
Abstract
Reflex responses to unexpected stretches are well documented for selected muscles in both animal and human. Moreover, investigations of their possible functional significance have revealed that stretch reflexes can contribute substantially to the overall stiffness of a joint. In the lower extremity only the muscles spanning the human ankle joint have been investigated in the past. This study implemented a unique hydraulic actuator to study the contributions of the knee extensor stretch reflex to the overall knee joint torque. The quadriceps muscles were stretched at various background torques, produced either voluntarily or by electrical stimulation, and thus the purely reflex mediated torque could be calculated. The stretch had a velocity of 67°/s and an amplitude of 20°. A reflex response as measured by electromyography (EMG) was observed in all knee extensors at latencies of 26 – 36 ms. Both phasic and tonic EMG stretch responses increased with increasing background torques. Lines of best fit produced correlation coefficients of 0.59 – 0.78. This study is the first to examine the reflex contribution of the knee extensors to the total torque at background torques of 0 – 90% MVC. The contribution of the reflex mediated torque is initially low and peaked at background torques of 20 – 40% MVC. In terms of the total torque the reflex contributed 16 – 52% across all levels of background torque. It is concluded that during medium background torque levels such as those obtained during walking, the stretch reflex of the quadriceps muscle group contributes substantially to the total torque around the knee joint.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
In both human and animal reflex responses to sudden changes in muscle length are well documented in the literature (Allum and Mauritz 1984; Allum et al. 1982; Kearney et al. 1997; Lin and Rymer 1993; Nichols and Houk 1976; Sinkjaer et al. 1988; Toft et al. 1991; Yang et al. 1991). Recently evidence has been presented that the stretch reflex also contributes to human muscle activity in unperturbed movements (Nielsen and Sinkjaer 2002; Sinkjaer et al. 2000). The functional role of the stretch reflex in terms of its mechanical significance is still controversial. Continuous substantiation that the stretch reflex is strongly modulated during different tasks suggests that it may play a significant role at some times but not at others (Capaday 2002; Grey et al. 2002; Stein and Capaday 1988).
It has been suggested that the stretch reflex not only increases muscle and thus joint stiffness but also allows for a more uniform stiffness over a diverse range of the muscles physiological operating lengths and tension (Allum et al. 1982; Nichols and Houk 1976). In the cat the reflex-mediated stiffness has been shown to be independent of changes in muscle length (Hoffer and Andreassen 1981; Nichols and Houk 1976), indicating that the short latency reflex is controlled by muscle and/or joint stiffness rather than by purely muscle length changes. Human studies have revealed that the stretch reflex at low and medium level contractions contributes importantly (approximately 50%) to the overall joint stiffness (Crago et al. 1976; Sinkjaer et al. 1988; Stein and Kearney 1995; Thomson and Chapman 1988). This has led to the suggestion that the short latency stretch reflex is part of a central control strategy where the central nervous system (CNS) as the first part of its defense to an unexpected joint perturbation stiffens up the joint (Sinkjaer 1997).
To determine the contribution of the reflex to the total torque produced around a joint several researchers have taken the approach of investigating the mechanical state before and after deafferentation. The latter state was accomplished by various techniques such as cutting the nerve supply (Hoffer and Andreassen 1981; Nichols and Houk 1976), imposing ischemia (Allum and Mauritz 1984; Sinkjaer and Hayashi 1989), electrical stimulation (Carter et al. 1990; Sinkjaer et al. 1988), administering drugs, and more recently through mathematical modeling (Kearney et al. 1997; Mirbagheri et al. 2000; Stein and Kearney 1995). During low ankle extensor contraction levels up to two-thirds of the total stiffness arose from the reflex mediated stiffness when measured 200 ms after stretch onset (Toft et al. 1991). At higher contraction levels the reflex contribution became less. In contrast, dissimilar results have been reported for muscles acting across the wrist and elbow joints of the upper limb. Here it appears that the reflex contribution to joint stiffness increases across all levels of background torque (Carter et al. 1990; Crago et al. 1976).
Information on the mechanical properties of the human knee joint is scarce. A joint driving device was implemented by Zhang et al. (1998) to examine the total knee joint stiffness changes as a function of both joint position and muscle contraction. Stiffness was highest at a knee joint angle of 30° flexion increasing progressively as joint torque was increased. Unfortunately their set-up did not allow them to determine reflex and nonreflex contribution to total joint torque.
