
Abstract
Rationale
Intermittent Access (IntA) cocaine self-administration, which models intermittent patterns of cocaine use in humans during the transition to addiction, is especially effective in producing incentive-sensitization and other addiction-like behavior in male rats. However, female rats show more robust psychomotor sensitization than males, and following initial use, women develop problematic patterns of drug use more readily than men. We hypothesized, therefore, that female rats might be more susceptible to the incentive-sensitization produced by IntA experience.
Objective
To assess changes in motivation for cocaine, using a behavioral economic indicator of cocaine demand (“elasticity” of demand curves), and other addiction-like behavior, as a function of IntA cocaine self-administration experience in male and female rats.
Results
IntA experience produced a progressive increase in motivation for cocaine in both males and females, as indicated by a decrease in the elasticity of cocaine demand curves, and this persisted undiminished following 14 days of abstinence. However, IntA produced a more rapid and greater increase in motivation for cocaine (incentive-sensitization) in females than males. Females also consumed more cocaine than males, although this did not predict changes in motivation. On the other hand, there were no sex differences in the preferred level of cocaine consumption when cost was low (Q0), nor in cocaine- or cue-induced reinstatement of drug-seeking behavior.
Conclusions
The observation that females are more susceptible to incentive-sensitization when intermittently exposed to cocaine may provide a mechanism for the more rapid development of problematic drug use in females (“telescoping effect”) reported in clinical studies.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Women are reported to transition from initial casual or recreational patterns of drug use to the problematic patterns of use that define addiction more readily than men (for reviews, see Fattore et al. 2008; Becker 2016; Becker and Koob 2016; Lynch 2018). For example, from the time of initial use, women tend to seek treatment sooner than men, across a number of drug classes (Anglin et al. 1987a; Griffin et al. 1989; Brady and Randall 1999), and when they do seek treatment, they present with more severe symptoms (Anglin et al. 1987b; Kosten et al. 1993). In addition, women report higher levels of craving induced by the presentation of cocaine-associated cues, more frequent cravings, greater difficulty in controlling their cocaine use, and often use more cocaine than intended (Robbins et al. 1999; Elman et al. 2001; Kennedy et al. 2013). Although men and women do not differ in the amount of cocaine they report using per use event, women provide fewer cocaine-negative samples than men, suggesting they use cocaine more frequently (Kennedy et al. 2013). Taken together, these findings have been characterized as reflecting a “telescoping effect” in women—that is, they progress from casual drug use to addiction faster than men and have a smaller window for medical intervention and treatment (Piazza et al. 1989; Brady and Randall 1999).
Related findings have been reported in preclinical self-administration studies in rats. Perhaps mirroring the “telescoping effect” in humans, female rats are more likely to acquire cocaine self-administration (Lynch and Carroll 1999; Hu et al. 2004; see discussion for more details). In addition, after limited drug experience, female rats are more motivated to self-administer cocaine (Roberts et al. 1989; Cummings et al. 2011), as assessed using a progressive-ratio test (however, also see Lynch and Taylor 2004). In addition, when given extended access to cocaine using a 24-h access self-administration procedure, females self-administer more drug than males (Lynch and Taylor 2004; Lynch et al. 2005) and the same is true when male and female rats are given access to cocaine using the Long Access (LgA) self-administration procedure (Roth and Carroll 2004; Smith et al. 2011).
It is still unclear, however, why women may transition to addiction more readily than men. One possibility is suggested by the preclinical literature on behavioral sensitization produced by intermittent exposure to a number of drugs of abuse (Robinson and Becker 1986). Behavioral sensitization refers to a persistent increase in the ability of a drug to induce a number of drug effects as a result of past drug experience, and sensitization-related neuroadaptations are hypothesized to play a critical role in the development of addiction (Robinson and Berridge 1993; Vezina 2004; Berridge and Robinson 2016). It is relevant, therefore, that female rats show more robust psychomotor sensitization than males following repeated, intermittent exposure to experimenter-administered psychomotor stimulant drugs (Robinson 1984; Robinson and Becker 1986; van Haaren and Meyer 1991).
Although females show greater psychomotor sensitization than males, the degree to which there are sex differences in the incentive-sensitization produced by cocaine self-administration is not well studied. Following 24-h cocaine self-administration access, results are mixed as to whether motivation is increased more as a function of cocaine experience in female than male rats (Lynch and Taylor 2004; but see Ramôa et al. 2013, 2014), and in studies that examined sex differences during LgA self-administration, tests for motivation were not conducted after self-administration experience.
