
Abstract
Healthy persons exhibit relatively small temporal and spatial gait variability when walking unimpeded. In contrast, patients with a sensory deficit (e.g., polyneuropathy) show an increased gait variability that depends on speed and is associated with an increased fall risk. The purpose of this study was to investigate the role of vision in gait stabilization by determining the effects of withdrawing visual information (eyes closed) on gait variability at different locomotion speeds. Ten healthy subjects (32.2 ± 7.9 years, 5 women) walked on a treadmill for 5-min periods at their preferred walking speed and at 20, 40, 70, and 80 % of maximal walking speed during the conditions of walking with eyes open (EO) and with eyes closed (EC). The coefficient of variation (CV) and fractal dimension (α) of the fluctuations in stride time, stride length, and base width were computed and analyzed. Withdrawing visual information increased the base width CV for all walking velocities (p < 0.001). The effects of absent visual information on CV and α of stride time and stride length were most pronounced during slow locomotion (p < 0.001) and declined during fast walking speeds. The results indicate that visual feedback control is used to stabilize the medio-lateral (i.e., base width) gait parameters at all speed sections. In contrast, sensory feedback control in the fore-aft direction (i.e., stride time and stride length) depends on speed. Sensory feedback contributes most to fore-aft gait stabilization during slow locomotion, whereas passive biomechanical mechanisms and an automated central pattern generation appear to control fast locomotion.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
A stable walking pattern depends on neuronal spinal and supraspinal pattern generators as well as sensory feedback from visual, vestibular, and proprioceptive systems. The system primarily responsible for dynamic stability in normal, unimpeded walking is the visual system (Warren 2009; Patla 1997; Iosa et al. 2012; Hallemans et al. 2010). When it is absent as in darkness or with eyes closed, walking is always more difficult. Indeed, previous studies showed that absence or perturbation of visual information flow affects multiple aspects of gait kinematics: heading direction, walking speed, cadence, stride length, stance phase duration, swing limb trajectory, foot elevation, foot positioning, and upper body stability (Patla 1997; Assaiante et al. 1989; Cromwell et al. 2002; Hollands and Marple-Horvat 1996; Jahn et al. 2001; Patla and Goodale 1996; Rhea and Rietdyk 2007; Rietdyk and Rhea 2006; Bauby and Kuo 2000; Iosa et al. 2012). However, walking speed itself also affects both visual and vestibular locomotor control. The impact of sensory loss or perturbation on gait decreases as walking speed increases (Jahn et al. 2001; Brandt et al. 1999; Brandt 2000) because the control of locomotion at high speed is more under automated control than under sensory control. Functional imaging has confirmed that the activity in sensory cortex areas is decreased at faster walking speeds (Jahn et al. 2008). Accordingly, patients with unilateral or bilateral vestibular deficits perform better at high walking velocity (Schniepp et al. 2012; Brandt et al. 1999).
The healthy walking pattern is characterized by moderate natural gait variability, that is, stride-to-stride fluctuations of spatiotemporal gait cycle parameters. The structure of these fluctuations is complex, exhibiting long-range correlations, that is, fluctuations at any arbitrary point of time are statistically related to fluctuations at other time points (Hausdorff et al. 1995). The magnitude and intrinsic structure of these gait fluctuations depend on the locomotion speed (Jordan et al. 2007; Hausdorff et al. 1996; Wuehr et al. 2012). Several earlier studies on gait variability showed that these fluctuations contain important information on the regulation process and the interaction between different systems of the locomotion network (Hausdorff 2005). Increased variability is critically linked to a higher risk of falls (Hausdorff et al. 2001; Maki 1997). Recent studies reported a tight connection between deficient sensory locomotion control and alterations of gait variability. This was observed in patients with chronic vestibular failure who show pathologically increased gait variability during slow locomotion; it decreases to a normal level when walking velocity increases (Schniepp et al. 2012). Furthermore, it has been shown that the absence of visual information leads to increased levels of medio-lateral gait variability (Bauby and Kuo 2000).
