
Abstract
The reduction in saccade latency when the fixation point is removed (fixation offset effect–FOE) reflects the degree to which fixation neurons are under influence by a stimulus at fixation. Strategic manipulations of oculomotor readiness that bring these neurons under endogenous control reduce the magnitude of the FOE. Using an aging foreperiod paradigm, and the FOE as a marker for cortical control of reflexive fixation, we showed that, for both prosaccades and antisaccades, increasing preparation across the foreperiod reduced both saccade latency and the FOE. Consistent with Los’s trace conditioning account, these effects reflected greater preparation for trials when the current short foreperiod was preceded by a trial with a short foreperiod. The FOE was also smaller for antisaccades than for prosaccades demonstrating strategic modulation. However, the effects of trace conditioning were comparable in the two tasks, demonstrating that strategic and unconscious priming effects both independently modulate the control of ocular fixation.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Easton argued that the neural circuits that subserve reflexes are the building blocks for more complex behavior; and that the nervous system routinely goes about its business through an orchestration of those circuits by cortical processes that activate or inhibit them (Easton 1972). The evolution of more complex behaviour required cortico-subcoritcal integration to regulate reflexes in the service of goal directed action (Ingle 1973; Rozin 1976). Eye movements provide an attractive model to study how preparatory states influence reflexive behavior. The current investigation employs the fixation offset effect (FOE) as a marker task for probing the effects of strategic control and more automatic, non-specific preparation on oculomotor reflexes.
The fixation offset effect
In the rostral pole of each superior colliculus (SC) are cells that are active during fixation, even in the dark, and whose activity is further increased by a visual signal at fixation (Munoz and Wurtz 1992, 1993b). These fixation neurons help anchor the eyes at fixation. Caudal to the fixation neurons, and inhibited (either directly or indirectly) by them, are neurons (movement cells) whose activity moves the eyes to a new position (Munoz and Istvan 1998). Eye movements toward a peripheral target, then, are controlled by an opponent process: there is mutual inhibition between the visual grasp reflex (VGR), activated by abrupt signals in the visual periphery and mediated by movement neurons, and the fixation reflex, activated by visual signals at fixation and mediated by fixation neurons. Together, the activity of these two types of cells determines when and where the eyes will move (Findlay and Walker 1999; Munoz and Fecteau 2002).
The offset of a fixated stimulus prior to, or simultaneous with, the onset of a peripheral target disinhibits the VGR and reduces the latency to initiate an eye movement to the target (Saslow 1967). This benefit of fixation offset on saccadic latencies has been termed the FOE (Klein and Kingstone 1993), and has been shown to reflect neural activity within the colliculus. When a fixated stimulus offsets, the activity of fixation neurons decreases, and inhibition of movement cells is reduced resulting in shorter saccade latency (Dorris and Munoz 1995; Munoz and Wurtz 1992). Conversely, electrical stimulation of fixation neurons just prior to or during an eye movement can delay or arrest the eye movement (Munoz and Wurtz 1993a). When fixation cell activity decreases in response to fixation offset, movement cells are disinhibited and more quickly reach threshold for saccade initiation thereby reducing saccade latency. The difference in saccade latency between fixation offset and fixation overlap conditions (the FOE) is a measure of the degree to which fixation neurons are under external control by the fixation stimulus.
Strategic control of ocular fixation during the visual grasp reflex
The SC receives visual input both directly from the retina and from visual cortex, as well as input from oculomotor cortex of frontal and parietal lobes. Competition between the demands of voluntary and reflexive eye movement signals are resolved in the SC by their interacting influences on fixation and movement neurons. The emerging evidence indicates that the opponent interactions between collicular fixation and movement neurons are not sequestered in a simple intra-collicular circuit. There seems to be no direct inhibitory connections between the rostral pole fixation neurons and the movement neurons of the SC (Isa 2002; Lee and Hall 2006). In addition, direct retinotectal input to fixation neurons in the rostral pole of the colliculus is not necessary for an FOE to occur (Sumner et al. 2006), suggesting that visual cortex is part of the circuitry of the FOE. Lesions of the human pulvinar abolish the FOE (Rafal et al. 2004), indicating that it, too, is part of the circuitry.
