
Abstract
The goal of this study was to examine the characteristics of planar fingertip movements with respect to the hand. Ten subjects with no known neuromuscular impairments performed a series of point-to-point movements with their dominant index fingertips. Subjects were instructed to move between five pairs of targets within the workspace of the index finger in each direction, for a total of ten separate movement tasks. We hypothesized that the trajectories with respect to the hand of these movements would exhibit curved paths contrary to the findings of similar hand path studies. The ratio of the path taken to the straight-line distance between the two targets was dependent upon the movement task (P<0.01), as was the mean residual between the actual and straight-line paths (P<0.001). For selected pairs of targets, these values were significantly different for the two opposing movement directions between a given pair of targets. This directional dependence of the curvature of the chosen finger-only trajectory observed in the initial protocol is incompatible with motor planning based solely on kinematic constraints, instead mechanical properties of the finger are likely incorporated.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
The primary function of the upper extremities is to interact with the environment. Point-to-point movements comprise a fundamental component of these interactions. As part of the motor control paradigm, a movement pathway between the starting and ending hand locations must be chosen from an infinite set of possibilities to execute a reach. The selected pathway under a given set of conditions has shown remarkable consistency within a single individual over multiple trials or even among multiple individuals (Soechting and Lacquanti 1981; Flash and Hogan 1985; Wolpert et al. 1995; Kamper et al. 2003), even in the presence of perturbing forces (Shadmehr and Mussa-Ivaldi 1994; Haggard and Wing 1995).
Many hypotheses have been postulated in an effort to explain the consistency of the pathways. As the trajectory of the hand tends to follow a straight-line in point-to-point movements with a single peak in tangential velocity, researchers have proposed the minimum jerk hypothesis (Flash and Hogan 1985), which predicts that trajectories are planned in a manner that minimizes the first derivative of acceleration, or jerk, resulting in the smoothest possible movement. While other minimization strategies for path selection have been suggested, such as minimization of torque change (Uno et al. 1989), metabolic energy consumption (Alexander 1997), and discomfort (Cruse et al. 1990), the majority of researchers support the concept that planning occurs with respect to kinematic constraints (Wolpert et al. 1995; Dingwell et al. 2004). Indeed, in planar reaching tasks the shape of the trajectory appears independent of the direction of movement, further suggesting that planning is performed with respect to kinematic constraints.
Similar control strategies are thought to be employed for control of the fingers. Although fingertip pathways during reach-to-grasp tasks tended to be curved, they could still be explained according to minimum jerk theory (Smeets and Brenner 1999). In a recent study, however, the fingertip trajectories with respect to the hand we observed did not correspond to a minimum jerk solution (Kamper et al. 2003). These movements, though, may have been confounded by the execution of functional grasping as part of the experimental protocol.
The experiments in this study, consequently, sought to determine the nature of true point-to-point movements of the finger. We hypothesized that the trajectories followed during execution of these tasks would be curved in nature with respect to the hand, and that the amount of curvature would vary with movement direction. Ten subjects were asked to perform point-to-point movements with their dominant index fingers. Specifically, we wanted to determine if the trajectories demonstrated consistent characteristics similar to those seen during grasping tasks and whether or not these characteristics varied throughout the workspace. Our results showed that subjects followed a consistent trajectory whose degree of curvature was dependent upon direction. These results indicate that movements are planned with regard to criteria additional to kinematic constraints.
