wrist-speed profiles, and circles indicate tests with multimodal profiles. The rectangle at the center of the figure describes a transition zone with both unimodal and multimodal profiles. The blue, red, and green colors represent fast-, normal-, and slow-speed movements, respectively (color figure online)
Discussion
There are four main findings of this study. First, wrist paths in spatial reaching movements tend to lie in planes. Second, the wrist-path shape is speed dependent. Third, the planes in which wrist paths lie are independent of movement speed. Fourth, the shape of the normalized wrist-speed profile, which is known to be a robust invariant characteristic of reaching motion, becomes variable as movement speed reduces, and it becomes multimodal for slow and long-lasting reaching movements. Thus, two speed-invariant wrist-path characteristics were obtained in this study: path torsion and the orientation of the wrist-path planes. These findings are now interpreted in the context of prior methods to characterize wrist paths and in light of current views on motor control.
Characterizing wrist-path geometry
Curves in three-dimensional space are characterized locally by their differential geometric properties. Position is the zeroth-order property, path tangent is the first-order property, path curvature is the second-order property, and torsion is a third-order property of a spatial curve. Footnote 2 Wrist paths are clearly three-dimensional curves, and their shape is characterized up to the third order in this paper, using nonlocal equivalents of the differential geometric properties. The first-order geometric property of the path, the movement direction, was a controlled variable. The second-order path property, path curvature, decreases as movement speed increases. Since wrist paths were planar, the third-order path property, path torsion (‘out-of-plane’ movement of a curve), was close to zero. Most prior work on wrist path concentrates on the second-order property, path curvature.
Speed-dependent wrist-path shapes contradict the observations of Adamovich et al. (1999) and Nishikawa et al. (1999) who use the same linearity metric, but it supports the findings of Breteler et al. (1998) despite their use of a different metric. One reason why speed dependence was not observed in some previous work may be the lengths of the reaching paths studied. There is significant variation in the lengths of the various tasks in this study (see “Results” section), and four of the six tasks in this study are longer than any task required in previous spatial reaching studies (Adamovich et al. 1999; Breteler et al. 1998; Haggard and Richardson 1985; Nishikawa et al. 1999; van der Well et al. 2010). The target placement in the present study also requires movement through a larger region of the subject’s workspace than previous studies. More details regarding the comparison of the reaching tasks in prior studies are available in Ambike (2011).
Planarity of wrist paths has been observed before by Atkeson and Hollerbach (1985) and Breteler et al. (1998). However, in Atkeson and Hollerbach (1985), the target placement alone explains arm movement being predominantly in a para-sagittal plane. In Breteler et al. (1998), the arm movements were less varied compared to those in this study, and screens placed within the subject’s workspace constrained the movements. Thus, the planar nature of wrist paths for unconstrained, spatial pointing motions is established herein for movements spanning a large portion of the subjects’ extra-personal space.
What affects the wrist-path shape?
As per one opinion in the motor control community, some observed characteristics of human movement, such as the wrist-path shape, are consequences of a process of optimization of some perceived cost of movement (Todorov 2004). Another opinion is provided by Loeb (2012) who points out that movement organization may resemble a constraint-satisfaction process more than an optimization process; animals may be motivated by ‘good-enough’ rather than optimal behavior. From this point of view, one can argue that the speed dependence of the wrist path is an ‘acceptable’ deviation from some optimal path shape (which may be explicitly specified, like a minimum-jerk solution, or a consequence of some other optimal policy), since the effects of these deviations are not large enough to affect the task, damage the hand, demand too much energy, etc.
Several motor control theories postulate that the CNS specifies an ideal wrist trajectory for reaching movements. According to the time-invariance hypothesis (Ambike and Schmiedeler 2006), the CNS specifies the wrist path to which the movement timing is added, and the resulting trajectory is then tracked by specifying muscle forces that are computed by internal models in the CNS. Internal models are neural mechanisms that mimic the input-output relations of the motor apparatus (Atkeson 1989; Kawato 1999). Alternatively, according to the referent-configuration hypothesis, which is an extension of the equilibrium-point hypothesis (Feldman 1986, 2011), observed movements are consequences of shifts in referent body configurations (i.e., a referent trajectory) and environmental factors. Referent configurations are defined as configurations at which all muscles are at their respective activation thresholds via the tonic stretch reflex.