Thus, although the FOE reflects a reflexive response of fixation neurons, the circuitry for this reflex appears to involve other cortical and subcortical structures; and this reflex circuit can be modulated by cognitive control. Manipulations of strategic set can bring the fixation neurons under endogenous control, reducing the influence of the external stimulus at fixation and, thereby, the size of the FOE. The logic here is straightforward. If endogenous control is exercised over fixation neurons before a target appears—either by tonically increasing their activity (for example to prevent errors in an antisaccade task), or by reducing their activity because of an increased readiness to make an eye movement—then the fixation point will have less effect on these neurons, and the FOE will be reduced. It should be noted that any endogenous control of fixation neurons, regardless of whether the exercise of this control increases or decreases fixation neuron activity, may render them less responsive to the exogenous influence of a fixation stimulus and reduce the FOE. The size of the FOE then is not an index of fixation cell activity but, rather, an index of the degree to which they are susceptible to reflexive activation.
In previous experiments manipulating oculomotor set, we have shown that normal adults can modulate the magnitude of the FOE. For example, the FOE decreased when oculomotor readiness was increased by reducing the proportion of catch trials (i.e. in which no target was presented and no eye movement was made) (Machado and Rafal 2000b). In another study the subjects received either informative cues so they could prepare for an upcoming endogenous saccade, or uninformative cues (Machado and Rafal 2004; Rafal et al. 2000). Preparation of a voluntary saccade, prior to the appearance of a peripheral saccade target, not only reduced saccade latencies, but also reduced the FOE. These observations suggest that increasing oculomotor readiness is normally associated with reduced influence of the fixation stimulus on the fixation reflex.
The FOE is also smaller for anti-saccades than for pro-saccades (Forbes and Klein 1996; Machado and Rafal 2000a; Reuter-Lorenz et al. 1991). To prevent errors in a block of anti-saccades, the VGR needs to be suppressed, which is achieved by adopting a strategic oculomotor set that increases fixation cell activity (Everling et al. 1999). This strategic manipulation causes longer saccade reaction times (RT), since more movement cell activity is necessary to reach the saccade threshold. However, since the fixation cells are endogenously activated by the strategic set required in the anti-saccade task, they are less influenced by the removal of an external visual fixation point. This results in a smaller difference in RT between overlap and offset trial, i.e. a reduced FOE.
Priming effects in the aging foreperiod paradigm
The research summarized above demonstrates that strategic preparation can regulate and modulate the circuitry of oculomotor reflexes. However, it is unknown whether more automatic and unconscious cognitive processes, which occur without intention, can influence these reflexes. Here our focus is on non-specific response readiness over time. The effect of non-specific preparation on RT has been studied by varying the foreperiod, which is the time between the onset of a neutral warning stimulus and the onset of the target stimulus, on a trial by trial basis. In experiments with a variable foreperiod duration, a longer foreperiod is associated with a reduction in RT. This is referred to as the variable foreperiod effect, and it is thought to be the result of a higher non-specific preparation for the longer foreperiod (Niemi and Näätänen 1981). In addition to the variable foreperiod effect, there are also sequentional effects; the reaction times on a trial are influenced by the length of the foreperiod of the preceding trial. Sequential effects are asymmetric, i.e. the effect of the preceding trial depends on the length of the foreperiod of the current trial. The RT for long foreperiods are generally not affected by the foreperiod of the previous trial, but the RT for short foreperiods are shorter if the foreperiod of the previous trial is short, compared to when the previous foreperiod is long (Niemi and Näätänen 1981).