Methods
Protocol
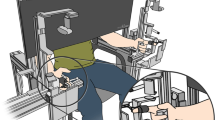
Ten subjects (six males, four females), aged 20–58 years, with no sign of neurological or joint impairment participated in this experiment. Each subject signed a consent form approved by the Northwestern University Institutional Review Board. The subject’s wrist was placed in a fiberglass cast that was clamped to the table top, preventing wrist and forearm movement, and keeping the palm of the hand perpendicular to the tabletop (Fig. 1a). This arrangement permitted flexion-extension of the three joints of the finger (metacarpophalangeal, proximal interphalangeal, and distal interphalangeal) in a plane parallel to the surface of the table. Fingertip location was tracked with a camera system (Optotrak 3020, Northern Digital Inc., Waterloo, Canada). A custom plastic fixture, which contained an IRED marker as well as a red light-emitting diode, was attached to the tip of subject’s index finger (Fig. 1b). The fixture was lightweight (<10 g) and did not impede range of motion within the testing workspace according to subject feedback. The Optotrak system sampled three-dimensional fingertip location at 100 Hz.

Experimental set-up. a Projector and camera set-up used to display targets and record movement. b LED fixture used to provide user feedback and fingertip tracking
A plastic screen was held in place above the index finger, parallel to the tabletop. Targets were projected onto this screen using an overhead projector (CD-450 m, BOXLIGHT, Poulsbo, WA, USA) mounted above the screen. Subjects were instructed to move between one of five pairs of targets. Eight of the targets were evenly distributed around the circumference of a circle whose diameter was maximized for the workspace of each finger using the segment lengths of the subject (average circle diameter = 4.6 cm). Pairs of targets were located opposite each other, separated by the circle diameter. Two more targets were located twice the diameter of the circle apart, spanning the bulk of the theoretical workspace of the finger (Fig. 2).

Target locations within finger workspace. Movement task is denoted by starting point to stopping point, e.g., 0-4, 4-0, 1-5, 5-1, 2-6, 6-2, 3-7, 7-3, 8-9, and 9-8
Movement tasks consisted of moving between pairs of targets, presented in a randomized order. The LED in the fingertip fixture projected upwards through the screen to provide visual feedback of the fingertip location to the subject. Before the trial began, one green target was illuminated to guide the subject to the starting position. When the red light from the fingertip LED was properly aligned with the green target, the target changed color to yellow, indicating to the subject that the target had been reached. With the subject properly aligned with that starting position, sampling of the fingertip location was initiated. After a 500-ms delay, a second target was illuminated and the first target was erased. The subject moved his/her fingertip to the new position, using the same visual feedback of change in target color to indicate that the target had been reached. This process constituted a single trial. For consistency, audio and visual cues were used to guide subjects to complete the movement at 0.25 s duration. A total of ten movements were performed for each movement task, thereby yielding a total of 100 trials for each subject.
Three of the original ten subjects (all males) participated in a follow-up study to ensure that biomechanical constraints did not limit the straightness of the trajectories between the targets. Subjects were again instructed to move their fingertips between pairs of targets arranged as in the previous study. For the new protocol, a straight line was also projected, connecting the initial point to the target point. Subjects were instructed to follow this straight line with their fingertip to the best of their ability. To facilitate accurate movement along the line, no time constraints were imposed for these movements.
Data analysis
The kinematic data were digitally low-pass filtered forward and backward with a 30th-order finite impulse response filter having a cut-off frequency of 10 Hz. The start and stop of movement for each trial were found using the tangential velocity of the fingertip marker. The tangential velocity was calculated through a 5-point differentiation of the x- and y-position vectors. Start and stop were defined as the times at which tangential velocity either exceeded or dropped below, respectively, a threshold value set at 10% of the maximum tangential velocity. Each trial was resampled along the Cartesian axis of greatest change to create data sets with an equal number of points between the start and stop points. For each movement the straight-line distance between the start and stop of movement was computed (d), as was the distance of the path taken between the start and stop points (r) (Fig. 3). The ratio of r/d was used to compute the path length ratio (PLR). The path shape was computed by calculating the average path between the start and stop point with a 95% confidence interval for all path data between the same targets. The mean residuals from the straight-line fit were calculated to examine the consistency of any deviation from a straight-line path. The mean square error from a straight-line path was also computed for each trial.