The purpose of this study was to systematically investigate how the absence of visual locomotion control influences gait variability at different locomotion speeds. We therefore examined the alterations in the magnitude (i.e., the coefficient of variation) and structure (i.e., the fractal dimension) of stride-to-stride fluctuations in different medio-lateral and fore-aft gait parameters during the conditions of walking with eyes open and with eyes closed. We hypothesized that the absence of visual information might have a similar speed-dependent impact on walking variability as was shown previously for deficient vestibular locomotion control (Schniepp et al. 2012). This would suggest a general principle of sensory locomotor control.
Methods
Subjects and procedures
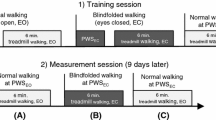
Ten healthy subjects (32.2 ± 7.9 years, 5 women) participated in the study (Table 1). The walking pattern of each subject was measured under normal walking conditions with eyes open (EO) and with eyes closed (EC). While walking with eyes closed, subjects wore light-impenetrable eyeglasses. Gait was analyzed with two different systems to capture both normal over-ground locomotion on a carpet system and steady-state locomotion on a treadmill system. For both conditions (EO and EC), normal over-ground locomotion was recorded with a pressure-sensitive carpet system (GAITRite®, CIR System, Havertown, USA) 6.7-m long with a sampling rate of 120 Hz. Steady-state locomotion was measured on a pressure-sensitive treadmill system (Zebris®, Isny, Germany; h/p/cosmos®, Nussdorf-Traunstein, Germany) 1.6-m long with a sampling rate of 100 Hz. The individual preferred walking velocity (PWS) and maximal walking velocity (MWS) were determined during over-ground locomotion on the carpet system. Each walk was started 1.5 m in front of the sensor mat and continued for 1.5 m beyond it in order to provide uninterrupted locomotion in the sensor area. Subsequently, five different walking velocities (20, 40, 66, 70, 80 % of MWS and PWS) were recorded on the treadmill system in a randomized order, each for a period of 5 min. Before the recording, participants were given 1 min to acclimatize to the preset treadmill speed. Between trials, participants had at least 2 min to recover. Treadmill data for the right and left legs were later pooled, since inter-limb correlations showed no significant difference in the median of both limbs for all subjects and both walking conditions (Wilcoxon-test and Mann–Whitney U test, p > 0.05).
All subjects gave their written informed consent prior to the experiments. The study protocol was approved by the local Ethics Committee. The study was conducted in conformity with the Declaration of Helsinki.
Data analysis
Walking velocity
The walking velocities measured on the carpet system (PWS and MWS as well as the ratio PWS/MWS) were compared for both walking conditions to determine whether the absence of visual information has an impact on maximal and preferred walking speeds.
Three different gait cycle variables of the treadmill recordings were examined: stride time, stride length, and base width. Stride time was defined as the time elapsed between the first sensor contacts of two consecutive footfalls of the same foot. Stride length was defined as the distance on a line of progression between the heel points of two consecutive footprints of the same foot. Base width was defined as the vertical distance from the heel center of one foot to the line of progression formed by two consecutive footprints of the opposite foot.
Coefficient of variation
The coefficient of variation (CV) was calculated for the time series of each dependent gait cycle variable. The CV is defined as the standard deviation σ divided by the mean μ:
Fractal dimension
The fractal dimension provides a measure for the structure of the variability of a time series and gives a quantitative measure for the strength of long-range correlations within the time series. Stronger correlations indicate a more predictable time series, whereas weaker correlations indicate a less predictable time series. The fractal dimension of the stride-to-stride fluctuations was calculated by a type of root mean square analysis, the detrended fluctuation analysis (DFA) (Hausdorff et al. 1995). This method first forms an accumulated sum of the time series with N samples and divides it further into boxes ranging from 4 to N/4 data points. Finally, the log of the average size of fluctuation for a given box size is plotted against the log of the actual box size. This computation is repeated over all box sizes. The slope of the line relating the log of the average box size to the log of the average fluctuation size, α, gives a quantitative measure for the strength of long-range correlations within the time series. An α value of 0.5 or less indicates a white noise process, whereas α values between 0.5 and 1.0 indicate persistent long-range correlations.