It has recently been proposed that preparatory effects across the foreperiod result from priming in which the length of the foreperiod of the previous trial influences the RT of the next trial (Los 1996; Los and Heslenfeld 2005; Los and van den Heuvel 2001). According to their trace conditioning model, the RT associated with a foreperiod depends on the conditioning strength that is associated with it. The way in which the foreperiod of the previous trial influences the conditioning strength for specific foreperiods on the current trial follows a set of rules. Firstly, the conditioning strength corresponding to a certain foreperiod is reinforced if the stimulus is presented at that foreperiod. Secondly, it is suppressed if the stimulus is presented at a later foreperiod. Thirdly, there is no change if the stimulus is presented at a shorter foreperiod. According to these rules, priming by the previous trials is only present for events with short foreperiods, because on the previous trial these conditioning strength are either reinforced (foreperiod previous trial is short) or suppressed (foreperiod previous trial is long). Importantly, because the conditioning strength of the long foreperiod is never suppressed, i.e. it is not bypassed in time, it approaches some asymptotic value in the course of a few trials, resulting in no priming effect of the foreperiod of the previous trial for the long foreperiod. Los and Heslenfeld (2005) provided empirical evidence for this model in an experiment in which they presented the stimulus after either a short (400 ms) or a long foreperiod (1,200 ms). In addition, they demonstrated that subjects had no intentional control over this process. An attraction of Los’ model is its parsimony as a ‘single process’ account: trace conditioning accounts for both the sequential effect and for its asymmetry. Another, dual-process, account has been proposed to explain the asymmetric sequential effects (Vallesi and Shallice 2007). This too construe sequential effects as being due to automatic priming, but argues that the asymmetry of the sequential effect results from strategic preparation supervening on trials with a long foreperiod, thus abolishing the effects of priming on these trials.
In two experiments, the present study investigated the effect of non-specific preparation on the cognitive control of oculomotor reflexes by measuring the size of the FOE, while manipulating the amount of non-specific preparation by systematically varying sequences of the foreperiod of the previous trial and current trial. In addition, we manipulated the strategic set using a pro-saccade task in experiment 1 and an anti-saccade task in experiment 2 to determine if non-specific preparation and strategic preparation are independent processes.
Experiment 1
Methods
Participants
Fifteen undergraduate psychology students at the University of Wales, Bangor participated for course credits.
Stimuli and procedure
Horizontal eye position was recorded with an Eye Trac 210 scleral reflectance device (ASL) at a sampling rate of 1,000 Hz. A 50 deg/s velocity criterion was used to compute the latency of saccade onset. Presentation software (Neurobehavioral Systems) was used for stimulus presentation, and recording of saccade RT.
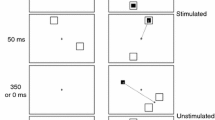
Throughout the experiment, two white marker boxes (1.5°) on a black background were presented at 9° to the left and right of the centre of the screen. After an inter-trial interval of 2,500 ms, each trial began with the onset of a Fixation point, a 0.4° white circle, in the centre of the screen. After either 500, or 1,500 ms, the left (50%), or right (50%) marker box turned white. Participants were instructed to make an eye movement to the centre of this box as fast as possible. On half of the trials, the fixation point remained visible (overlap condition), while on the other half it disappeared simultaneously with the onset of the visual target (offset condition). The target remained on the screen until subjects made a response. A total of 384 trials were presented in six blocks. An algorithm was used to ensure that each previous foreperiod–current forepriod combination had an equal probability within each block of trials. The algorithm randomized the sequence, and checked whether all combinations had an equal probability in each block. The randomized sequence was only used when all the combinations had an equal probability, else the randomization procedure was repeated.
Since the onset of the fixation point signaled the start of the trial, and it is crucial that subjects are aware of the start of the trial in a foreperiod paradigm, subjects were instructed to look at the centre of the screen at the start of the trial, and keep their eyes at the centre throughout the whole foreperiod. This was monitored online, and if the subject failed to do this, the trial was ended and an error sound was presented for 100 ms. An error sound was also presented if subjects blinked during the foreperiod, moved their eyes in the wrong direction, or responded too fast (<50 ms), or too slow (>800 ms).