Graphical representation of path length ratio (PLR) and mean square error data analysis. The gray shadow represents the 95% confidence interval about the averaged trajectory. Dots represent actual data points. Grey cloud represents the 95% confidence interval of the pathway
Details of the velocity profiles for the movements were also analyzed. The peak tangential velocity was determined for each trial. The number of velocity peaks during a task was found by searching the velocity signal for local minima and maxima. A peak occurred if the maximum velocity between local minima and maxima exceeded 20% of the peak velocity for that trial, in accordance with a previous study (Kamper et al. 2002b).
Repeated measures anova was used to analyze both the PLR and mean residual data, with movement task serving as the within-subject factor. Using the conservative Bonferroni estimate of overall significance, a significance level of α=0.025 was employed for each individual anova to ensure an overall significance level of α=0.05. Results from the initial protocol and follow-up protocol were compared using a two-sided paired T-test of significance.
Results
Ten subjects with no known neuromuscular deficits performed a total of 100 trials, representing ten trials each of a single movement task. Across subjects, the PLR was dependent upon the movement task (P<0.01, Fig. 4). For example, movements directly toward (4-0) or away (0-4) from the palm, had significantly smaller PLR values (P<0.02) than movements parallel to the palm (2-6, 6-2). The mean PLR for a few of these directions approached a value 10% greater than that of the corresponding straight-line movement. For two pairs of movements with the same end points, differing only by direction, the PLR was significantly (P<0.03) different between the two directions (1-5 and 5-1; 8-9 and 9-8, see Fig. 4).

Summary of path length ratio (PLR) data, grouped by movements between the same endpoints in opposite directions. Error bars represent 95% confidence intervals
The area representing a 95% confidence interval deviation from the average path was found for each movement task. Across all tasks, 49.2% of all points along a mean trajectory were significantly (P<0.05) different than the corresponding straight line. The mean residuals between this average path and the straight-line fit were computed for each of the ten distinct movements.
Mean residual was also dependent upon movement task and direction (P<0.001). The mean residual was positive for some movement tasks and negative for others (Fig. 5). Six of these movements (1-5, 4-0, 6-2, 7-3, 8-9, and 9-8) demonstrated mean residuals that were consistently significantly (P<0.05) deviated to one side of the straight line connecting the two points. For three pairs of movements with the same end points, (0-4 and 4-0; 3-7 and 7-3; 8-9 and 9-8), the mean residual was significantly higher for movement in one direction than for the other direction (P<0.03, see Fig. 6 for an example of one pair).

Summary of mean residual data, grouped by pairs of movements between the same endpoints but in opposite directions. Error bars represent 95% confidence intervals

Averaged trajectories (solid black line) with the 95% confidence interval (gray shadow) between two points (8, 9) in opposite directions for a single subject
Three subjects repeated the 100 trials, with the added cue of a straight-line guide. While the results for the PLR of the repeated subjects (1.03±0.02) was slightly lower than for the same subjects during the initial protocol (1.04±0.03), this difference was not statistically significant (P=0.16). However, the mean square error from the straight line for the follow-up trials (0.37 cm) was significantly smaller (P<0.001) than for trials performed during the first testing session (0.83 cm, see Fig. 7 for an example). Only two movement tasks (0-4, 6-2) demonstrated a mean residual significantly different (P<0.05) than zero when subjects were attempting to follow the straight-line guides.

Comparison of average trajectory shape for a single subject without the straight-line cue (dark shadow) and with cue (light shadow). The dashed line depicts the actual straight line connecting the endpoints 3 and 7
Qualitatively, it was more difficult for subjects to move along the straight-line cue than to move without a guide. As a result, the peak velocity for the follow-up protocol (4.5 cm/s) was significantly (P<0.001) lower than for the same subjects during the initial protocol (9.6 cm/s). Additionally, the number of velocity peaks during the movements (2.8) significantly (P<0.005) increased as compared to the initial protocol (1.2). For one task (7-3), there was even a learning effect evidenced by a significant (P=0.01) decrease in error over time for the repeated movement as determined from a linear regression analysis of error versus trial number.