Although the electromyographic (EMG) stretch reflex of the knee extensors has been described in the past (Dietz et al. 1990; Lamontagne et al. 2000; Mrachacz et al. 2000) its importance from a mechanical point of view has only been investigated in the cat (Wilmink et al. 1997). During a dynamic task such as walking the size of the reflex is modulated in such a way that it shows higher magnitudes while the leg is loaded during the stance phase (Mrachacz et al. 2000). In the human leg only ankle joint stiffness has been investigated in the past.
The aim of this study was to examine the relative contributions of the quadriceps stretch reflex-mediated torque to the total torque of the knee joint at various background torques and to investigate whether similar stretch reflex control takes place at the knee joint as earlier reported for the ankle joint.
Methods
Ten healthy subjects aged 26.5±4.5 years participated in the study. All subjects gave their informed consent. The local ethics committee, in conformation with the Declaration of Helsinki, approved the study.
Apparatus and instrumentation
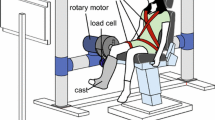
The subject was seated in a fixed chair with the hip at 90° flexion and the knee at 30° flexion. This knee angle was chosen, as at this position the total joint stiffness is maximal (Zhang et al. 1998). The right leg was affixed to a PID controlled hydraulic actuator (MTS Systems, 215.35; Voigt et al. 1999), such that the anatomical knee axis of rotation was closely aligned with the fulcrum of the actuator. This ensured that any movement at either the hip or ankle joint due to the rotation of the knee joint was kept at a minimum such that stretch reflexes were elicited solely in the knee extensors. The foot and leg segments of the right leg of the subject were firmly strapped to a custom-made plate, which extended out from the actuator, via velcro straps, producing a tight interface between the arm of the motor and the leg of the subject. In addition a custom made adjustable bar was attached to the chair. It was placed over the upper thighs of the subjects just behind the knee joint. The bar served to restrict the subjects' impulse to rise up from the chair during the trials at higher background torques. The plate was instrumented with an accelerometer (Kistler, Piezotron) and a load cell (Kistler, Slimline), which were used to measure the force exerted at the plate by the actions of the knee extensors. The position of the rotary actuator was monitored with an angular displacement transducer (Transtek DC ADT series 600).
Pairs of surface electrodes (Medicotest 720-01-K) were used to record the EMG of rectus femoris (RF), vastus lateralis (VL), vastus medialis (VM), biceps femoris (BF), and medial hamstrings (MH) of the left leg. The electrodes were placed following the recommendations of Garland et al. (1994). The EMG signals were amplified and band pass filtered at 20 Hz–2 kHz, rectified and first-order low pass filtered at 40 Hz (Butterworth 1st order digital filter). All data were sampled at a frequency of 4 kHz.
Data collection
The torque applied at the plate by the subject was displayed on an oscilloscope. Initially a maximal voluntary contraction (MVC) of the quadriceps was performed to determine the maximum possible force, which the subject could voluntarily exert at the plate. Subjects were instructed to extend their knee joint and thus lower leg against the plate as powerful as possible on the word go and maintain it there until instructed to relax. They were then allowed to relax for 1–3 min prior to the next trial. The best of a total of three collected trials was deemed the subject's MVC. To determine increments of torque as a result of the stretch, the actuator applied stretches with an amplitude of 20°, a rise time of 300 ms, and a duration of 500 ms.
Measurement of non-reflex torque
During the imposed stretch, the femoral nerve innervating the quadriceps muscle complex was stimulated electrically. It is known that all reflex activity is abolished during electrical stimulation high enough to produce a tetanic contraction (Hoffer and Andreassen 1981). The resulting measure of torque can thus be attributed to the intrinsic properties of the contracting muscle. A suitable position for stimulation, defined as the site where a maximal M-wave was produced in the RF with minimal activity from the synergists VL and VM and no activity from the antagonists BF and the MH, was located. Palpation of RF, VL, VM, BF, and MH muscles was performed during stimulation trials to ensure that this was occurring. The femoral nerve was stimulated in the femoral triangle. A constant current isolated stimulator (Axon Isolator-11) provided 1 ms squared pulses at a frequency of 1 Hz and various manually adjusted amplitudes. No stretches were applied during these trials. In this way the input-output relationship between stimulation amplitude and peak to peak M-wave amplitude was obtained. It also served as an indication of the maximum M-wave for the RF (M-max).
To be able to measure the non-reflex torque increment, the pulse frequency was increased to 20 Hz. This ensured that a fused contraction of the RF was produced which exerted a constant force on the plate. Five consecutive stretches were now applied at the knee joint while the EMG, force, and position data were collected. The peak to peak M-wave was monitored on-line to ensure a constant stimulus to the RF muscle as well as to monitor possible unwanted reflex responses and activation of synergistic muscles. To attain various background torque levels, the stimulus intensity was varied manually for each subsequent trial. Depending on the subject, five to eight torque levels were attained in this way.