The purpose of the present study was to determine whether male and female rats differ in the extent to which they are susceptible to incentive-sensitization following cocaine self-administration. To do this, we used a recently developed Intermittent Access (IntA) cocaine self-administration procedure (Zimmer et al. 2012; Kawa et al. 2016; Allain et al. 2017). Relative to the widely used Long Access (LgA) self-administration procedure (Ahmed and Koob 1998; Ahmed 2012), IntA is thought to better reflect patterns of cocaine use in humans and is particularly effective at producing incentive-sensitization and addiction-like behavior, despite producing far less total drug intake than LgA (see Zimmer et al. 2012; Kawa et al. 2016; Allain et al. 2017; James et al. 2018; Allain et al. 2015 for review). In addition, in male rats, the degree of psychomotor sensitization seen during IntA self-administration predicts subsequent motivation for cocaine (Allain et al. 2017). IntA self-administration has not yet been studied in females, and so, we compared the degree of incentive-sensitization produced by IntA experience in male and female rats, assessed using a behavioral economic indicator of cocaine demand (Bentzley et al. 2013; Kawa et al. 2016). We hypothesized that, as with psychomotor sensitization, IntA experience would produce a more rapid and greater increase in cocaine demand (incentive-sensitization) in female than male rats. The results support this hypothesis.
Materials and methods
A total of 52 (28 males; 24 females) Sprague-Dawley rats (Envigo, Haslett, MI) weighing 250–275 g on arrival were housed individually on a reverse 12-h light/12-h dark cycle (lights on at 20:00) in a climate-controlled colony room. All rats were 9–14 weeks old upon arrival (females tended to be slightly older), and all rats were beyond adolescence by the time the experiment began (Ojeda et al. 1980; Sisk and Zehr 2005). Males and females were housed in separate but identical housing rooms. All testing was conducted during the 12-h lights off period. After arrival, rats were given 1 week to acclimate to the colony room before testing began. Water and food were available ad libitum throughout the experiment. The rats were weighed regularly throughout the experiment, and once/week, the concentration of each rat’s cocaine solution was adjusted so that the self-administration dose was accurate for the current body weight. All procedures were approved by the University of Michigan Committee on the Use and Care of Animals (UCUCA).
Apparatus
Behavioral testing was conducted in standard (22 × 18 × 13 cm) test chambers (Med Associates, St Albans, VT, USA) located inside sound-attenuating cabinets. A ventilating fan masked background noise. Within the test chambers, two nose poke ports were located 3 cm above the floor on the left and right side of the wall, and one port was designated active and the other inactive (side counter-balanced across chambers). A red house light was located at the top, center of the wall opposite the nose ports. During self-administration portions of the experiment, a nose poke into the active port was detected by an infrared photo beam inside the hole and resulted in an intravenous cocaine infusion, delivered by a pump mounted outside the sound attenuating cabinet, through a tube connected to the rat’s catheter back port. The infusion tube was suspended into the chamber via a swivel mechanism, allowing the rat free movement. All measures were recorded using Med Associates software.
Intravenous catheter surgery
Male and female rats underwent intravenous catheter surgery as described previously (Crombag et al. 2000). Briefly, rats were anesthetized using ketamine hydrochloride (90 mg/kg i.p.) and xylazine (10 mg/kg i.p.), and a catheter was inserted into the right jugular vein, and tubing was run subcutaneously to a port located on the rat’s back. During recovery from surgery, rats were administered the analgesic carprofen (5 mg/kg s.c.). Following surgery, catheters were flushed daily with 0.2 ml sterile saline containing 5 mg/ml gentamicin sulfate (Vedco, MO). Catheter patency was tested periodically with intravenous injection of 0.1 ml methohexital sodium (10 mg/ml in sterile water, JHP Pharmaceuticals). If a rat did not become ataxic within 10 s of the injection, the catheter was considered not patent and the rat was removed from the study.