Discussion
Ten subjects with no known neuromuscular impairments performed a series of point-to-point movements with their dominant index fingers. Contrary to data found in similar arm-reaching tasks (Flash and Hogan 1985; Shadmehr and Mussa-Ivaldi 1994), the fingertip trajectories with respect to the hand across subjects demonstrated a dependence upon movement task and location within the workspace. The path length was greater for some movement tasks than others. For two pairs of targets, the pathway length was significantly different across subjects depending solely on the direction of movement. This should not occur if movement were governed by purely kinematic constraints. Thus, other criteria must factor into the motor planning.
Movement trajectories were consistently curved in a specific direction for a given task. The 95% confidence interval for each trajectory did not include the straight-line pathway for 50% of its length and the mean residual was significantly different than zero for the majority of movement tasks. The magnitude of the residual was dependent upon movement task and direction. For multiple pairs of targets, the mean residual for movement between the targets in one direction was greater than for movement between the targets in the opposite direction.
This curvature in finger pathway has been described previously by other researchers (Smeets and Brenner 1999). They discussed how for their task the curved pathway could result from motion planning with respect to kinematic constraints, namely minimum jerk. This would require non-zero acceleration at the endpoint, however, which was not feasible for our task. Additionally, the curvature we observed varied with movement direction. This result is counter to the minimum jerk hypothesis.
The chosen trajectories did not result solely from physical constraints on joint rotation. While mechanical coupling is present between the distal and proximal interphalangeal joints, subjects were still able to follow a much straighter pathway when instructed to do so. This movement, however, was more difficult for them to perform. Velocities were halved (although it should be noted that no time constraint was imposed on the movement with straight-line cues) and the number of velocity peaks was more than doubled.
The directional dependence and curvature of the chosen fingertip trajectory may relate to the passive impedance of the finger joints. Dynamic considerations may be more important for the fingers than the arm because of greater relative passive joint impedance. The passive stiffness of the elbow is quite low (Zhang and Rymer 1997) when compared to other joints of similar size such as the knee (Stein et al. 1996) or ankle (Mirbagheri et al. 2000). When normalized by the voluntary moment that can be produced at the joint, passive joint torques for the fingers are roughly 5–10 times as great as the normalized passive joint torques for the elbow (Kamper et al. 2002a; Mirbagheri 2005 unpublished data; Schmit 2005 unpublished data). Additionally, elbow stiffness has been shown to change little with angle across much of the elbow range of motion (Mirbagheri et al. 2003). In contrast, finger joint stiffness has been shown to exhibit a parabolic relationship with joint angle (Kamper et al. 2002a), suggesting a greater relative cost for attaining certain finger postures than seen at the elbow.
Finally, neuromechanics may play a large role in path selection. Muscles controlling the fingertip typically cross three or more joints and are constrained within the hand by ligamentous tissues on both the palmar and dorsal aspects of the hand. Tendons from five different muscles converge to form the extensor hood. Thus, activation of a single muscle influences several degrees of freedom and impacts the effects of other muscles on the finger. Researchers have theorized that the nervous system reduces the control complexity inherent to the large number of actuating muscles and degrees of freedom by the use of synergistic combinations of muscle activation patterns. Studies have shown that hand postures during a wide range of movements are often comprised of a limited set of finger postures (Soechting and Flanders 1997; Santello and Soechting 1998; Mason et al. 2001). Similarly, force sharing among digits seems to follow fixed patterns (Li et al. 1998; Kang et al. 2004). The pathways in our study may represent the simplest solutions from a control aspect.
In conclusion, while the chosen trajectories for fingertip point-to-point movements were very consistent across subjects, they differed significantly depending on the direction of movement within the finger workspace. Thus, motor planning for the fingers seems to include factors other than uniform kinematic constraints.