Measurement of total torque
To determine the total torque increment, the above trials were repeated, while the subjects were asked voluntarily to reproduce the forces attained during the electrical stimulation by matching the torque level which was preset on an oscilloscope. During all trials the subjects were asked to maintain the background torque without interfering with the imposed stretch.
Data analysis
The increments in torque from the stretch, in relation to the background torque were measured from averaged signals at 300 ms following stretch onset. Non-reflex torque was subtracted from the total torque at matched background torques to obtain the torque attributable to the stretch reflex (Sinkjaer et al. 1988).
The EMG signals for the RF, VL, and VM were averaged and the onset latencies determined by visual inspection. The onset was defined as the first major deflection in the rectified EMG signal following the imposed stretch. The average phasic reflex response was defined as the increase in the EMG signal during the ramp part of the imposed stretch. It was measured from the onset of stretch response until a distinct decrease was observed. The total time window for the phasic reflex response was 140 ms. The average tonic reflex response was defined as the increase in EMG during the hold phase of the imposed stretch. It was measured from 300 ms until 500 ms following stretch onset.
Results
Electromyographic results
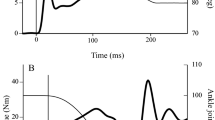
Figure 1 is an example from a recording during a voluntary contraction. Data are the average of ten trials from one subject. The upper two curves show the position of the knee (Fig. 1A) and the total torque response (Fig. 1B) prior to, during and after an imposed stretch. The background torque indicated by the arrow (Fig. 1B) is 30 Nm, which represents 22% of this subjects MVC. As the stretch is applied, the total torque increases reaching its maximum 200 ms after stretch onset. There it remained until the cessation of the stretch (indicated by arrows). Interestingly, the maximum total torque occurred while the amplitude of the stretch was still increasing. The small oscillations observed in the torque trace following stretch onset are most likely the result of movement of the foot segment due to the perturbation. Great care was taken to strap the foot, tightening the interface between the foot and lower leg segments. However, to avoid occlusion of blood flow, it was not possible to attain a completely rigid ankle joint. The lower two curves indicate the associated responses from one knee extensor (RF, Fig. 1C) and one knee flexor (BF, Fig. 1D). The initial RF EMG response to the imposed displacement commences 31±5 ms following stretch onset (Fig. 1C). For VL and VM the reflex appeared at 32±5 ms and 33±3 ms, respectively (not shown). A typical response consists of a number of peaks followed by a more tonic response (Fig. 1C). The small increase in EMG of the knee flexors (Fig. 1D) is attributed to cross-talk. In Fig. 1 the vertical scale for the RF and BF are maintained the same to illustrate this. We chose to analyze a phasic and a tonic reflex response (Toft et al. 1991, 1993). The phasic response continued for approximately 100–120 ms following response onset (Fig. 1C). During this time the torque had reached its maximum value. Typically a silent period followed where after a tonic reflex response was observed for the remainder of the stretch. At the cessation of stretch both the tonic reflex response and the total knee torque decreased towards background level. The changes in magnitude of both phasic and tonic reflex response with increases in background torque are shown in Fig. 2. For all knee extensors investigated, both the phasic and tonic reflex responses increased with increases in background torque. The two are similar in magnitude. Also displayed in the graph are the lines of best fit for the data with their associated correlation coefficients (r) values. These ranged from 0.59 to 0.78.

A Knee flexion angle. B Torque at plate. C Rectified rectus femoris (RF) EMG. D Rectified biceps femoris (BF) EMG. Data are averaged over ten stretches. Background torque represents 30% of the subject's MVC. Phasic and tonic EMG increments were measured as the mean EMG from stretch reflex onset for a duration of 140 ms and from 300 to 500 ms after stretch onset. All increments were calculated relative to the average background EMG prior to the onset of stretch

Averaged rectified phasic and tonic rectus femoris (RF) EMG measurements for all subjects as a function of background torque. B Averaged rectified phasic and tonic vastus lateralis (VL) EMG measurements for all subjects as a function of background torque. C Averaged rectified phasic and tonic vastus medialis (VM) EMG measurements for all subjects as a function of background torque. Linear lines of best fit with associated R values are also shown. In all graphs the solid line indicates tonic EMG increment, and the dotted line indicates phasic EMG increment
Mechanical results
Non-reflex torque
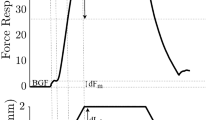
Figure 3 illustrates a recording of non-reflex torque. The upper two curves indicate the position of the knee joint (Fig. 3A) and the torque increment (Fig. 3B). Data are averages of five stretches for one subject. The stimulus intensity was set to 20 mA resulting in a background torque of 40 Nm (indicated by the first arrow in Fig. 3B). This represents 25% of the maximum force this subject can produce by voluntarily contracting the knee extensors. As the stretch is applied, the non-reflex torque commences to increase attaining its maximum value at approximately 180–200 ms following stretch onset (indicated by second arrow in Fig. 3B). It then remains relatively stable until cessation of the stretch when it then decreases to background levels.

