
Abstract
Seasonal variations in the fatty acid compositions of gilthead sea bream (Sparus aurata) and white sea bream (Diplodus sargus), captured in Iskenderun Bay, Eastern Mediterranean of Turkey, were investigated. Results from studying the composition over all seasons showed that the basic saturated, monounsaturated and polyunsaturated fatty acids for gilthead sea bream and white sea bream were palmitic acid (16:0), oleic acid (18:1) and docosahexaenoic acid (DHA, 22:6ω3). The other main fatty acids for both species were myristic acid (14:0), stearic acid (18:0), palmitoleic acid (16:1), linoleic acid (18:2ω6) (especially in autumn and winter for gilthead sea bream), and eicosapentaenoic acid (EPA, 20:5ω3). Gilthead sea bream and white sea bream exhibited seasonal fluctuations in their fatty acid contents. EPA ratios in gilthead sea bream in the autumn, winter, spring and summer were 5.42%, 4.69%, 5.20% and 4.27%, whereas the ratios in white sea bream in autumn, spring and summer were found to be 5.03%, 4.53% and 6.97%, respectively. DHA ratios in gilthead sea bream in autumn, winter, spring and summer were 15.37%, 14.16%, 9.51% and 7.07%, whereas the ratios in white sea bream in autumn, spring and summer were found to be 11.49%, 20.17% and 7.74%, respectively. The present study suggests that the daily consumption of either 100 g of gilthead sea bream captured in any season or 100 g white sea bream captured in spring or summer could meet people’s needs for EPA+DHA fatty acids.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Fatty acids of fish oil are currently being championed as beneficial for human health [1]. Consumption of modest amounts of ω-3 fatty acids is associated with reduced incidence of fatal coronary heart disease and decreased vulnerability to ventricular fibrillation, perhaps because the fish provide an essential amount of long chain ω-3 PUFA [2, 3]. Since these fatty acids only occur in high amounts in seafood, it is necessary to determine the fatty acid compositions and total lipid contents of various seafoods in order to be able to recommend a suitable preventive diet.
Gilthead sea bream (Sparus aurata) and white sea bream (Diplodus sargus), which belong to the same family (Sparidae) are two of the most commercially important demersal fish species in Turkey and many Mediterranean countries. Gilthead and white sea bream are consumed locally and exported as a processed product to Europe. It is known that fish composition, such as lipid, protein and water content, is subject to seasonal variations that can influence its processing properties. Although some papers have been published on the proximate composition and on comparisons of fatty acids in wild and cultured forms of these species [4, 5, 6, 7], little data is available on seasonal changes in the fatty acid compositions of gilthead sea bream and white sea bream. Due to the popularity of these fish in Turkey and Europe, it its important to establish the seasonal variations in fatty acid composition in these fish. This information, besides being important when recommending a suitable diet for consumers, is useful for the fish processing industry when they need to store the fish for long periods.
Materials and methods
Sample preparation
Gilthead sea bream (112.72±5.57 g) and white sea bream (126.55±8.40 g) were captured with longlines in Iskenderun Bay, on the eastern Mediterranean coast of Turkey in February, April, July and November 2002. The samples were kept in ice after capture and transferred to the laboratory. The fish were immediately beheaded, eviscerated and filleted. Chemical composition analyses were performed on these fresh samples, and these analyses were performed in triplicate at the very least.
Proximate analyses
Moisture content and crude ash were determined in an oven at 103 °C and 550 °C respectively until the weight became constant. Total crude protein was determined by Kjeldahl’s method [8], and lipid content was analyzed according to the procedure of Bligh and Dyer [9].
Fatty acids
The lipids were saponified and esterified for fatty acid analysis by the method of Metcalfe et al. [10]. The fatty acid methyl esters (FAMEs) were separated and quantified with a Hewlett-Packard 5880gas-liquid chromatograph (GC) using a capillary column equipped with a flame ionization detector (FID). Separation was achieved on a 50 m×0.20 mm i.d. wall-coated open tubular fused silica capillary column coated with Carbowax 20 M. The column, injector, and detector temperatures were maintained at 200 and 300 °C, respectively. The carrier gas was helium with a split ratio of 1:100. Identification was made by comparison to retention times of authentic standards, argentation TLC followed by GC of the bands separated by degree of unsaturation, and mass spectrometry. The fatty acids were calculated by percentage. For conversion of the percentile values to units of weight, the formulae recommended by Paul and Southgate were employed [11].