Self-administration: acquisition
Rats were given ~ 7 days to recover from the catheter surgery, and then, self-administration training commenced (see Fig. 1). The rats were placed into the chamber with the house light illuminated, and extinguishing the house light signaled the beginning of each session. At that time, a nose poke into the active port resulted in an intravenous infusion of cocaine hydrochloride (NIDA) dissolved in 0.9% sterile saline (0.4 mg/kg/infusion in 50 μl delivered over 2.6 s) on a fixed ratio-1 (FR-1) schedule. Each infusion was paired with the illumination of a cue light in the active nose port for 20 s. Nose pokes during this 20 s period were recorded but had no consequences. An inactive nose port was also present at all times, and pokes there were recorded but had no consequences. To ensure that during initial training all rats received the same amount of drug and cue exposure, an infusion criteria (IC) procedure was imposed on self-administration sessions, as described previously (Saunders and Robinson 2010). During these sessions, session length was determined by how long it took each rat to reach the predetermined number of infusions, not by an explicit time limit. Each rat had 3 sessions at IC10 and 5 sessions at IC20. A total of 4 rats (3 males, 1 female) were excluded during acquisition training because they failed to reach the infusion criteria or failed to discriminate between the active nose port and the inactive nose port.

A flow diagram illustrating the overall experimental design and timeline. Each “probe” was a 2-day probe test using the within-session threshold procedure to assess cocaine demand. IntA, Intermittent Access; Abst, Abstinence; Rein, Reinstatement
Self-administration: within-session threshold procedure
The day after the final acquisition session, rats were trained on a within-session threshold procedure, as described previously (Oleson and Roberts 2009; Oleson et al. 2011; Bentzley et al. 2013; Kawa et al. 2016). Briefly, each session (1/day) was 110 min in length, FR-1 throughout, and every 10 min the dose of drug was decreased on a quarter logarithmic scale (1.28, 0.72, 0.40, 0.23, 0.13, 0.072, 0.040, 0.023, 0.013, 0.007, 0.004 mg/kg/infusion). During the threshold procedure, the nose port cue light was illuminated for the duration of each infusion. Importantly, there was no timeout period following each infusion. As during acquisition, the house light was illuminated when the rats were placed in the chambers and extinguishing the house light signaled the beginning of each session.
Demand curve fitting
Demand curves were generated from the threshold procedure using a focused-fitting approach, as described previously (Bentzley et al. 2013; Kawa et al. 2016). Briefly, each animal’s brain cocaine concentration was calculated to determine relative stability during a session. Demand data points that failed to meet stability criteria were truncated before demand curves were fit by standard techniques (Bentzley et al. 2013). This typically resulted in elimination of the data point from the first 10-min bin, during which the subject “loaded” on cocaine (Oleson et al. 2011), and elimination of all data points that occurred more than 20 min (two data points) after Pmax, when the brain cocaine concentration had dropped significantly (Bentzley et al. 2013). Using this focused-fit approach, the values alpha (α) and Q0 in the exponential demand equation (Hursh and Silberberg 2008) were manipulated to minimize the residual sum of squares, i.e., the square of the difference between the logarithm of the experimentally measured demand and the logarithm of the demand predicted by the exponential demand equation (ln Q = ln Q0 + k(e−αQ0C − 1)) was found for each price and then summed across all prices. Demand curves were fit using an Excel macro that was developed and optimized for use with the within-session threshold procedure (see Bentzley et al. 2013 for details).
The two metrics obtained from this procedure reported here are Q0 and α. Q0 is the preferred level of cocaine consumption when no effort is required; that is, an inherent extrapolation of the animal’s consumption at very low prices (Hursh and Silberberg 2008; Oleson et al. 2011; Bentzley et al. 2013). Q0 can be calculated several ways, but here, it is an estimate of what consumption would be if cost were truly zero. α is a measure of normalized demand elasticity and is equivalent to the slope of the demand curve (Hursh and Silberberg 2008; Bentzley et al. 2013). We report α because it is a uniquely unambiguous measure of motivation that is inherently normalized with respect to Q0. Thus, changes in motivation that are accompanied by changes in Q0 (Bentzley et al. 2014) can be determined with greater confidence than by Pmax (another behavioral economic metric not reported here), or breakpoint on a progressive ratio schedule (Hursh and Silberberg 2008; Bentzley et al. 2013, 2014; James et al. 2018). Motivation is inversely proportional to α, meaning a lower α value corresponds to greater motivation (i.e., less elasticity and higher motivation).