A Knee flexion angle. B Torque at plate. C Raw rectus femoris (RF) EMG. Data are of a single perturbation during electrical stimulation. Background torque represents 30% of the subject's MVC
The lower trace of Fig. 3 is the recorded EMG from one knee extensor (RF, Fig. 3C). It shows both the stimulus artifact and the stable M-wave, which appeared approximately 3–5 ms following the stimulus. Trials in which the peak to peak M-wave varied by more than 5% were discarded, as were trials in which the antagonists showed activation. From time to time a Hoffman reflex appeared with the M-wave. These trials were not discarded, except if the H-reflex varied in amplitude throughout the perturbation, as it is believed that the intrinsic properties of a muscle activated by a H-reflex do not differ from when it is activated by an M-wave (Sinkjaer 1997).
Total torque vs. non-reflex torque
For the first 60–80 ms following the onset of the stretch the non-reflex torque as recorded during electrical stimulation and the total torque increment recorded during voluntary contraction exhibit the same pattern (Fig. 4B). This similarity is in part due to the inertia of the moved plate, leg and foot segment (Allum and Mauritz 1984). Following this initial period the non-reflex torque increased further but to a lesser degree than the total torque, until it reached its maximum value 150 ms after stretch onset, at which it stabilized. It returned towards the level of background torque either at the cessation of the stretch or slightly earlier.

A Knee flexion angle. B Time course of total and non-reflex torque of knee extensors in response to a 20° stretch. Data are averages over ten trials for one subject. The background torque represents 30% of the subject's MVC
The total torque reached its maximum at approximately 200–300 ms after stretch onset. It remained constant until cessation of the stretch when it decreased to the background torque. The difference between the nonreflex and total torque in Figure 4 is attributed to the stretch reflex. The associated EMG is shown in Fig. 2. The following paragraphs present the relationship of total, nonreflex and reflex torque increments to background torque at 300 ms following stretch onset.
Total, reflex, and non-reflex torque increments at different levels of background force
Non-reflex torque increased linearly with background torque (Fig. 5A). The line of best fit for the data from all subjects has a correlation coefficient of 0.90. Regression lines for individual subjects demonstrate an even stronger relationship. Correlation coefficients range from 0.96 to 0.99 (Fig. 5B).

A Non-reflex torque increments with changing background torques measured 300 ms following the onset of the stretch. Each data point represents the average of ten trials for one subject. Also shown is the linear function fitted to the data. B Regression lines of incremental non-reflex torque vs. background torque for individual subjects. Measures were made 300 ms following the onset of the stretch
The upper part of Fig. 6 shows the total torque increment vs. background torque for all subjects. A second order polynomial with a correlation coefficient of 0.90 could be fitted to the data. The total torque increased without peaking even at high levels of background torque. Only two subjects were able to maintain background torques above 70% MVC during the voluntary contraction. In the lower part of Fig. 6 polynomial regression lines of incremental total torque compared to background torque for individual subjects have been fitted to the data. Correlation coefficients ranged from 0.95 to 0.99.

A Total torque increments with changing background torques measured 300 ms following the onset of the stretch. Each data point represents the average of ten trials for one subject. Also shown is the polynomial function fitted to the data. B Regression lines of incremental total torque vs. background torque for individual subjects. Measures were made 300 ms following the onset of the stretch
The reflex torque was calculated by subtracting the non-reflex torque from the total torque increment at 300 ms following stretch onset (Fig. 4B). The upper trace of Fig. 7 displays the reflex mediated torque in relation to the background torque for all subjects. The lower trace of Fig. 7 exhibits fitted second order polynomials to the data of individual subjects. Individual subjects vary in the magnitude of reflex torque contribution to the total joint torque.