Statistical analysis
All data obtained separately for each sampling season were subjected to analysis of variance (one-way ANOVA), at 5% confidence level using the Duncan [12] multiple range test.
Results and discussion
Proximate composition
The proximate compositions of gilthead sea bream and white sea bream over four seasons are shown in Table 1. Mean proximate values of gilthead sea bream found in the study were similar to the findings of Çaklı [7], Alaşalvar et al. [5], and Grigorakis et al. [4]. There is no report on the proximate composition of white sea bream. Mean proximate values of white sea bream were coincident with other species of the same family, as reviewed by the Torry Research Station [13].
The results obtained from this study indicate that the moisture content in gilthead sea bream fillets in winter was significantly higher than in the other seasons (p<0.05). Similarly, the moisture content in white sea bream fillets in autumn was the highest (p<0.05). Depending on the diet of the fish in summer when nutrients are more abundant, an increase in the proportion of dry fish matter is observed, while there is a decrease in the moisture content accordingly. While no significant seasonal changes were observed in gilthead sea bream fillets in terms of crude ash and crude protein (p>0.05), it was found out that the content of crude ash and crude protein in white sea bream fillets in autumn differed greatly compared with the other seasons (p<0.05). This significant increase in the protein content of the white sea bream fillets is thought to be in proportion with the decrease in the lipid content.
As shown in Table 1, the lipid contents of both gilthead sea bream and white sea bream fillets in spring and summer are significantly higher than those in autumn and winter (p<0.05). Grigorakis et al. [4] reported that the gilthead sea bream does not change its diet during winter, but its feeding levels do decrease. Therefore, this decrease in the feeding level may have caused the lipid content of the gilthead sea bream to decrease. Numerous researchers have pointed out that seasonal changes in the lipid contents of various species particularly depend on the abundance of fish food [14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24].
Fatty acid compositions
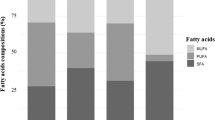
The fatty acid compositions of gilthead sea bream and white sea bream captured in different seasons are given in Tables 2 and 3 respectively. It was determined that the major fatty acids in both gilthead sea bream fillets and white sea bream fillets throughout all of the seasons were palmitic acid (16:0), myristic acid (14:0), stearic acid (18:0), palmitoleic acid (16:1), oleic acid (18:1), linoleic acid (18:2ω6) (especially in autumn and winter for gilthead sea bream), eicosapentaenoic acid (EPA, 20:5ω3) and docosahexaenoic acid (DHA, 22:6ω3). Grigorakis et al. [4] also determined that these fatty acids constituted the basic components of the fatty acids of gilthead sea bream fillets. On the other hand, Imre and Saglık [6] reported that palmitic acid, palmitoleic acid, stearic acid and oleic acid constituted the basic fatty acids of white sea bream. Similarly, Varljen et al. [25] found that the basic fatty acids of Diplodus vulgaris, belonging to the same genus as white sea bream, were palmitic acid, palmitoleic acid, stearic acid, oleic acid, arachidonic acid, eicosapentaenoic acid and docosahexaenoic acid.
The levels of palmitic acid versus the total fatty acids of the gilthead sea bream fillets in autumn, winter, spring and summer were identified to be 16.87%, 15.73%, 20.82% and 16.48%, respectively. These levels were observed to be 16.39%, 17.25% and 18.48% in white sea bream fillets in autumn, spring and summer, respectively. Similarly, oleic acid was the basic monounsaturated fatty acid (MUFA) in both species. The seasonal changes observed in the levels of oleic acid in both species were also recorded for Baltic herring (Clupea harengus membras) by Aro et al. [26]. For this reason, palmitic acid and oleic acid could, respectively, be referred to as the basic saturated and monounsaturated fatty acids. Similar results were reported for gilthead sea bream by Çaklı [27] and Grigorakis et al. [4], for white sea bream by Imre and Sağlık [6] and for other species by numerous researchers [18, 26, 28, 29, 30]. In our study, the lowest total saturated fatty acid (∑SFA) level was found in winter for gilthead sea bream (Table 2). The decrease in the level of ∑SFA in winter for gilthead sea bream is thought to be due to the catabolization of saturated fatty acids to compensate for the extra metabolic energy required in that period.