Results and discussion
Errors
An analysis of variance (ANOVA) with the foreperiod of the current trial [Foreperiod(current)], the foreperiod of the previous trial [Foreperiod(previous)], and Fixation point condition as factors, showed a significant main effect of Foreperiod(current), F(1,14) = 6.81, P < 0.05, η 2 p = 0.33, indicating that subjects made more errors if the current trial had a long foreperiod (5.3%) compared to if the current trial had a short foreperiod (3.6%). In addition, there was a significant main effect of Foreperiod(previous), F(1,14) = 4.96, P < 0.05, η 2 p = 0.26, reflecting that less errors were made in trials that were preceded by a short foreperiod (4.0%) than in trials that were preceded by a long foreperiod (5%). No other effects were found to be significant (see Table 1).
Saccadic RT
Trials on which an error was made were excluded from the Saccadic RT analyses. Since a preliminary analyses showed no difference in saccade RT for left and right eye movements (P > 0.1) data for left and right eye movements were pooled. Mean saccade latency for correct responses were calculated in each condition for each participant and subjected to a repeated measures ANOVA with the foreperiod of the current trial [Foreperiod(current)], the foreperiod of the previous trial [Foreperiod(previous)], and Fixation point condition (offset and overlap) as factors. Figure 1 (top panel) depicts the mean saccadic RT of Experiment 1. Saccade RT was shorter (FOE) for trials on which the fixation point offset (283 ms) than when it overlapped (331 ms) with the target [F(1,14) = 35.88, P < 0.01, η 2 p = 0.72]. Saccade RT was shorter for trials with a long (295 ms) than for those with a short foreperiod (319 ms), F(1,14) = 20.11, P < 0.01, η 2 p = 0.59, demonstrating that the variable foreperiod effect (Niemi and Näätänen 1981) is manifest for saccadic responses. As expected, the effect of the preceding trial was asymmetrical and, as shown in Fig. 1, was only present for trials on which the current foreperiod was short. This was confirmed by a highly significant interaction between Foreperiod(current) × Foreperiod(previous), F(1,14) = 55.02, P < 0.01, η 2 p = 0.78. Pair wise comparisons (Bonferroni corrected) revealed that saccade RT was longer (P < 0.001) if a short foreperiod was preceded by a long foreperiod on the previous trial (334 ms) compared to all other Foreperiod(previous) × Foreperiod(current) conditions (short–short 305 ms; long–long 296 ms; short–long 293 ms). None of the other conditions differed significantly from each other.

Mean saccadic reaction times for pro-saccades (top panel) and anti-saccades (bottom panel) as a function of the foreperiod on the current trial, the foreperiod on the preceding trial, and fixation condition. The size of the fixation offset effect (FOE) is displayed above each bar
The key finding of the experiment was that the FOE was modulated by the preparatory state over the foreperiod, and that this modulation also reflected the asymmetric influence of priming on preparatory state by the preceding trial. As shown in Fig. 1, the FOE was larger on trials with a short foreperiod when preceded by a trial with a long foreperiod: Foreperiod(current) × Foreperiod(previous) × Fixation Point Condition, [F(1,14) = 5.04, P = 0.04, η 2 p = 0.27]. Six pair wise comparisons of the magnitude of the FOE for each Foreperiod(current) × Foreperiod(previous) condition (Bonferroni corrected) were conducted. The FOE was significantly smaller in the Foreperiod(current=short) − Foreperiod(previous=short) condition (40 ms) than the Foreperiod(current=short) − Foreperiod(previous=long) condition (56 ms), t(14) = 3.94, P < 0.01. None of the other pair wise comparisons were significant.
The results revealed that when the conditioning strength of the current trial was reinforced on the previous trial [Foreperiod(current=short) − Foreperiod(previous=short)], the Saccadic RTs were faster compared to when the condition strength was not reinforced [Foreperiod(current=short) − Foreperiod(previous=long)]; critically the FOE was also reduced, indicating that non-specific preparation influences the responsiveness of fixation neurons to visual signals. To examine whether strategic modulation and non-specific preparation engage independent mechanisms, an anti-saccade paradigm was employed in Experiment 2 to induce an inhibitory strategic set.