For the initial (baseline) test, each rat was tested daily using the threshold procedure for a minimum of four sessions and until it produced three consecutive sessions with less than ± 25% variation in α. For baseline data analysis, α and Q0 values were averaged over these last 3 sessions for each rat. Each probe test that followed the baseline test consisted of testing each rat for two days using the threshold procedure. Data (not shown) from other experiments have shown that after initial training the rats no longer require multiple days for their behavior to stabilize. The same procedures were used in males and females for fitting demand curves as there are no sex differences in the brain’s absorption of cocaine or the resulting brain cocaine concentration (Bowman et al. 1999). For probe test data analysis, α and Q0 values were averaged over the 2 sessions for each rat. A total of 7 rats (2 males, 5 females) were excluded during the baseline threshold procedure because their behavior failed to stabilize or their catheters failed.
Self-administration: IntA procedure
After completion of the baseline threshold test, the rats were allowed to continue to self-administer cocaine, but now using an intermittent access (IntA) procedure, similar to that described previously (Zimmer et al. 2012; Kawa et al. 2016). Briefly, the rats were placed into the chamber with the house light illuminated. The beginning of the first 5-min Drug Available period started 2 min after the rats were placed into the chamber and was signaled by extinguishing the house light. During the Drug Available period, a nose poke into the active port resulted in an intravenous infusion of cocaine hydrochloride (NIDA) dissolved in 0.9% sterile saline (0.4 mg/kg/infusion in 50 μl delivered over 2.6 s) on a FR-1 schedule. Each infusion was paired with the illumination of a cue light in the nose port for the duration of the infusion. Pokes that were made during the 2.6-s infusion period were recorded but not additionally reinforced. It is important to note that there was no timeout period following the infusion, so the rats could earn another infusion as soon as the preceding infusion ended. After the 5-min Drug Available period, the house light turned on and signaled a 25-min No-Drug Available period. During the No-Drug Available period, nose pokes were recorded but had no consequences. After 25 min, the house light was extinguished and another 5-min Drug Available period began. An inactive port was also present at all times, and pokes there had no consequences.
In this study, we adjusted the traditional IntA procedure in an attempt to make it more intermittent and less predictable. To this end, two of the No-Drug Available periods each day were 50 min long instead of 25 min. The two periods that were extended were randomly determined each day. Each IntA session consisted of 8 Drug-Available and 8 No-Drug Available periods (six 25 min and two 50 min), resulting in a 4-h and 50-min session.
Each rat underwent one IntA session/day for an average of 5 days/week. We varied the number and pattern of days off each week to accentuate the intermittency—for example, on one week, rats may have had only 1 day off and then on the next week, the rats may have had 3 days off. However, rats were never given the day off directly before a probe test. The rats were given a total of 30 IntA sessions and underwent probe tests, using the threshold procedure, after the 10th and 30th IntA sessions and then again after a 14-day abstinence period (see Fig. 1). A total of 41 rats began IntA testing, but 7 (5 males, 2 females) lost catheter patency before the reinstatement tests, and therefore, the N is lower for those later tests. In all, the rats self-administered cocaine for a total of approximately 50 days, combining acquisition (mean of 7 days), four threshold tests (mean of 11 days), and IntA (30 days).
Cocaine-induced reinstatement test
Following the final probe test (after the 14-day abstinence period), rats were tested for cocaine-induced reinstatement using procedures similar to those described previously (Deroche et al. 1999; Kawa et al. 2016). On the first day of this two-day test, rats were placed in the self-administration chambers with the house light illuminated. When the session started 2 min later, the house light was extinguished. All nose pokes during this test were recorded but had no consequences (that is, neither drug nor cue was presented). After a 90-min extinction period, the rats received four IV saline infusions (25, 50, 100, 200 μl), each separated by 30 min. On the following day, the rats were tested using the same procedure, except the saline was replaced by a cocaine solution (0.2, 0.4, 0.8, 1.6 mg/kg).
Extinction and cue-induced reinstatement test
After the cocaine-induced reinstatement test, the rats underwent two-hour extinction sessions (1/day) for at least 5 days and until they made less than 20 active nose pokes for two consecutive sessions. The rats were placed into the chamber with the house light on and the session started 2 min later. Upon the start of the session, the house light turned off and remained off for the duration of the session. Responses into the nose ports during these sessions were recorded but had no consequences. On the day after a rat met the extinction criterion, it underwent a day of testing identical to extinction, except on this day, pokes in the active port were reinforced by illumination of the cue light for 2.6 s.