If the variable path curvature is indeed a deviation from some ideal path/trajectory, one possible cause for this deviation is gravity. Gravitational influence is movement-speed dependent, with the limb segments experiencing larger gravitational impulses (time integral of gravitational force) in slower movements. These are coupled with comparatively small inertial accelerations for slow movements. It has been suggested (Atkeson and Hollerbach 1985; Hollerbach and Flash 1982; Soechting et al. 1986) and experimentally verified (Flanders and Herrmann 1992; Yadav 2010) that the CNS computes the gravity- and motion-dependent muscle forces separately. Additional evidence is provided by Gentili et al. (2007) who show that opposite kinematic patterns emerge for upward and downward arm movement, suggesting that the motor plan is strongly dependent on the direction of gravity, exploiting it to decelerate upward movements near their end point and to reduce muscle force required for downward movements. The same asymmetry is not seen in horizontal reaching or in vertical reaching in the absence of gravity (Papaxanthis et al. 2005). Therefore, the wrist-path curvature patterns may result from the inability of the CNS to accurately compensate for gravity effects, or from the purposeful exploitation of gravity to minimize muscular effort.
Note that since the slow movements are considered deviant and since the slower paths are more curved, the composed wrist path must be straight. This prediction is consistent with the observed, speed-invariant, straight wrist paths in gravity-free, horizontal-plane reaching studies (Morasso 1981; Papaxanthis et al. 2005; Soechting et al. 1995), as well as the work of Wolpert et al. (1995) who showed that the desired trajectory for two-joint planar reaching is a straight hand path in the visual space.
Alternatively, for the referent-configuration hypothesis, gravity effects constitute an environmental factor that interacts with the system to produce the observed wrist path. It is argued that greater interaction of gravity with the system for slower movements leads to greater wrist-path curvature. Again, the (ideal) referent trajectory must be straighter than the observed one.
Another explanation is provided by Dounskaia (2007) who suggested that invariants in hand motion (the straight path, bell-shaped velocity profile, etc.) are a consequence of the geometry of the arm and of sinusoidal joint motions. Deviations of joint motions from sinusoids may account for the increased path curvature and multimodal speed profiles during slow movements observed in the present study.
Wrist-path planes
The tendency of wrist paths to lie in planes may also be a consequence of some optimal policy. The speed-invariance of the wrist-path planes is another interesting finding of this study that may be an artifact of the gravitational influences on reaching motion. These phenomena warrant further study.
Wrist-speed profiles
The occurrence of peaks in the wrist-speed profile close to movement termination is a well-studied phenomenon (Dounskaia et al. 2005; Meyer et al. 1988). In contrast, multiple peaks in the wrist speed appear far from movement termination in Fig. 5b. This phenomenon was not observed in previous work on spatial human reaching, possibly because the studied movements were not sufficiently slow and/or long-lasting. A detailed comparison of movement times and speeds for the present study and previous work is presented in Ambike (2011).
Multiple modes in wrist-speed profiles have been reported earlier for temporally constrained movements (Isenberg and Conrad 1994; van der Well et al. 2010). The conditions in the present study are different, though, since the movement time is chosen by the subject and not externally constrained. Messier et al. (2003) conducted a similar spatial reaching study with target placement in 3D space, but used movements that did not cover a large portion of the subject’s workspace. Therefore, the present study reports multimodal wrist-speed profiles for externally untimed arm movements in a large portion of the subject’s extra-personal space.
Multiple peaks in the wrist-speed profile indicate the presence of submovements. Liao et al. (1997) define a submovement as a brief episode in which the absolute velocity of movement increases and then decreases. Motions with submovements were previously observed for spatially constrained tasks in which subjects performed accurate, discrete, target-acquisition movements while simultaneously minimizing movement time. Fitts’ law (Meyer et al. 1990) captures a compromise that exists between movement speed and accuracy, and normative theories explaining Fitts’ law choose submovement durations that optimize the total movement duration and the variability in the final position (Liao et al. 1997; Meyer et al. 1990). The tasks in the present study do not resemble spatially constrained tasks because the motions were not performed to minimize total movement time. These motions resemble temporally constrained tasks in which subjects are required to perform accurate, discrete, target-acquisition movements in a self-selected time that was not rigorously controlled.