Statistical analysis
The CV and the fractal dimension were calculated for each trial and each analyzed gait cycle parameter. Afterward, nonlinear regression analysis was performed on the CV and DFA results to test for the presence of a curvilinear dependency on walking speed. The effects of each dependent variable were examined by a two-way repeated measures ANOVA with walking condition and velocity as factors. Post hoc analysis was carried out using a Bonferroni post hoc test. Results are considered significant if p < 0.05. Statistical analysis was performed using SPSS (Version 20).
Results
The results of the comparison of walking speed during normal walking and walking with eyes closed are presented in Table 1. Results of the two-way repeated measures ANOVA are given in Table 2.
Walking velocity
Under absence of visual information, subjects walked at a slower PWS (EC 120.4 ± 23.6 cm/s vs. EO 138.0 ± 23.4 cm/s; n.s.) as well as at a significantly slower MWS (EC 182.9 ± 22.2 cm/s vs. EO 208.5 ± 25.2 cm/s; p < 0.027) compared to normal walking conditions. However, the ratio of preferred to maximal walking velocity (PWS/MWS) remained unchanged (~66 %) (Table 1).
Variability magnitude
Significant speed effects on stride time CV as well as stride length CV were observed under the condition EO (20 % vs. all other walking speeds; p < 0.001) and the condition EC (20 % vs. all other walking speeds; p < 0.001). Moreover, base width CV showed significant speed effects for both conditions: EO (20 % vs. all other walking speeds; p = 0.002) as well as EC (20 % vs. all other walking speeds; p = 0.004). CV values of stride time, stride length, and base width were significantly higher under EC conditions for all walking speeds (p = 0.002) (Table 2).
Variability structure
Under EO conditions, no significant differences of α were found for stride time or stride length. Under the condition of EC, α of stride time (40 vs. 80 %; p = 0.003) and α of stride length (20 vs. 80 %; p = 0.035, 40 % vs. 70 and 80 %; p = 0.001) were significantly different. Base width α showed significant speed effects for the condition EO (20 % vs. all other walking speeds; p = 0.005) and EC (20 vs. 63 %; p = 0.007 and 20 vs. 80 %; p = 0.011). Stride time and stride length α were significantly lower for the condition EC at 20 and 40 % of MWS compared to EO (p < 0.001). The fractal dimension of base width was significantly higher with EC at 20 % of MWS than with EO (p = 0.010) (Table 2).
Regression analysis
Second-order polynomials were fit to the CV results (Fig. 1) and the fractal dimension results (Fig. 2). Under the condition EO, adjusted r-squared values of 84, 87, 32, 21, and 24 % were found for stride time CV, stride length CV, stride time α, stride length α, and base width α. Under the condition EC, adjusted r-squared values of 80, 87, 50, and 39 % were observed for stride time CV, stride length CV, stride length α, and base width α. For all variables except for base width CV (EO and EC) and stride time α (EC), significant quadratic components were found, but r-squared values for linear fits were smaller than that for quadratic fits in all of these cases.