Experiment 2
In Experiment 2, the display and stimuli were identical to those used in Experiment 1, except that participants were required to perform an antisaccade task in which they had to inhibit the prepotent response of making a saccade to the target and, instead, to execute a voluntary saccade to the marker box in the visual field opposite the target. Previous research in humans has shown that the FOE is smaller in the antisaccade than the prosaccade task (Forbes and Klein 1996; Machado and Rafal 2000a; Machado and Rafal 2004; Reuter-Lorenz et al. 1991), and that in non-human primates, an instruction to execute an antisaccade is associated with an increase in neural activity in fixation neurons in the rostral pole of the SC (Everling et al. 1999). Thus, the strategic oculomotor set required to inhibit prosaccades results in a top-down activation of fixation neurons and reduces the influence of an external stimulus at the fovea on them. This experiment examined whether non-specific preparatory effects also exert an effect on the FOE that is independent from those engendered by a strategic oculomotor set. Twenty one undergraduate psychology students at the University of Wales, Bangor participated for course credit.
Results and discussion
Errors
Trials where subjects did not look at the centre of the screen at the start, blinked during the foreperiod, or moved their eyes before target onset, were scored as fixation errors. Only a low number of fixation errors were made (3.30%), and were therefore not further analyzed. If a reflexive eye movement was not successfully suppressed and subjects made an eye movement towards the target, the trial was scored as a direction error. An ANOVA of the direction error data revealed only a significant main effect of Fixation Point, F(1,20) = 5.91, P = 0.03, η 2 p = 0.23: Participants had more difficulties suppressing the VGR towards the visual target on trials with a fixation offset (4.2%) than when the fixation stimulus overlapped (3.0%) the target (see Table 2).
Saccadic RT
All errors were excluded from the Saccadic RT analyses. An initial analysis demonstrated that there was no significant difference between eye movements to the right and left (P > 0.5), and saccade RTs for leftward and rightward saccades were therefore pooled. The resulting mean saccadic RTs are shown in the bottom panel of Fig. 1. An ANOVA with Foreperiod(current), Foreperiod(previous), and Fixation Point Condition as within-subject factors, revealed the same main effects and interactions that were observed in Experiment 1. Saccade RT was shorter (i.e. FOE) on trials with a fixation point offset (342 ms), than on trials with a fixation point overlap (361 ms) [F(1,20) = 29.95, P < 0.001, η 2 p = 0.60]. The effect of foreperiod [Foreperiod(current)] was also reliable: saccade RT was longer for trials with a short foreperiod (367 ms), compared to trials with a long foreperiod (336 ms), F(1,20) = 102.91, P < 0.001, η 2 p = 0.84. In addition, there was a significant main effect of Foreperiod(previous). Saccade RT of trials which were preceded by a trial with a long foreperiod were longer (361 ms) than trials preceded by a short foreperiod (342 ms) [F(1,20) = 143.28, P < 0.001, η 2 p = 0.88]. There was an asymmetric effect of priming from the previous trial for short and long foreperiods: Foreperiod(current) × Foreperiod(previous), F(1,20) = 50.88, P < 0.001, η 2 p = 0.72. As was the case for prosaccades (Experiment 1) the FOE was modulated by preparatory state over the foreperiod [Foreperiod(previous) × Fixation Point Condition, (F(1,20) = 7.45, P = 0.01, η 2 p = 0.27)], and this modulation also reflected the asymmetric influence of priming by the preceding trial on preparatory state [Foreperiod(current) × Foreperiod(previous) × Fixation Point Condition, (F(1,20) = 9.40, P < 0.01, η 2 p = 0.32)]. Six pair wise comparisons (adjusted for Bonferroni) compared the size of the FOE for each Foreperiod(current) − Foreperiod(previous) condition. The FOE was significantly smaller for trials with a short foreperiod that were preceded by a short foreperiod (14 ms), compared to trials with a short foreperiod that were preceded by a long foreperiod (27 ms). None of the other pair wise comparisons were significant.