A Reflex torque increments with changing background torques measured 300 ms following the onset of the stretch. Each data point represents the average of ten trials for one subject. B Regression lines of incremental reflex torque vs. background torque for individual subjects. Measures were made 300 ms following the onset of the stretch
Total torque, non-reflex torque, and reflex torque across various background torques for one subject are shown in the upper trace of Fig. 8A. Note that at lower background torques the total torque and non-reflex torque are almost identical. With increases in background torque the contribution from reflexes rapidly increases to a background torque of 30–40% MVC. Data were also pooled for all subjects and averaged in ten blocks of 10% increments of background torque (Fig. 8B). The abscissa expresses the reflex torque as a percentage of the total torque. Reflex contribution to total torque was found to peak between 20% and 40% of background torque attaining values of approximately 50% of total torque. Across all background levels of torque the subjects showed an average reflex mediated contribution of 16–52% of total torque.

A The total and non-reflex torque plotted against the background torque at 300 ms after knee extensor stretch for one subject. Data points indicate means over ten stretches. B Reflex torque increments as a percentage of total torque with changing background torques measured 300 ms following the onset of the stretch. Data were averaged for 10% background torque windows. Standard deviations are also shown
Discussion
Methodological considerations
In this study electrical stimulation was used to obtain the intrinsic torque contribution to the total joint torque. It is well known that the recruitment pattern of motor units differs when this is performed electrically vs. voluntarily (Nichols and Houk 1976). Muscle activation is changed not only by recruitment of more motor units but also by increased firing rate. In this study we maintained the stimulation frequency constant and varied only the amplitude of the stimulation. In the cat soleus muscle it has been shown that the force resulting from stretch does not vary if stimulation frequency is altered, or if the crossed extensor reflex is used to recruit motor units (Nichols and Houk 1976). Carter et al. (1990) found no significant differences in torque produced with either amplitude or frequency of stimulation. It may be argued, however, that since the knee joint is comprised of a number of extensor muscles it may be difficult to accurately match the conditions during voluntary contractions vs. electrical stimulation. In this study torque records produced by either method prior to possible effects of the reflex are closely matched. This lends support to the notion that the non-reflex torque contribution to the total torque was accurately predicted.
When a sudden and unexpected stretch is applied to a joint, the muscle being stretched responds with a reflex. The latency of this is short enough to be comparable to the myotatic reflex described by Liddell and Sherrington (1924). This short latency reflex component is usually accompanied by later occurring reflex responses. Recent publications have increased our understanding of the origins of these responses immensely, although some controversies remain (Christensen et al. 2000; Grey et al. 2001; Nardone et al. 2000; Schieppati and Nardone 1999).
Phasic and tonic reflex responses with increases in background torque
The magnitudes of the various components of the quadriceps reflex response with increases in background torque have been described in the past (Lamontagne et al. 2000). In this study the slow velocity of the imposed stretch produced a reflex response without discernible peaks. Studies on the human ankle joint have revealed quite distinct peaks in the EMG response to an imposed stretch (Toft et al. 1991). These are likely to be mediated by different afferents and through different pathways (Grey et al. 2001). In this study the knee joint was examined, and these clearly defined peaks are not so evident here. This may simply be due to the closer proximity of the knee joint to the spinal cord and therefore the muscles to the spinal circuitry. This may cause some of the observed responses to overlap. In addition, stretches performed in this study were much slower and of greater amplitude than in past experiments. No attempt was thus made to separate the various reflex components. Instead a phasic and a tonic reflex response were evaluated (see Methods). For all knee extensors the size of the phasic reflex increased proportionally with the background torque. This agrees well with the automatic gain principle (Marsden et al. 1976; Matthews 1986) and past studies on the quadriceps (Lamontagne et al. 2000). The tonic response seen when the stretch is held constant is only slightly smaller than the phasic response. The slower velocity and larger amplitude of the stretch applied in this study compared to past studies (Crago et al. 1976; Sinkjaer et al. 1988) may explain why the phasic response is nearly comparable to the tonic response. Lamontagne et al. (2000) suggested that the phasic response is mostly a velocity sensitive group Ia mediated response, whereas the tonic response is largely influenced by group II and Ia length sensitive afferents. The effects of these EMG responses on torque pattern are discussed below. Similar findings have been made for the ankle extensors (Grey et al. 2002; Toft et al. 1991). The automatic gain principle is still upheld for the tonic reflex response.