Ackman [19] reported that EPA and DHA of the ω-3 series fatty acids were the dominant polyunsaturated fatty acids of fish oil. In our study, DHA was the basic polyunsaturated fatty acid for both species. Numerous researchers have reported that DHA constitutes a majority of the PUFA in most marine fish [4, 5, 25, 30, 31]. For gilthead sea bream, DHA levels in autumn, winter, spring and summer were determined as 15.37%, 14.16%, 9.51% and 7.07%, respectively. For white sea bream, DHA levels in autumn, spring and summer were 11.49%, 20.17% and 7.74%, respectively. EPA levels of gilthead sea bream were found to be 5.42%, 4.69%, 5.20% and 4.27% in autumn, winter, spring and summer, respectively. For white sea bream, EPA levels were found to be 5.03% for autumn, 4.53% for spring and 6.97% for summer. No increase was observed in the ratios of EPA and DHA (Table 2) parallel to the increase in lipid content in gilthead sea bream in spring and summer. This result indicates that the saturated fatty acids rather than the ω3 series fatty acids led to the increase in the gilthead sea bream fillet lipid content in spring and summer, since the levels of ω3 series fatty acids decline from 19.81% in winter to 15.32% and 12.08% in spring and summer, while those of saturated fatty acids were 32.72% and 32.17% (Table 2). Love [32] reported that the water temperature had a significant influence on the polyunsaturated fatty acid levels of lipids, in that when the polyunsaturated fatty acid levels of the phospholipids, an important component of cell membranes, increased, the melting point of the lipid mixture decreased, and therefore the flexibility and permeability of the cells increased. In the present study, the DHA level (7.07%) decreased significantly (p<0.05) in summer when the temperature was high (Table 2). The same results were recorded for white sea bream, where the DHA level (7.74%) dropped significantly (p<0.05) (Table 3). Furthermore, gamma linolenic acid (GLA, 18:3ω6), which is essential for humans and is converted from linoleic acid in a rather slow process, and which is mostly found in nutrients in low levels, was found to be in low levels in both species, while displaying a significant increase (p<0.05) in summer (Tables 2 and 3). Parallel to the data obtained from this study, a number of other researchers have recorded that the amounts and types of fatty acids in tissues of other fish species were affected by seasonal conditions [18, 26, 28, 31, 33, 34, 35]. However, various other factors, such as the size and age of the fish, the maturity period, and the geographical location also affect the fatty acid content and composition of fish muscle [36, 37].
Epidemiological data indicated that there was a decrease in the number of deaths caused by coronary heart diseases in people who consumed fish or fish oil containingsmall amounts (0.4 g) of ω3 series fatty acid on a regularly daily basis [2]. The British Nutrition Foundation has recommended that for a balanced and healthy diet, we should all consume 0.2 g of EPA+DHA daily or 1.5 g on a weekly basis. Our data shows that the consumption of 100 g of gilthead sea bream daily in all seasons could meet this demand (Table 4). It was also determined that the daily consumption of 100 g of white sea bream in spring and summer can also meet this demand, but this fish should be consumed in greater amounts in autumn (Table 4). The seasonal changes observed in the total saturated, monounsaturated, and unsaturated fatty acid levels provide researchers with valuable information for preparing diet tables. Additional studies on the fatty acid composition of the other popular species in Mediterranean are also needed.
References
British Nutrition Foundation (1992) Unsaturated fatty acids. Nutritional and physiological significance (Report of the British Nutrition Foundation). Chapman and Hall, London, pp 156–157
Kromhout D, Bosschieter EB, Coulander C (1985) New Engl J Med 312:1205–1209
Siscovick DS, Raghunathan TE, King I, Weinmann S, Wicklund KG, Albright J, Bovjerg V, Arbogast P, Smith H, Kushi LH, Cobb LA, Copass MK, Psaty BM, Lemaitre R, Retzlaff B, Childs M, Knopp RH (1995) JAMA–J Am Med Assoc 274:1363–1367