Speed dependency of variability magnitude. Dependency of the coefficient of variation (CV) of stride time (a), stride length (b), and base width (c) on the walking speed. Black diamonds represent mean CV values for walking with eyes open (EO) at 20, 40, 70, 80 % of maximal walking speed and preferred walking speed (PWS; major black diamond). White circles represent mean CV values for walking with eyes closed (EC) at 20, 40, 70, 80 % of maximal walking speed and preferred walking speed (PWS; major white circle). A polynomial function was calculated for each gait cycle parameter (black line for the EO condition, dotted line for the EC condition). *Significant difference of the mean CV value between the conditions at the same walking speed

Speed dependency of variability structure. Dependency of long-range correlations (α) of stride time (a), stride length (b), and base width (c) on the walking speed. Black diamonds represent mean α values for walking with eyes open (EO) at 20, 40, 70, 80% of maximal walking speed and preferred walking speed (PWS; major black diamond). White circles represent mean α values for walking with eyes closed (EC) at 20, 40, 70, 80% of maximal walking speed and preferred walking speed (PWS; major white circle). A polynomial function was calculated for each gait cycle parameter (black line for the EO condition, dotted line for the EC condition). *Significant difference of the mean α value between the conditions at the same walking speed
Discussion
The present study examined the influence of an absent visual locomotion control on the variability in the walking pattern during different locomotion speeds. We analyzed the variability magnitude (CV) and structure (fractal dimension) of three standard temporospatial gait cycle parameters during walking with eyes open and with eyes closed. The results show that withdrawing visual information has a direct effect on the stride-to-stride fluctuations. These fluctuations are differentially regulated for the medio-lateral and the fore-aft walking planes and are also dependent on the walking speed.
Dependency of gait variability on visual locomotion control
The healthy human walking pattern is characterized by highly complex dynamics that result from the interaction of active sensory and passive biomechanical regulatory processes operating on multiple time scales. Consequently, temporospatial gait cycle parameters not only exhibit a stride-to-stride variability, but also comprise complex fractal-like long-range correlations (Hausdorff 2005; Hausdorff et al. 1996; Terrier et al. 2005). This inherent complexity in the healthy walking pattern allows for adaptive responses, making possible fast reactions to sudden changes or obstacles in the environment and so ensuring dynamic stability of the walking subject. Perturbations of the walking condition such as the absence of sensory feedback or shifts of the walking speed to increasingly non-preferred velocities are thought to switch the inherent walking dynamics to a less complex response mode of reactive tuning, which operates over relatively short time periods to restore the stability of the walking subject (Lipsitz 2002; Lauk et al. 1998).
Medio-lateral gait variability
The present data show that the stride-to-stride fluctuations in the walking pattern during normal unperturbed walking differ in dynamics for the fore-aft plane (i.e., stride time and stride length) compared to the medio-lateral plane (i.e., base width). Apparently different control strategies underlie each of these directions. Base width fluctuations showed large variability magnitudes that even increased for faster walking speeds due to a decrease of the base of support with faster walking modes. The variability structure of base width fluctuations showed low fractal dimensionality close to a random-like white noise process. Walking adjustments in the medio-lateral direction are believed to be actively controlled by integrative sensory feedback because the passive dynamics of walking appear to be quite unstable in this direction (Bauby and Kuo 2000; O’Connor and Kuo 2009). The observed large variability magnitudes and the absence of long-range correlations within the stride-to-stride fluctuations of base width confirm this hypothesis. Our findings indicate that the stabilization of the medio-lateral walking plane is governed by the active control mode of reactive tuning that acts on the short-term time scale thereby evoking single-frequency responses to restore the stability of the walking subject (Lipsitz 2002). Moreover, the active stabilization in the medio-lateral plane, which is based on active sensory feedback control, should be highly sensitive to perturbation or absence of visual feedback. Again our results agree with previously published data, showing that the absence of visual locomotion control leads to a general increase in the magnitude of medio-lateral walking variability at all walking speeds (Bauby and Kuo 2000).
Fore-aft gait variability
During unperturbed walking with eyes open, the variability magnitude of stride time and stride length was considerably smaller than that of the medio-lateral fluctuations in base width. The CV of stride time and stride length decreased as walking speed increased, suggesting that the gait cycle in the fore-aft plane becomes more consistent when the walking speed increases (Winter 1983). Fractal-like long-range correlations were present in the stride-to-stride fluctuations of stride time and stride length indicating the presence of long-term passive control mechanisms that govern the stabilization of the fore-aft walking plane (Lipsitz 2002). Moreover, the variability structure of the fore-aft parameter fluctuations showed a U-shaped pattern of change with increasing speed. Thereby lowest fractal dimensionality is associated with preferred walking velocity, a finding that agrees with previously published data (Jordan et al. 2007; Wuehr et al. 2012).