Non-reflex torque
Previous studies in both animals (Nichols and Houk 1976) and humans (Carter et al. 1990) have demonstrated a significant yield in torque in the absence of reflexes, as soon as the short range stiffness of the muscle was exceeded. This effect was not observed to the same extent in the present results. Rather the non-reflex torque peaked at approximately 60–80 ms following stretch onset then increased further until 200 ms where it remained until just prior to or at cessation of the stretch. It is likely that this may be attributable to the high amplitude and slow velocity of the stretch. The broken cross-bridges may have been able to reform at the new length without any discernible decreases in torque. Carter et al. (1990) reported no yielding in hand muscles when the amplitude of the stretch was 5° or higher and had a rise time of 100 ms or more. Alternatively in this study, the high inertia of the foot and lower leg segment may have masked any obvious yielding effect. Sinkjaer et al. (1988) did not observe yielding in ankle extensors or flexors stretched 5° in 30 ms, and they attributed this to the high inertia of the combined foot and leg segment.
When reflexes are absent, the joint torque is dependent on the mechanical properties of both passive tissue and active muscle fibers. In this study we did not account for these passive and intrinsic components separately. However, the non-reflex torque measured at zero background torque is assumed to reflect the passive torque contribution (Sinkjaer et al. 1993). The intercept of the curves in Fig. 5 panels A and B have values approaching 0 Nm, implying a very small contribution from passive tissues. In addition, the relationship between non-reflex torque and background torque at 300 ms following stretch was highly linear (r=0.9), suggesting that the number of attached cross bridges increases linearly with increasing background torque. It may thus be assumed that at least at 300 ms after stretch onset, the intrinsic torque was dominant at all levels of background torque.
Reflex torque
The reflex torque was obtained by subtracting the total torque produced during a voluntary contraction from the non-reflex torque measured as a result of directly stimulating the nerve innervating the quadriceps. Reflex torque exceeded non-reflex torque 60–80 ms following stretch onset. The reflex response observed in the EMG of the knee extensors appeared approximately 30 ms earlier. This is a reasonable time interval when electromechanical delay is considered. Previous studies have demonstrated that total joint torque is affected by a number of factors. These include both stretch parameters (Allum and Mauritz 1984; Sinkjaer et al. 1988) and the state of the muscle in terms of its length and contractile state prior to stretch (Kearney et al. 1997; Lin and Rymer 1993; Mirbagheri et al. 2000). To eliminate the effects the stretch that parameters may have on the reflex torque we applied stretches of constant amplitude, velocity, and duration. To isolate the effects of muscle length and state we applied the stretches with the knee joint in a fixed position while the precontraction of the extensors was varied.
Only a few studies have examined the torque generated from reflexes at background torques beyond 60% MVC (Sinkjaer et al. 1988). The inability of subjects to maintain high precontraction levels and the necessity of a powerful actuator to apply the stretches are the major limiting factors. In this study background torques of up to 90% were investigated. However, only a few subjects were able to maintain background torques higher than 70% MVC. A slight tremor in the force trace was observed when subjects had difficulties. Small oscillations superimposed on the perturbation result in a significant decrease in the reflex torque (Stein and Kearney 1995). This is synonymous to the vibration reflex. Great care was taken to eliminate such trials. Mirbagheri et al. (2000) also observed variability in the reflex torque between subjects. They calculated stiffness rather than torque, and argued that reflexes in subjects expressing low stiffness may not be of importance. Although variability was high, especially at low background torques, reflex torque remained a significant part of total joint torque. It is difficult to explain the discrepancies in reflex torque magnitude between subjects. It is possible that in this study differing anthropometric characteristics may have resulted in a greater muscle stretch from the same perturbation in some subjects than in others. Also, stronger reflex bursts as seen in the EMG may have allowed a greater reflex torque contribution in some subjects than in others.
The highest contribution of the reflex to the total joint torque was measured for background torques of 20–40% MVC. This confirms reports from cat and human ankle extensors (Hoffer and Andreassen 1981; Mirbagheri et al. 2000; Nichols and Houk 1976; Toft et al. 1991) but not human hand (Carter et al. 1990) and forearm muscles (Thomson and Chapman 1988). Differences in the investigated muscles and their reflex control may reasonably explain the differences between the lower and the upper limb. Across all background torques the average values for reflex torque attained in this study ranged from 16% to 52% of total torque. Past studies have reported similar values for reflex contribution from human ankle flexors (Sinkjaer et al. 1988) and extensors (Kearney et al. 1997; Toft et al. 1991).