Grigorakis K, Alexis MN, Taylor KDA, Hole M (2002) Int J Food Sci Tech 37:477–484
Alaşalvar C, Taylor KDA, Shahidi F (2002) J Agr Food Chem 50(7):2039–2045
Imre S, Sağlik S (1998) Turk J Chem 22:321–324
Çaklı Ş (1996) Gıda Teknolojisi Dergisi 21:243–250
AOAC (1984) Official methods of analysis, 14th edn. Association of Official Analytical Chemists, Washington, DC
Bligh EG, Dyer WJ (1959) Can J Biochem Physiol 37:911–917
Metcalfe LD, Schmitz AA, Pelka JR (1966) Anal Chem 38:514
Paul AA, Southgate DAT (1988) In: McCance A, Widdowsons F (eds) The composition of foods. HMSO, London, pp 16–17
Duncan DB (1955) Biometrics 11:1–42
Torry Research Station (1989) Yield and nutritional value of the commercially more important fish species (FAO Fisheries Technical Paper No 309). FAO, Rome, p 187
Craig S, Mackenzie DS, Jones G, Gatlin DM III (2000) Aquaculture 190:89–102
Robards DM, Anthony JA, Rose GA, Piatt JF (1999) J Exp Mar Biol Ecol 242:245–258
Gökoğlu N, Özden Ö, Erkan N, Baygar T, Metin S (1999) Int J Food Sci Tech 34:401–402
Bandarra NM, Batista I, Nunes ML, Empis JM, Christine WW (1997) J Food Sci 62(1):40–42
Mendez E, Gonzalez RM (1997) Food Chem 59(2):213–217
Ackman RG (1995) Composition and nutritive value of fish and shellfish lipids. In: Ruither A (ed) Fish and fishery products. CAB International, Oxford, UK, pp 117–156
Sargent JR (1995). Origins and functions of egg lipids: nutritional implications. In: Bromage NR, Roberts RJ (eds) Broodstock management and egg and larval quality. Blackwell, Oxford, UK, pp 353–372
Leonor Nunes M, Cardinal M, Mendes R, Morao Campos R, Bandarra NM, Lourenço H, Jerome M (1992) Effect of season and storage on proteins and lipids of sardine (Sardinae pilchardus) minces and surimi. In: Huss HH et al (eds) Quality assurance in the fish industry. Elsevier, Amsterdam, pp 73–79
Kolakowska A, Kwiatkowska L, Lachowicz K, Gajowiecki Bortnowska G (1990) Effect of fishing season on frozen storage quality of Baltic herring. Int Conf on Advancement in Seafood Science and Technology (Seafood 2000), Halifax, Canada, 13–16 May 1990
Krzynowek J (1985) Food Technol 39:61–68
Henderson RJ, Sargent JR, Hopkins CCE (1984) Mar Biol 78:255–263
Varljen J, Sulic S, Brmalj J, Baticic L, Obersnel V, Kapovic M (2003) Food Technol Biotech 41(2):149–156
Aro T, Tahvonen R, Mattila T, Nurmi J, Sivonen T, Kallio H (2000) J Agr Food Chem 48:6085–6093
Çaklı Ş (1994) PhD Thesis. Department of Fisheries and Processing Technology, Institute of Natural and Applied Science, Ege University, Turkey, p 71
Luzia LA, Sampaio GR, Castellucci CMN, Torres EAFS (2003) Food Chem 83:93–97
Grün IU, Shi H, Fernando LN, Clarke AD, Ellersieck MR, Beffa DA (1999) Lebensm Wiss Technol 32:305–311
Tanakol R, Yazıcı Z, Şener E, Sencer E (1999) Lipids 34(3):291–297
Orban E, Lena G Di, Nevigato T, Casini I, Santaroni G, Marzetti A, Caproni R, (2002) J Food Sci 67(2):542–546
Love RM (1992) Biochemical dynamics and the quality of fresh and frozen fish. In: Hall GM (ed) Fish processing technology. Chapman and Hall, London, pp 1–31
Kolakowska A, Czerniejewska-Surma B, Gajowiecki L, Lachowicz K, Zienkowicz L (1992) Effect of fishing season on shelf life of iced Baltic herring. In: Huss HH et al (ed) Quality assurance in the fish industry. Elsevier, Amsterdam, pp 81–91
Gamez-Meza N, Higuera-Ciapara L, Calderon De La Barca AM, Vazquez-Moreno L, Noriega-Rodriguez J, Angulo-Guerrero O (1999) Lipids 34(6):639–642
Bandarra N, Batista I, Nunes ML, Empis JM (2001) Eur Food Res Technol 212(5):535–539
Ackman RG (1989) Prog Food Nutr Sci 13:161–241
Kiessling A, Pickova J, Johansson L, Asgard T, Storebakken T, Kiessling KH (2001) Food Chem 73:271–284
Acknowledgements
The authors are grateful to the Scientific Research Project Fund of Cukurova University for their financial support of this research project.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Özyurt, G., Polat, A. & Özkütük, S. Seasonal changes in the fatty acids of gilthead sea bream (Sparus aurata) and white sea bream (Diplodus sargus) captured in Iskenderun Bay, eastern Mediterranean coast of Turkey. Eur Food Res Technol 220, 120–124 (2005). https://doi.org/10.1007/s00217-004-1060-9
Received:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00217-004-1060-9