Meyer et al. (1988) define a submovement as any one of three events: wrist velocity zero-crossing from positive to negative (type 1), wrist-acceleration zero-crossing from negative to positive (type 2), and zero-crossing of the third wrist displacement derivative (jerk) from positive to negative values (type 3). Type 1 submovements are interpreted as an overshoot (typically close to the target), type 2 as undershooting and re-accelerating to the target, and type 3 as slight decreases in the rate of deceleration. The multimodal wrist-speed profiles for slow and long movements in this study do not represent type 1 submovements, since there are no velocity zero-crossings. These are wrist-acceleration zero-crossings, and since they occur far from the time of movement termination, the movements are re-accelerations toward the target.
Occurrence of submovements in these tasks is attributed to the workings of an internal clock. The CNS perceives longer time intervals with greater variability (Ivry and Hazeltine (1994). It is proposed that the subject estimates the total time for the current task based on the speed cue. For slow movements, this interval is often large. Consequently, the estimate is vague, and this translates into difficulty in selecting the average motion speed. If the subject’s selected average speed is low, the error may become apparent after crossing the first peak in the wrist-speed profile when the drop in speed is sensed, but the target is not sufficiently close. At this point, a submovement is appended to the first submovement, thus adding another peak to the wrist-speed profile.
A similar explanation is offered by van der Well et al. (2010) for the occurrence of submovements in temporally constrained motion at low metronome rates. The CNS can establish a pattern of occurrence of a periodic, external stimulus, such as a metronome, only when the stimuli occur ‘sufficiently’ fast. Consequently, human response to the external stimulus is anticipatory for the high-frequency stimuli, and a reaction otherwise (Engströ m et al. 1996). The movements in the present study were not timed with a metronome; therefore, the subjects were not reacting to an external, experimental stimulus after a movement had commenced. Regardless, the subjects produced submovements for several slow- and normal-speed movements.
Note that the idea of the CNS using an internal model for time estimation is consistent with the time-invariance hypothesis (Ambike and Schmiedeler 2006), which proposes that the CNS composes movement geometry and appends movement timing to the geometric plan. This explanation is similar to the theories of Meyer et al. (1990) and Liao et al. (1997). Both explanations hinge on the selection of movement time; however, the accuracy-influenced movement duration for spatially constrained tasks is near optimal, whereas in the present work, submovement timing is selected to meet a vague temporal constraint.
Optimization approach to motor control
The results of this work suggest the characteristics that the results of optimization procedures should display. Previously suggested objective functions should be tested for these characteristics, and/or new objective functions can be proposed.
It is relevant to mention here some drawbacks of the optimization approach to motor control. First, there is no accepted standard on what to consider a successful prediction of an optimal policy (Prilutsky and Zatsiorsky 2002). Secondly, if voluntary movements are ‘good-enough’ rather than optimal (Loeb 2012), then there is a low possibility of isolating the actual cost that the animal may be (almost) optimizing. (Presumably, ‘good -enough’ behavior is in an acceptable neighborhood of some optimal solution.) The results of this paper may eliminate some candidate cost functions (like, somewhat trivially, the minimum-jerk criterion which predicts straight-line wrist paths), but they do little else to single out the costs of movement that the CNS may be (almost) minimizing.
Conclusions
Geometrical analysis of the wrist movement during spatial reaching revealed that wrist paths tend to lie in planes whose orientation is movement-speed independent and the wrist-path shape within these planes is speed dependent, with slower movements being more curved. The wrist-speed profile resembles the classical bell shape for fast- and normal-speed pointing movements, but high variability in unimodal wrist-speed profiles and multimodal profiles was observed for long-lasting and slow untimed arm movements. Thus, two speed-invariant wrist-path characteristics were obtained in this study: path torsion and the orientation of the wrist-path planes. Explanations for most of the observed features of the wrist path and wrist speed are offered based on separate existing motor control theories. However, a unified explanation of all observed movement features is lacking. The dependence of wrist path on movement speed may be due to gravity effects which dominate the inertial effects at lower movement speeds. The variability in the wrist-speed profile could result from inaccuracies in the CNS’s estimate of longer time intervals.