It is interesting to note that in confirmation with others (Hoffer and Andreassen 1981; Nichols and Houk 1976; Toft et al. 1991) the reflex EMG recordings (both phasic and tonic) were well correlated with reflex torque at low levels of precontraction but less at the higher levels. Both tonic and phasic EMG reflex responses increase approximately linearly with increases in background torque. However, at background torques higher than 50% MVC the additional muscle activity is unable to enhance reflex torque further. Similar results were reported in cat ankle extensors (Hoffer and Andreassen 1981). Although most motor units are recruited at the higher force levels, further increases in EMG magnitude can be obtained by enhancing their firing frequency. Since at high background torques no additional torque is generated, it stands to reason that all motor units are already firing close to their fusion frequency. Thus there was saturation at the muscular level. Toft et al. (1991) applied a simple model which successfully verified this. At low precontraction levels the effect of the relationship between background torque and muscle activity was able to accurately predict the torque increment. At higher levels the magnitude behavior of a single twitch elicited by supramaximal nerve stimulation had to be included in the model to accurately predict the total torque.
Functional implications
The total torque is comprised of passive, intrinsic and reflex components. During lengthening the intrinsic contribution is limited by the elasticity of the contractile machinery. If this is exceeded, intrinsic torque is unable to compensate for changes in total torque. Reflexes, however, can compensate for this discrepancy. During dynamic tasks such as walking the knee extensor stretch reflex is highly modulated during the progression from the late swing to the early stance phase (Mrachacz et al. 2000). The highest values are present while the knee extensors are contracting to raise the center of gravity upwards in readiness for the following take off phase. While it has been found that much of the ongoing activity in ankle extensors during this phase is attributable to afferent feedback and thus reflex activity (Nielsen and Sinkjaer 2002; Sinkjaer et al. 2000), this has not been demonstrated for the extensors of the knee. Even so, reflexes during the static task presented here can contribute significantly to the total torque produced around the knee joint. It stands to reason that with a high reflex gain during the stance phase of walking that they have the potential to contribute to the joint torque during a more dynamic task, at least in a situation when unexpected perturbations are encountered.
Conclusion
The reflex mediated torque was shown to increase with increasing levels of background torque for all subjects investigated. Depending on the subject the reflex mediated torque contributed on average 16–52% of the total torque. The results presented substantiate that the stretch reflex elicited in the knee extensors contributes to the overall stiffness of the knee joint as the ankle extensors do at the ankle joint following an unexpected stretch (Toft et al. 1991). Functionally it is of importance to maintain the knee joint stiffness as stable as possible throughout a movement, especially as the muscles spanning the knee are very powerful such that any uncontrolled motion could lead to severe trauma. It is concluded that the stretch reflex of the quadriceps muscle has the potential to contribute substantially to the torque produced at the knee joint.
Reference
Allum JH, Mauritz KH (1984) Compensation for intrinsic muscle stiffness by short-latency reflexes in human triceps surae muscles. J Neurophysiol 52:797–818
Allum JH, Mauritz KH, Vogele H (1982) The mechanical effectiveness of short latency reflexes in human triceps surae muscles revealed by ischaemia and vibration. Exp Brain Res 48:153–156
Capaday C (2002) The special nature of human walking and its control Trends Neurosci 25:370–376
Carter RR, Crago PE, Keith MW (1990) Stiffness regulation by reflex action in the normal human hand. J Neurophysiol 64:105–118
Christensen LO, Petersen N, Andersen JB, Sinkjaer T, Nielsen JB (2000) Evidence for transcortical reflex pathways in the lower limb of man. Prog Neurobiol 62:251–272
Crago PE, Houk JC, Hasan Z (1976) Regulatory actions of human stretch reflex J Neurophysiol 39:925–935
Dietz V, Discher M, Faist M, Trippel M (1990) Amplitude modulation of the human quadriceps tendon jerk reflex during gait. Exp Brain Res 82:211–213
Garland SJ, Gerilovsky L, Enoka RM (1994) Association between muscle architecture and quadriceps femoris H-reflex. Muscle Nerve 17:581–592
Grey MJ, Ladouceur M, Andersen JB, Nielsen JB, Sinkjaer T (2001) Group II muscle afferents probably contribute to the medium latency soleus stretch reflex during walking in humans. J Physiol (Lond) 534:925–933