An 2 × 2 × 2 × 2 mixed AVOVA compared performance for prosaccades (Experiment 1) and antisaccades (Experiment 2) with Task as between subject factor, and Foreperiod(current), Foreperiod(previous), and Fixation Point Condition as within-subject factors. Saccade RT was longer for antisaccades (351 ms) than for prosaccades (306 ms) [F(1,34) = 12.5, P = 0.001, η p = 0.26). The FOE was smaller for the antisaccade (19 ms) than for the prosaccade (49 ms) task [F(1,34) = 13.64, P < 0.01, η 2 p = 0.29] demonstrating that the more inhibitory oculomotor strategic set required for antisaccades modulated the FOE. However, the influence of non-specific preparation on the size of the FOE was comparable [Task × Foreperiod(current), Foreperiod(previous), and Fixation Point Condition, F(1,34) < 1, P > 0.9, η 2 p < 0.01] in the two experiments (16 ms for prosaccades and 13 ms for antisaccades).
In addition to the ANOVA, six independent samples t tests were conducted with task as a grouping factor, and priming effect as testing variable. The priming effects were calculated by computing the difference in size of the FOE for all the six different Foreperiod(current) − Foreperiod(previous) conditions. For example, an independent samples t test examined whether the priming effect FOE(current=short)(previous=short) − FOE(current=short)(previous=long) was different for pro and anti-saccades. None of these t tests were significant (all P > 0.36), confirming the results of the 2 × 2 × 2 × 2 mixed AVOVA that the influence of non-specific preparation on the fixation reflex was the same for pro and anti-saccades.
Subjects were slower for anti-saccades than prosaccades. As expected, the size of the FOE was smaller in Experiment 2 (anti-saccades) than in Experiment 1 (pro-saccades). The results revealed that asymmetrical sequential effects were also present for the more voluntary anti-saccades. More interestingly, the size of the FOE was influenced by the amount of non-specific preparation. Like Experiment 1, the size of the FOE was the smallest when the non-specific preparation was the highest, i.e. for Foreperiod(current=short) − Foreperiod(previous=short) combinations.
Additional statistical tests found no significant difference in the effect of non-specific preparation on the size of the FOE between Experiment 1 and 2. This suggests that strategic modulation and non-specific preparation reflect independent processes.
General discussion
For both prosaccades and for antisaccades we observed a non-specific preparatory effect of priming from the previous trial. Consistent with Los’s (Los 1996; Los and Heslenfeld 2005; Los et al. 2001; Los and van den Heuvel 2001) trace conditioning account of non-specific preparation on response latency during an aging foreperiod, this priming effect was asymmetric: it occurred only for the short foreperiod. In addition to extending Los’s model to oculomotor preparation, we showed that the effect of non-specific preparation was comparable for reflexive prosaccades and for more voluntarily controlled antisaccades, indicating that this is a general mechanism that operates independently of the task that has to be performed. Sollers and Hackley (1997) have shown that the effect of an aging foreperiod on response latency is greater for voluntary (manual) than for reflexive (acoustic startle) blink responses. While prosaccades are not as reflexive as eye blinks—subjects are, after all, instructed to make a saccade to the target—they are clearly more reflexive than antisaccades. Our observations suggest that there is not a systematic relationship between the degree of automaticity of a response and the degree to which it is influenced by non-specific preparation. However, the foreperiod effects examined in the current investigation were quite different from those reported by Sollers and Hackley: their study examined foreperiods of several seconds, whereas the foreperiod range in the current investigation was much earlier and narrower.
By manipulating offset of the fixation point, it was possible to study the effect of both strategic set and non-specific readiness on reflexive fixation. We confirmed previous studies showing that the strategic set required to suppress the VGR in the antisaccade task reduces the FOE. We also showed that the non-specific effect of response readiness across an aging foreperiod induced by priming from the preceding trial modulated the size of the FOE. Specifically, the FOE was smallest when readiness to respond was the lowest. Critically, we also showed that the effect of trace conditioning in modulating the FOE was comparable for the two saccade tasks.