Statistical analysis
Linear mixed-models analyses were used for all repeated measures data. The best-fitting model of repeated measures covariance was determined by the lower Akaike information criterion score (West et al. 2007). Depending on the model selected, the degrees of freedom may have been adjusted to a non-integer value. Data for the α measure was not normally distributed, and therefore, all statistical tests involving α were made using log-transformed data, consistent with previous studies (Bentzley et al. 2014). Planned post-hoc contrasts (and Bonferroni corrections) were done to compare data between the two sexes from the baseline threshold test. In addition, data from the first three probe tests were analyzed together to test the effects of increasing IntA experience. The fourth probe test was analyzed separately and compared only to the third probe test to test the effect of an abstinence period on motivation following IntA. Statistical significance was set at p < 0.05.
Results
There was no sex difference in the acquisition of cocaine self-administration
Rats were first trained to nose poke for cocaine. Due to the nature of the infusion criteria procedure, differences in the acquisition of self-administration would appear in the number of responses made or the length of each session. The number of responses at the active nose port increased across sessions (as the infusion criteria increased) (effect of IC, F(1, 46) = 18.9, p < 0.001; Fig. 2a), but sex had no effect on the number of active pokes (F(1, 46) = 0.82, p = 0.37). Further, despite the increasing number of infusions, the rats finished their sessions faster with increasing experience (effect of IC, F(1, 46) = 8.27, p = 0.006; Fig. 2b), but sex had no effect on session length (F(1, 46) = 0.23, p = 0.636). Responding at the inactive nose port was low in all rats, and neither sex (F(1, 46) = 0.03, p = 0.87) nor IC (F(1, 45) = 3.85, p = 0.056; data not shown) had any effect.

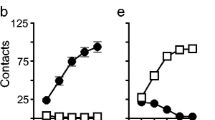
Acquisition and baseline demand. Acquisition of cocaine self-administration behavior using an Infusion Criterion procedure (see the “Materials and methods” section) and baseline demand for cocaine prior to IntA experience. There were no differences between males (n = 25) and females (n = 23) in the acquisition of self-administration as indicated by active nose pokes (a) or the time to meet each criterion number of injections (b). Baseline demand (prior to IntA experience) did not differ between males (n = 23) and females (n = 18) indicated by α (c) or Q0 (d). Values represent means ± SEMs
There was no sex difference in cocaine demand after limited cocaine experience
After limited cocaine self-administration experience, females have been reported to be more motivated to self-administer cocaine than males (Roberts et al. 1989). Therefore, we separately analyzed data from the baseline threshold test to determine if there were sex differences after limited cocaine experience (the acquisition period), prior to IntA experience. Males and females did not differ in the number of sessions required to stabilize on the threshold procedure. Females showed a trend toward a lower α (greater motivation) than males (F(1, 87) = 3.32, p = 0.072; Fig. 2c), and females and males did not differ in their preferred level of consumption when cost was nil (Q0) (F(1, 28) = 0.02, p = 0.885; Fig. 2d). A similar non-significant trend was obtained from statistical analysis of the behavioral economic metric Pmax (data not shown).
There was no sex difference in learning to discriminate drug available and no-drug available periods during IntA self-administration
Following the baseline threshold test, all rats underwent 30 IntA self-administration sessions. All rats quickly learned to discriminate between the alternating Drug Available and No-Drug periods, which were signaled by off/on cycles of the house light, respectively (Fig. 3a,b). In addition, both males and females made more active nose pokes during the Drug Available period with increasing IntA experience, indicating an escalation of responding (effect of session, F(29, 98.9) = 5.03, p < 0.001; Fig. 3c). Males and females did not differ in the number of active nose pokes made during the Drug Available period (F(1, 10.6) = 1.4, p = 0.262). Also, both males and females decreased the number of nose pokes they made at the active nose port during the No-Drug periods of IntA sessions (effect of session, F(29, 79) = 2.84, p < 0.001; Fig. 3d). However, females made more nose pokes at the active port during the No-Drug period than males (F(1, 23.8) = 10.0, p = 0.004). After the first 5 IntA sessions, the majority of No-Drug responding came in the last 5 min of the 25-min period (data not shown), as we have reported previously (Kawa et al. 2016), suggesting persistent, anticipatory responding. It is important to note that the number of inactive nose pokes during the Drug Available and No-Drug periods did not increase with increasing IntA experience nor was there a difference between males and females in the number of inactive responses made during either the Drug Available or No-Drug periods (all p values > 0.1; Fig. 3c,d).