Grey MJ, Larsen B, Sinkjaer T (2002) A task dependent change in the medium latency component of the soleus stretch reflex. Exp Brain Res 145:316–322
Hoffer JA, Andreassen S (1981) Regulation of soleus muscle stiffness in premammillary cats: intrinsic and reflex components. J Neurophysiol 45:267–285
Kearney RE, Stein RB, Parameswaran L (1997) Identification of intrinsic and reflex contributions to human ankle stiffness dynamics. IEEE Trans Biomed Eng 44:493–504
Lamontagne A, Voigt M, Larsen K, Sinkjaer T (2000) Early and late components of the quadriceps stretch reflex in human. Proceedings of the Society for Neuroscience, Annual Meeting 26, New Orleans
Liddell EGT, Sherrington CS (1924) Reflexes in response to stretch-myotatic reflexes. Proc R Soc Lond B Biol Sci 96:212–242
Lin DC, Rymer WZ (1993) Mechanical properties of cat soleus muscle elicited by sequential ramp stretches: implications for control of muscle. J Neurophysiol 70:997–1008
Marsden CD, Merton PA, Morton HB (1976) Servo action in the human thumb. J Neurophysiol 257:1–44
Matthews PB (1986) Observations on the automatic compensation of reflex gain on varying the pre-existing level of motor discharge in man. J Physiol (Lond) 374:73–90
Mirbagheri MM, Barbeau H, Kearney RE (2000) Intrinsic and reflex contributions to human ankle stiffness: variation with activation level and position. Exp Brain Res 135:423–436
Mrachacz N, Lavoie B, Sinkjaer T (2000) Characterization of the human quadriceps stretch reflex during human walking. Proceedings of the Society for Neuroscience, Annual Meeting 26, New Orleans
Nardone A, Tarantola J, Miscio G, Pisano F, Schenone A, Schieppati M (2000) Loss of large-diameter spindle afferent fibres is not detrimental to the control of body sway during upright stance: evidence from neuropathy. Exp Brain Res 135:155–162
Nichols TR, Houk JC (1976) Improvement in linearity and regulation of stiffness that results from actions of stretch reflex. J Neurophysiol 39:119–142
Nielsen JB, Sinkjaer T (2002) Afferent feedback in the control of human gait. J Electromyogr Kinesiol 12:213–217
Schieppati M, Nardone A (1999) Group II spindle afferent fibers in humans: their possible role in the reflex control of stance. Prog Brain Res 123:461–472
Sinkjaer T (1997) Muscle, reflex and central components in the control of the ankle joint in healthy and spastic man. Acta Neurol Scand Suppl 170:1–28
Sinkjaer T, Hayashi R (1989) Regulation of wrist stiffness by the stretch reflex. J Biomech 22:1133–1140
Sinkjaer T, Toft E, Andreassen S, Hornemann BC (1988) Muscle stiffness in human ankle dorsiflexors: intrinsic and reflex components. J Neurophysiol 60:1110–1121
Sinkjaer T, Toft E, Larsen K, Andreassen S (1993) EMG-Torque dynamics at different contraction levels in human ankle muscles. J Electromyogr Kinesiol 3:67–77
Sinkjaer T, Andersen JB, Ladouceur M, Christensen LO, Nielsen JB (2000) Major role for sensory feedback in soleus EMG activity in the stance phase of walking in man. J Physiol (Lond) 523:817–827
Stein RB, Capaday C (1988) The modulation of human reflexes during functional motor tasks. Trends Neurosci 11:328–332
Stein RB, Kearney RE (1995) Nonlinear behavior of muscle reflexes at the human ankle joint. J Neurophysiol 73:65–72
Thomson DB, Chapman AE (1988) The mechanical response of active human muscle during and after stretch. Eur J Appl Physiol Occup Physiol 57:691–697
Toft E, Sinkjaer T, Andreassen S, Larsen K (1991) Mechanical and electromyographic responses to stretch of the human ankle extensors. J Neurophysiol 65:1402–1410
Toft E, Sinkjaer T, Andreassen S, Hansen HJ (1993) Stretch responses to ankle rotation in multiple sclerosis patients with spasticity. Electroencephalogr Clin Neurophysiol 89:311–318
Voigt M, de Zee M, Sinkjaer T (1999) A fast servo-controlled hydraulic device for the study of muscle mechanical and reflex properties in humans. Proceedings of the 17th Congress of the International Society of Biomechanics, Calgary
Wilmink RJH, Huyghues CMJI, Nichols TR (1997) Gastrocnemius muscles provide a major source of interjoint force feedback in the cat hindlimb. Proceedings of the Society for Neuroscience, Annual Meeting 23, New Orleans
Yang JF, Stein RB, James KB (1991) Contribution of peripheral afferents to the activation of the soleus muscle during walking in humans. Exp Brain Res 87:679–687
Zhang LQ, Nuber G, Butler J, Bowen M, Rymer WZ (1998) In vivo human knee joint dynamic properties as functions of muscle contraction and joint position. J Biomech 31:71–76
Acknowledgements
This study was funded by The Danish National Research Foundation. The authors also express their gratitude to Knud Larsen for his technical support.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Mrachacz-Kersting, N., Sinkjaer, T. Reflex and non-reflex torque responses to stretch of the human knee extensors. Exp Brain Res 151, 72–81 (2003). https://doi.org/10.1007/s00221-003-1483-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-003-1483-8