It has been proposed that the stabilization of fore-aft locomotion is established by passive biomechanical regulatory processes and should be essentially independent of high-level neural feedback control (Mcgeer 1990; Gates et al. 2007; Bauby and Kuo 2000). Consequently, the regulation of fore-aft gait variability should be rather insensitive to perturbations in the active sensory feedback control. However, our present data show that the absence of visual feedback control leads to a significant increase in magnitude of stride time and stride length variability and a significant decrease in the fractal dimensionality of the fluctuations in these gait cycle parameters, thereby indicating an increased involvement of active control mechanism that stabilizes gait. This effect differs considerably for different walking speeds; while major changes occur at slow locomotion, the impact diminishes at preferred and fast locomotion speeds. This speed-dependent impact of visual information on the fore-aft gait variability supports the hypothesis of a speed-dependent sensory locomotion control in the fore-aft walking direction. Accordingly, active sensory feedback control is necessary for balance control mainly during slow locomotion, whereas fast locomotion is achieved by highly automated central pattern generators in the spinal cord (Brandt et al. 1999; Brandt 2000; Jahn et al. 2001).
Correspondingly to this model, motor imagery studies in fMRI have shown that activations of sensory cortex areas decrease during running and fast walking (Jahn et al. 2008). Interestingly, the speed-dependent effect of an acute absence of visual information on the fore-aft gait variability reported here is akin to the previously reported speed-dependent impact of a chronic bilateral vestibular failure on the magnitude of stride time variability (Schniepp et al. 2012). This supports the hypothesis that active feedback control of both sensory systems might contribute to the stabilization of fore-aft locomotion.
Gait variability at preferred walking speed
In normal unconstrained walking, the minimal gait variability, lowest energy expenditure, and symmetry of ground reaction forces occur at self-paced walking velocity (Jordan et al. 2007; Zarrugh et al. 1974; Breit and Whalen 1997). This pattern of optimization might be due to the spontaneous interaction of neural and biomechanical oscillations, with only minimal active control of high-level sensory feedback control. Previous studies could demonstrate that neither disturbed cerebellar locomotor function (Wuehr et al. 2012) nor disturbed vestibular feedback control (Schniepp et al. 2012) affect the occurrence of minimal fore-aft gait variability at preferred walking velocity, thereby supporting the view that passive tuning mechanisms of gait velocity and body dynamics can compensate for impaired supraspinal locomotion control. In the present study, we could observe that the absence of visual feedback also has only minimal or no effect on the fore-aft gait variability during preferred walking speed. Therewith this study provides further evidence that the pattern of optimization of fore-aft gait variability at self-paced walking might originate from passive biomechanical tuning mechanism and would be mainly independent of active sensory feedback control. In contrast, medio-lateral gait variability did not follow the same pattern of optimization during walking with eyes open or with eyes closed, a finding in line with the hypothesis that active control dominates over passive control of gait in the medio-lateral plane.
One limiting factor of the present study is the short recording time of 5 min per trial. The DFA algorithm has been shown to require time series data of at least 8 min recording time to produce reliable results for fractal dimension (Pierrynowski et al. 2005). Although the extensive study protocol of testing two different walking conditions, each for five different walking speeds, did not allow data collection for longer time periods, the DFA results are consistent with previously published data (Jordan et al. 2007; Wuehr et al. 2012). Besides, it has been shown that treadmill walking can modify long-range correlations in the gait pattern (Terrier and Deriaz 2011). However, variability magnitude results were comparable between over-ground walking and treadmill walking (data not shown). For the analysis of variability structure, we were restricted to treadmill walking recordings and could therefore not compare these results versus over-ground walking.