Notes
Tasks 6 and 7 and Tasks 3 and 8 are similar. The wrist paths of these two sets of movements are similar, and so Tasks 7 and 8 are not included in the data analysis of this paper.
The rate of curvature change with respect to the arc length is the ‘in-plane’ third-order geometric property of a curve. This property has not been studied in this work or elsewhere in the motor control/biomechanics literature. Together with torsion, it provides a complete third-order local characterization of a spatial curve.
References
Adamovich SV, Berkinbilt MB, Fookson O, Poizner H (1999) Pointing in 3D space to remembered targets. II: effects of movement speed toward kinesthetically defined targets. Exp Brain Res 125:200–210
Ambike SS (2011) Characteristics of spatial human arm motion and the kinematic trajectory tracking of similar serial chains. Dissertation, The Ohio State University
Ambike S, Schmiedeler JP (2006) Modeling time invariance in human arm-motion coordination. In: Lenarčič J, Roth B (eds) Advances in robot kinematics. Springer, Dodrecht, pp 177–184
Ambike S, Schmiedeler JP (2013) The leading joint hypothesis for spatial reaching arm motions. Exp Brain Res 224:591–603
Atkeson CG (1989) Learning arm kinematics and dynamics. Ann Rev Neurosci 12:157–183
Atkeson CG, Hollerbach JM (1985) Kinematic features of unrestrained vertical arm movements. J Neurosci 5:2318–2330
Bernstein NA (1967) The coordination and regulation of movement. Pergamon, New York
Breteler MDK, Meulenbroek RGJ, Gielen SCAM (1998) Geometric features of workspace and joint-space paths of 3D reaching movements. Acta Psychol 100:37–53
Conditt MA, Mussa-Ivaldi FA (1999) Central representation of time during motor learning. Proc Natl Acad Sci 96:11625–11630
Conditt MA, Gandolfo F, Mussa-Ivaldi FA (1997) The motor system does not learn the dynamics of the arm by rote memorization of past experience. J Neurophysiol 78:554–560
Dounskaia N (2007) Kinematic invariants during cyclical arm movements. Biol Cybern 96:147–163
Dounskaia N, Wiselder D, Johnson T (2005) Influence of biomechanical factors on substructure of pointing movements. Exp Brain Res 164:505–516
Engström DA, Kelso JAS, Holroyd T (1996) Reaction-anticipation transitions in human perception-action cycles. Human Movement Sci 15:809–832
Feldman AG (1986) Once more for the equilibrium-point hypothesis (lambda model) for motor control. J Mot Behav 18:17–54
Feldman AG (2011) Space and time in the context of equilibrium-point theory. Wiley Cogn Sci 2:287–304
Flanders M, Herrmann U (1992) Two components of muscle activation: scaling with the speed of arm movement. J Neurophysiol 67:931–943
Flash T, Hogan N (1985) The coordination of arm movements: an experimentally confirmed mathematical model. J Neurosci 5:1688–1703
Gentili R, Cahouet V, Papaxanthis C (2007) Motor planning of arm movements is direction-dependent in the gravity field. Neuroscience 145:20–32
Haggard P, Richardson J (1985) Spatial patterns in the control of human arm movement. J Exp Psych 22:42–62
Hollerbach JM, Flash T (1982) Dynamic interactions between limb segments during planar arm movement. Biol Cybern 44:67–77
Isenberg C, Conrad B (1994) Kinematic properties of slow arm movements in Parkinson’s disease. J Neurol 241:323–330
Ivry RB, Hazeltine RE (1994) Perception and production of temporal intervals across a range of durations: evidence for a common timing mechanism. J Exp Psych 21:3–18
Jones T (1968) Statistical analysis of orientation data. J Sediment Petrol 38:61–67
Kawato M (1999) Internal models for motor control and trajectory planning. Curr Opin Neurobiol 9:718–727
Lacquaniti F, Terzuolo C, Viviani P (1983) The law relating the kinematic and figural aspects of drawing movements. Acta Psychol 54:115–130