Self-administration behavior during IntA. Both males and females quickly learned to discriminate between the Drug Available periods and No-Drug Available periods. As expected, rats did not discriminate these periods on the first day of IntA testing (n = 23 males, 18 females) (a), but after five sessions, both males and females showed good discrimination (b). Across all 30 IntA sessions, both males and females made progressively more responses during the Drug Available periods as a function of increasing experience (c) and progressively fewer responses during the No-Drug periods, although females made more responses than males during the No-Drug period throughout testing (d). In panels (c) and (d), each point represents the average of three consecutive IntA sessions and arrowheads mark when Probe Tests were conducted (session 30: n = 18 males, 16 females). Values represent means ± SEMs
Females self-administered more cocaine than males during IntA and displayed a different pattern of intake
When analyzed collectively, both male and female rats consumed more cocaine (escalated intake) as a function of increasing IntA experience (effect of session, F(29, 128.2) = 5.23, p < 0.001; Fig. 4a). When analyzed separately, both males (F(29, 93.4) = 2.28, p = 0.002) and females (F(29, 94.4) = 4.03, p < 0.001) increased the number of infusions earned per session, but females took significantly more infusions than males (effect of sex, F(2, 97.3) = 10.6, p = 0.002). However, males and females did not differ in the rate at which their intake escalated (sex × session interaction, F(29, 128.2) = 1.18, p = 0.265).

Infusions during each minute of drug availability. Each Drug Available (DA) period during IntA lasted for 5 min. Across all 5 min, males and females both escalated cocaine intake as a function of increasing IntA experience, but overall, females consumed more cocaine than males (a). Analysis of each minute of the DA periods revealed that all of the escalation of intake in both sexes occurred during the first minute of drug availability, and there was no sex difference in cocaine consumption during the first minute, or in escalation of intake (b). There was no escalation of intake during minutes 2–5 of the DA periods, but females consumed more cocaine than males in minutes 2–5 (c–f). Each point represents the average of three consecutive IntA sessions, and arrowheads mark when Probe Tests were conducted. Values represent means ± SEMs
In light of the difference in the number of infusions earned, we analyzed the temporal pattern of intake in males and females (Fig. 4b–f). Both males and females took most of their infusions during the first minute of each Drug Available period, and the number of infusions earned in this 1st minute increased as a function of IntA experience (effect of session, F(29, 212.8) = 10.5, p < 0.001). However, there was no sex difference in the number of infusions earned in the 1st minute of the Drug-Available periods (effect of sex, F(1, 17.3) = 1.98, p = 0.18), nor in the rate of escalation of intake (sex × session interaction, F(29, 212.8) = 0.55, p = 0.97). Further, all of the intake escalation that occurred across sessions took place in the first minute, as there was no effect of session on the number of infusions earned in the 2nd, 3rd, 4th, or 5th minute of the Drug Available periods (all p values > 0.05). However, females took more infusions than males in minutes 2 (p < 0.001), 3 (p < 0.001), 4 (p < 0.001), and 5 (p = 0.007), which accounts for their greater total intake evident in Fig. 4a.
Motivation for cocaine increased with IntA experience in both males and females, but females showed a more rapid and greater increase in motivation than males
Rats were tested for their motivation to self-administer cocaine using the within-session threshold procedure prior to IntA experience, after 10 IntA sessions, and then again after 30 IntA sessions. Motivation for cocaine increased as a function of IntA experience in both sexes, indicated by a significant main effect of probe test on α (F(2, 62.8) = 7.01, p = 0.002; Fig. 5a). In addition, females were more motivated than males on these probe tests indicated by a lower α (effect of sex, F(1, 32.2) = 14.4, p = 0.001). When analyzed across all 3 of these probe tests, the motivation of females did not change to a greater extent than that of males (session × sex interaction, F(2, 62.8) = 2.21, p = 0.119). However, to assess how motivation changed with limited IntA experience, we also compared only the baseline probe test and 1st probe test (after 10 IntA sessions), as this was when the greatest change was seen in females. This yielded a significant interaction (session × sex interaction, F(1, 32.6) = 4.16, p = 0.049), suggesting that motivation for cocaine increased more rapidly in females than males.