Thus, both non-specific and strategic preparatory cognitive processes are capable of independently influencing oculomotor reflexes prior to target onset. The observation that non-specific and strategic preparatory processes do not interact suggests that different neural processes may be involved.
The FEF have been shown to be important in the strategic control of eye movements in the context of the anti-saccades task, and have been shown to have a role in oculomotor fixation (Connolly et al. 2002; Everling and Munoz 2000; Umeno and Goldberg 1997). In other words, a critical role in controlling strategic preparatory processes has been attributed to the FEF. For example, Machado and Rafal (2004) showed that patients with chronic unilateral frontal eye fields (FEF) lesions are impaired in voluntary controlling their fixation reflex. The patients showed a reduced saccadic RT when cued to prepare a voluntary saccade to a specified location compared to when they received non-informative cues. Although the RT was reduced, there was no reduction in the FOE. This suggests that they could prepare an eye movement prior to the target onset, but that they lost the ability to modulate the activity of fixation neurons.
The role of the FEF in controlling the SC is also supported by neurophysiological data. Everling and Munoz (2000) studied set-related activity for saccadic eye movements of neurons in the frontal eye field with direct projections to the SC (identified by antidromic stimulation of SC neurons with receptive fields of 10°, 20° or <2°). Monkeys were trained on a task in which they were cued on each trial to execute either a prosaccade or an antisaccade. The activity of set-related FEF neurons was higher for prosaccades than for antisaccades. Also, the lower prestimulus and stimulus-related activity on anti-saccade trials compared with pro-saccade trials correlated with RT, express saccade occurrence, and errors in the anti-saccade task. These observations further support the view that the FEF exert strategic control over eye movements by reducing the excitatory drive from saccade-related FEF neurons to the SC during anti-saccade trials.
Frontal eye field neurons with projections to collicular fixation neurons were not identified or studied in this experiment. It remains to be determined whether decreased activity of presaccadic burst neurons in an antisaccade task set (Everling et al. 1999) is implemented by the FEF modulation through direct projections to fixation neurons, or whether fixation neuron responsiveness is controlled by indirect projections of the FEF through the basal ganglia.
In conclusion, the current research has examined automaticity and control in the oculomotor system and has demonstrated that both strategic and automatic preparation independently regulate oculomotor reflexes.
References
Connolly JD, Goodale MA, Menon RS, Munoz DP (2002) Human fMRI evidence for the neural correlates of preparatory set. Nat Neurosci 5(12):1345–1352
Dorris MC, Munoz DP (1995) A neural correlate for the gap effect on saccadic reaction times in monkey. J Neurophysiol 73:2558–2562
Easton TA (1972) On the normal use of reflexes. Am Sci 60(5):591–599
Everling S, Munoz DP (2000) Neuronal correlates for preparatory set associated with pro-saccades and anti-saccades in the primate frontal eye field. J Neurosci 20(1):387–400
Everling S, Dorris MC, Klein RM, Munoz DP (1999) Role of primate superior colliculus in preparation and execution of anti-saccades and pro-saccades. J Neurosci 19:2740–2754
Findlay JM, Walker R (1999) A model of saccade generation based on parallel processing and competitive inhibition. Behav Brain Sci 22:661–674
Forbes K, Klein RM (1996) The magnitude of the fixation offset effect with endogenously and exogenously controlled saccades. J Cogn Neurosc 8(4):344–352
Ingle D (1973) Evolutionary perspectives on the function of the optic tectum. Brain Behav Evol 8:211–237
Isa T (2002) Intrinsic processing in the mammalian superior colliculus. Curr Opin Neurobiol 12(6):668–677
Klein R, Kingstone A (1993) Why do visual offsets reduce saccadic latencies?. Behav Brain Sci 16:583–584