Liao M, Jagacinski RJ, Greenberg N (1997) Quantifying the performance limitations of older and younger adults in a target acquisition task. J Exp Psych 23:1644–1664
Loeb GE (2012) Optimal isn’t good enough. Biol Cybern 106:757–765
Messier J, Adamovich S, Berkinblit M, Tunik E, Poizner H (2003) Influence of movement speed on accuracy and coordination of reaching movements to memorized targets in three-dimensional space in a deafferented subject. Exp Brain Res 4:399–416
Meyer DE, Abrams RA, Kornblum S, Wright CE (1988) Optimality in human motor performance: ideal control of rapid aimed movements. Psych Rev 95:340–370
Meyer DE, Smih JEK, Kornblum S, Abrams RA, Wrisht CE (1990) Speed-accuracy tradeoffs in aimed movement: toward a theory of rapid voluntary action. In: Jeannerod M (ed) Attention and performance XIII. Lawrence Erlbaum Associates, New Jersey, pp 172–226
Morasso P (1981) Spatial control of arm movements. Exp Brain Res 42:223–227
Nishikawa KC, Murray ST, Flanders M (1999) Do arm postures vary with the speed of reaching? J Neurophysiol 81:2582–2586
Papaxanthis C, Pozzo T, Schieppati M (2003) Trajectories of arm pointing movements on the sagittal plane vary with both direction and speed. Exp Brain Res 148:498–503
Papaxanthis C, Pozzo T, McIntyre J (2005) Kinematic and dynamic processes for the control of pointing movements in humans revealed by short-term exposure to microgravity. Neuroscience 135:371–383
Prilutsky B, Zatsiorsky V (2002) Optimization-based models of muscle coordination. Exerc Sports Sci Rev 30:1–13
Schaal S, Sternad D (2001) Origins and violations of the 2/3 power law in rhythmic three-dimensional arm movements. Exp Brain Res 136:60–72
Soechting JF, Lacquaniti F (1981) Invariant characteristics of a pointing movement in man. J Neurosci 1:710–720
Soechting JF, Lacquaniti F, Terzuolo CA (1986) Coordination of arm movements in three-dimensional space. Sensorimotor mapping during drawing movement. Neuroscience 17:295–311
Soechting JF, Buneo CA, Herrmann U, Flanders M (1995) Moving effortlessly in three dimensions: does Donder’s law apply to arm movement? J Neurosci 15:6271–6280
Soechting JF, Gordon AM, Engel KC (1996) Sequential hand and finger movements: typing and piano playing. In: Bloedel JR, Ebner TJ, Wise SP (eds) Acquisition of motor behavior in vertebrates. MIT Press, Cambridge
Todorov E (2004) Optimality principles in sensorimotor control. Natl Neurosci 7:907–915
Viviani P, Terzuolo C (1982) Trajectory determines movement dynamics. Neuroscience 7:431–437
van der Wel PRD, Sternad D, Rosenbaum DA (2010) Moving the arm at different rates: slow movements are avoided. J Motor Behav 42:29–39
Wolpert DM, Ghahramani Z, Jordan MI (1995) Are arm trajectories planned in kinematic or dynamic coordinates? An adaptation study. Exp Brain Res 103:460–470
Wua G, van der Helm FCT, Veeger HEJ, Makhsouse M et al (2005) ISB recommendation on definitions of joint coordinate systems of various joints for the reporting of human joint motion—part II: shoulder, elbow, wrist and hand. J Biomech 38:981–992
Yadav V (2010) Validation of a time-scaling-based model for representation of dynamics in humans and its applications in rehabilitation. Dissertation, The Ohio State University
Zar JH (1999) Biostatistical analysis. Prentice Hall, New Jersey
Acknowledgments
Support was provided by grant no. IIS-0937612 from the National Science Foundation. The authors wish to thank Mr. Julian Corona for his assistance in data collection, and the reviewers of this paper for their insightful comments.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Ambike, S., Schmiedeler, J.P. Invariant geometric characteristics of spatial arm motion. Exp Brain Res 229, 113–124 (2013). https://doi.org/10.1007/s00221-013-3599-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-013-3599-9