Abbreviations
- PLR:
-
Path length ratio
References
Alexander RM (1997) A minimum energy cost hypothesis for human arm trajectories. Biol Cybern 76:97–105
Cruse H, Wischmeyer E, Bruwer M, Brockfeld P, Dress A (1990) On the cost functions for the control of the human arm movement. Biol Cybern 62:519–528
Dingwell JB, Mah CD, Mussa-Ivaldi FA (2004) Experimentally confirmed mathematical model for human control of a non-rigid object. J Neurophysiol 91:1158–1170
Flash T, Hogan N (1985) The coordination of arm movements: an experimentally confirmed mathematical model. J Neurosci 5:1688–1703
Haggard P, Wing A (1995) Coordinated responses following mechanical perturbation of the arm during prehension. Exp Brain Res 102:483–494
Kamper DG, Cruz EG, Siegel MP (2003) Stereotypical fingertip trajectories during grasp. J Neurophysiol 90:3702–3710
Kamper DG, Hornby TG, Rymer WZ (2002a) Extrinsic flexor muscles generate concurrent flexion of all three finger joints. J Biomech 35:1581–1589
Kamper DG, McKenna-Cole AN, Kahn LE, Reinkensmeyer DJ (2002b) Alterations in reaching after stroke and their relation to movement direction and impairment severity. Arch Phys Med Rehabil 83:702–707
Kang N, Shinohara M, Zatsiorsky VM, Latash ML (2004) Learning multi-finger synergies: an uncontrolled manifold analysis. Exp Brain Res 157:336–350
Li Z, Latash ML, Zatsiorsky VM (1998) Force sharing among fingers as a model of the redundancy problem. Exp Brain Res 119:276–286
Mason CR, Gomez JE, Ebner TJ (2001) Hand synergies during reach-to-grasp. J Neurophysiol 86:2896–2910
Mirbagheri MM, Barbeau H, Kearney RE (2000) Intrinsic and reflex contributions to human ankle stiffness: variation with activation level and position. Exp Brain Res 135:423–436
Mirbagheri MM, Chen D, Harvey RL, Rymer WZ (2003) Identification of reflex and intrinsic mechanical properties in stroke and spinal cord injury. 25th IEEE Eng Med Biol Soc Annual International Conference. Cancun, Mexico
Santello M, Soechting JF (1998) Gradual molding of the hand to object contours. J Neurophysiol 79:1307–1320
Shadmehr R, Mussa-Ivaldi FA (1994) Adaptive representation of dynamics during learning of a motor task. J Neurosci 14:3208–3224
Smeets JB, Brenner E (1999) A new view on grasping. Motor Control 3:237–271
Soechting JF, Flanders M (1997) Flexibility and repeatability of finger movements during typing: analysis of multiple degrees of freedom. J Comput Neurosci 4:29–46
Soechting JF, Lacquanti F (1981) Invariant characteristics of a pointing movement in man. J Neurosci 1:710–720
Stein RB, Zehr EP, Lebiedowska MK, Popovic DB, Scheiner A, Chizeck HJ (1996) Estimating mechanical parameters of leg segments in individuals with and without physical disabilities. IEEE Trans Rehabil Eng 4:201–211
Uno Y, Kawato M, Suzuki R (1989) Formation and control of optimal trajectory in human multijoint arm movement. Minimum torque-change model. Biol Cybern 61:89–101
Wolpert DM, Ghahramani Z, Jordan MI (1995) Are arm trajectories planned in kinematic or dynamic coordinates? An adaptation study. Exp Brain Res 103:460–470
Zhang LQ, Rymer WZ (1997) Simultaneous and nonlinear identification of mechanical and reflex properties of human elbow joint muscles. IEEE Trans Biomed Eng 44:1192–1209
Acknowledgments
This study was supported by the Department of Veterans Affairs and the Coleman Foundation.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Cruz, E.G., Kamper, D.G. Kinematics of point-to-point finger movements. Exp Brain Res 174, 29–34 (2006). https://doi.org/10.1007/s00221-006-0416-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-006-0416-8