Conclusions
We showed that withdrawing visual information has a direct effect on the magnitude and structure of stride-to-stride fluctuations within the walking pattern. Absence of visual information leads to a general increase of base width variability, indicating that gait stabilization in the medio-lateral plane depends on active sensory feedback control. In contrast, withdrawing visual information leads to an increase of fore-aft variability during slow locomotion but not during faster walking speeds. This finding supports the hypothesis that sensory feedback control for the stabilization of fore-aft locomotion is speed-dependent. Absence of visual feedback control does not affect the pattern of optimization of gait variability in self-paced walking (observed during unconstrained walking). Thus, passive tuning of walking velocity and body dynamics can compensate for deficiencies in active sensory feedback control.
References
Assaiante C, Marchand AR, Amblard B (1989) Discrete visual samples may control locomotor equilibrium and foot positioning in man. J Mot Behav 21(1):72–91
Bauby CE, Kuo AD (2000) Active control of lateral balance in human walking. J Biomech 33(11):1433–1440
Brandt T (2000) Vestibulopathic gait: you’re better off running than walking. Curr Opin Neurol 13(1):3–5
Brandt T, Strupp M, Benson J (1999) You are better off running than walking with acute vestibulopathy. Lancet 354(9180):746. doi:10.1016/S0140-6736(99)03179-7
Breit GA, Whalen RT (1997) Prediction of human gait parameters from temporal measures of foot-ground contact. Med Sci Sports Exerc 29(4):540–547
Cromwell RL, Newton RA, Forrest G (2002) Influence of vision on head stabilization strategies in older adults during walking. J Gerontol A Biol Sci Med Sci 57(7):M442–M448
Gates DH, Su JL, Dingwell JB (2007) Possible biomechanical origins of the long-range correlations in stride intervals of walking. Phys A 380:259–270. doi:10.1016/j.physa.2007.02.061
Hallemans A, Ortibus E, Meire F, Aerts P (2010) Low vision affects dynamic stability of gait. Gait Posture 32(4):547–551. doi:10.1016/j.gaitpost.2010.07.018
Hausdorff JM (2005) Gait variability: methods, modeling and meaning. J Neuroeng Rehabil 2:19. doi:10.1186/1743-0003-2-19
Hausdorff JM, Peng CK, Ladin Z, Wei JY, Goldberger AL (1995) Is walking a random walk? Evidence for long-range correlations in stride interval of human gait. J Appl Physiol 78(1):349–358
Hausdorff JM, Purdon PL, Peng CK, Ladin Z, Wei JY, Goldberger AL (1996) Fractal dynamics of human gait: stability of long-range correlations in stride interval fluctuations. J Appl Physiol 80(5):1448–1457
Hausdorff JM, Rios DA, Edelberg HK (2001) Gait variability and fall risk in community-living older adults: a 1-year prospective study. Arch Phys Med Rehabil 82(8):1050–1056. doi:10.1053/apmr.2001.24893
Hollands MA, Marple-Horvat DE (1996) Visually guided stepping under conditions of step cycle-related denial of visual information. Exp Brain Res 109(2):343–356
Iosa M, Fusco A, Morone G, Paolucci S (2012) Effects of visual deprivation on gait dynamic stability. ScientificWorldJournal 2012:974560. doi:10.1100/2012/974560
Jahn K, Strupp M, Schneider E, Dieterich M, Brandt T (2001) Visually induced gait deviations during different locomotion speeds. Exp Brain Res 141(3):370–374. doi:10.1007/s002210100884
Jahn K, Deutschlander A, Stephan T, Kalla R, Wiesmann M, Strupp M, Brandt T (2008) Imaging human supraspinal locomotor centers in brainstem and cerebellum. Neuroimage 39(2):786–792. doi:10.1016/j.neuroimage.2007.09.047