Lee P, Hall WC (2006) An in vitro study of horizontal connections in the intermediate layer of the superior colliculus. J Neurosci 26(18):4763–4768
Los SA (1996) On the origin of mixing costs: Exploring information processing in pure and mixed blocks of trials. Acta Psychol 94(2):145–188
Los SA, Heslenfeld DJ (2005) Intentional and unintentional contributions to nonspecific preparation: electrophysiological evidence. J Exp Psychol Gen 134(1):52–72
Los SA, Knol DL, Boers RM (2001) The foreperiod effect revisited: conditioning as a basis for nonspecific preparation. Acta Psychol 106(1–2):121–145
Los SA, van den Heuvel CE (2001) Intentional and unintentional contributions to nonspecific preparation during reaction time foreperiods. J Exp Psychol Hum Percept Perform 27(2):370–386
Machado L, Rafal RD (2000a) Control of eye movement reflexes. Exp Brain Res 135(1):73–80
Machado L, Rafal RD (2000b) Strategic control over saccadic eye movements: studies of the fixation offset effect. Percept Psychophys 62(6):1236–1242
Machado L, Rafal RD (2004) Control of fixation and saccades in humans with chronic lesions of oculomotor cortex. Neuropsychology 18(1):115–123
Munoz DP, Fecteau JH (2002) Vying for dominance: dynamic interactions control visual fixation and saccadic initiation in the superior colliculus. Prog Brain Res 140:3–19
Munoz DP, Istvan PJ (1998) Lateral inhibitory interactions in the intermediate layers of the monkey superior colliculus. J Neurophysiol 79(3):1193–1209
Munoz DP, Wurtz RH (1992) Role of the rostral superior colliculus in active visual fixation and execution of express saccades. Vis Neurosci 9:409–414
Munoz DP, Wurtz RH (1993a) Fixation cells in monkey superior colliculus. I. Characteristics of cell discharge. J Neurophysiol 70(2):559–575
Munoz DP, Wurtz RH (1993b) Fixation cells in monkey superior colliculus. I. Characteristics of cell discharge. J Neurophysiol 70:559–575
Niemi P, Näätänen R (1981) Foreperiod and simple reaction-time. Psychol Bull 89(1):133–162
Rafal R, Machado L, Ro T, Ingle H (2000) Looking forward to looking: saccade preparation and control of the visual grasp reflex. In: Monsell S, Driver J (eds) Control of cognitive processes: attention and performance XVIII. MIT Press, Cambridge, MA, pp 155–174
Rafal R, McGrath M, Machado L, Hindle J (2004) Effects of lesions of the human posterior thalamus on ocular fixation during voluntary and visually triggered saccades. J Neurol Neurosurg Psychiatry 75:1602–1606
Reuter-Lorenz PA, Hughes HC, Fendrich R (1991) The reduction of saccadic latency by prior offset of the fixation point: an analysis of the gap effect. Percept Psychophys 49(2):167–175
Rozin P (1976) The evolution of intelligence and access to the cognitive unconscious. Progr psychobiol physiol psychol 12(1):245–280
Saslow MG (1967) Effects of components of displacement-step stimuli upon latency for saccadic eye movements. J Opt Soc Am 57:1024–1029
Sollers JJ 3rd, Hackley SA (1997) Effects of foreperiod duration on reflexive and voluntary responses to intense noise bursts. Psychophysiology 34(5):518–526
Sumner P, Nachev P, Castor-Perry S, Isenman H, Kennard C (2006) Which visual pathways cause fixation-related inhibition? J Neurophysiol 95(3):1527–1536
Umeno MM, Goldberg ME (1997) Spatial processing in the monkey frontal eye field. I. Predictive visual responses. J Neurophysiol 78(3):1373–1383
Vallesi A, Shallice T (2007) Developmental dissociations of preparation over time: Deconstructing the variable foreperiod phenomena. J Exp Psychol Hum Percep Perform 33(6):1377–1388
Acknowledgments
This research was supported by the Wellcome Trust [079886]. We thank Sander Los and one anonymous reviewer for helpful comments on an earlier draft of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Van Koningsbruggen, M.G., Rafal, R.D. Control of oculomotor reflexes: independent effects of strategic and automatic preparation. Exp Brain Res 192, 761–768 (2009). https://doi.org/10.1007/s00221-008-1655-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-008-1655-7