Jordan K, Challis JH, Newell KM (2007) Walking speed influences on gait cycle variability. Gait Posture 26(1):128–134. doi:10.1016/j.gaitpost.2006.08.010
Lauk M, Chow CC, Pavlik AE, Collins JJ (1998) Human balance out of equilibrium: nonequilibrium statistical mechanics in posture control. Phys Rev Lett 80(2):413–416
Lipsitz LA (2002) Dynamics of stability: the physiologic basis of functional health and frailty. J Gerontol A Biol Sci Med Sci 57(3):B115–B125
Maki BE (1997) Gait changes in older adults: predictors of falls or indicators of fear. J Am Geriatr Soc 45(3):313–320
Mcgeer T (1990) Passive dynamic walking. Int J Robot Res 9(2):62–82
O’Connor SM, Kuo AD (2009) Direction-dependent control of balance during walking and standing. J Neurophysiol 102(3):1411–1419. doi:10.1152/jn.00131.2009
Patla AE (1997) Understanding the roles of vision in the control of human locomotion. Gait Posture 5(1):54–69. doi:10.1016/s0966-6362(96)01109-5
Patla AE, Goodale MA (1996) Obstacle avoidance during locomotion is unaffected in a patient with visual form agnosia. NeuroReport 8(1):165–168
Pierrynowski MR, Gross A, Miles M, Galea V, McLaughlin L, McPhee C (2005) Reliability of the long-range power-law correlations obtained from the bilateral stride intervals in asymptomatic volunteers whilst treadmill walking. Gait Posture 22(1):46–50. doi:10.1016/j.gaitpost.2004.06.007
Rhea CK, Rietdyk S (2007) Visual exteroceptive information provided during obstacle crossing did not modify the lower limb trajectory. Neurosci Lett 418(1):60–65. doi:10.1016/j.neulet.2007.02.063
Rietdyk S, Rhea CK (2006) Control of adaptive locomotion: effect of visual obstruction and visual cues in the environment. Exp Brain Res 169(2):272–278. doi:10.1007/s00221-005-0345-y
Schniepp R, Wuehr M, Neuhaeusser M, Kamenova M, Dimitriadis K, Klopstock T, Strupp M, Brandt T, Jahn K (2012) Locomotion speed determines gait variability in cerebellar ataxia and vestibular failure. Mov Disord 27(1):125–131. doi:10.1002/mds.23978
Terrier P, Deriaz O (2011) Kinematic variability, fractal dynamics and local dynamic stability of treadmill walking. J Neuroeng Rehabil 8:12. doi:10.1186/1743-0003-8-12
Terrier P, Turner V, Schutz Y (2005) GPS analysis of human locomotion: further evidence for long-range correlations in stride-to-stride fluctuations of gait parameters. Hum Mov Sci 24(1):97–115. doi:10.1016/j.humov.2005.03.002
Warren WH (2009) How do animals get about by vision? Visually controlled locomotion and orientation after 50 years. Br J Psychol 100(Pt 1A):277–281. doi:10.1348/000712609X414150
Winter DA (1983) Biomechanical motor patterns in normal walking. J Mot Behav 15(4):302–330
Wuehr M, Schniepp R, Ilmberger J, Brandt T, Jahn K (2012) Speed-dependent temporospatial gait variability and long-range correlations in cerebellar ataxia. Gait Posture. doi:10.1016/j.gaitpost.2012.07.003
Zarrugh MY, Todd FN, Ralston HJ (1974) Optimization of energy expenditure during level walking. Eur J Appl Physiol Occup Physiol 33(4):293–306
Acknowledgments
The authors thank Judy Benson for copyediting the article. The work was supported by the German Research Foundation (Deutsche Forschungsgemeinschaft, DFG JA1087/1-1) and the German Hertie Foundation and the Federal Ministry for Education and Science (BMBF 01EO0901) of Germany.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Wuehr, M., Schniepp, R., Pradhan, C. et al. Differential effects of absent visual feedback control on gait variability during different locomotion speeds. Exp Brain Res 224, 287–294 (2013). https://doi.org/10.1007/s00221-012-3310-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-012-3310-6