Cocaine demand in males and females as a function of IntA self-administration experience. Cocaine demand was assessed prior to IntA experience (Baseline, BL), and then after 10 (Probe Test 1) and 30 (Probe Test 2) days of IntA experience, and again after a 14-day abstinence period (Probe Test 3; n = 18 males, 16 females). The elasticity of the demand curves (α) decreased (motivation increased) in both males and females with increasing IntA experience and further decreased following the abstinence period. α values decreased to a greater extent in females than males after only 10 IntA sessions and remained lower in females throughout testing (a). Q0 (preferred cocaine intake when cost is nil) did not change with increasing IntA experience and did not differ between males and females (b). Panels (c) and (d) show representative demand curves for male (c) and female (d) rats derived from Probe Test 3 (P3; after 30 IntA sessions and the abstinence period). In panels (a) and (b), the arrowhead marks a 14-day drug-free abstinence period. Values represent means ± SEMs
Rats were also tested on the within-session threshold procedure after a 14-day abstinence period that followed 30 IntA sessions. A test comparing motivation (α) on the 3rd probe test to the 4th probe test (post-abstinence) indicated a significant increase in motivation following the abstinence period (F(1, 57) = 4.1, p = 0.048) and that females remained more motivated than males (effect of sex, F(1, 57) = 25.8, p < 0.001; test × sex interaction, F(1,57) = 0.825, p = 0.37). We also analyzed Pmax using identical statistical tests to α and for each test the results (significant/non-significant) were consistent (data not shown).
Another measure that can be extracted from the within-session threshold procedure is Q0, a measure of preferred drug consumption when cost is essentially zero (Fig. 5b). There was no significant change in Q0 with increasing IntA experience (F(2, 41.8) = 0.27, p = 0.763), nor was there a difference in Q0 between males and females (F(1, 49) = 0.99, p = 0.325). In addition, there was no change in Q0 and no difference between males and females, following the 14-day abstinence period (all p values > 0.1).
Figure 5c and d shows demand curves for a representative male and representative female from the final probe test (following the 14-day abstinence period).
There was no correlation between total cocaine intake during IntA and α, but there was a correlation between IntA intake and Q 0
One possible interpretation of our results was that the greater motivation (α) that was observed in females was the result of them consuming more cocaine (relative to body weight) than males during IntA. In order to test this, we correlated α values from the 2nd probe test (post-30 IntA sessions) with the average number of infusions self-administered across the last 3 IntA sessions in both males and females (Fig. 6a,b). There was no correlation in males (R2 = 0.06, p = 0.30) or females (R2 = 0.01, p = 0.73). We conducted the same correlation analysis on α values from the 1st probe test (post-10 IntA sessions) with the average number of infusions self-administered across sessions 8–10 of IntA (data not shown). Again, there was no correlation in males (R2 < 0.01, p = 0.92) or females (R2 = 0.1, p = 0.17) suggesting that IntA intake and motivation on the within-session threshold procedure are dissociable.

Correlations between the behavioral economic metrics and cocaine consumption during IntA. Each rats’ α value from Probe Test 2 (after 30 IntA sessions) was correlated with the average number of infusions that rat self-administered over the last 3 IntA sessions in males (a) and females (b). α was not correlated with IntA intake in either males or females. Q0 (preferred cocaine intake when cost is nil) taken from the Baseline Probe Test (BL) was positively correlated with cocaine intake during the 5-min Drug Available (DA) periods of the first 3 IntA sessions (c), and Q0 taken from Probe Test 2 (P2) was similarly correlated with cocaine intake during the DA periods of the last 3 IntA sessions (d). Males and females were combined for panels (c) and (d) as there were no sex differences in Q0
We also asked whether Q0 from the within-session threshold procedure was correlated with IntA intake (Fig. 6c,d). To do this, we took the average intake during a Drug-Available block in both males and females from early-IntA (sessions 1–3) and correlated this with Q0 values from the baseline probe test (prior to IntA). We combined males and females for this analysis because Q0 did not differ between the sexes. Our results showed a significant correlation between preferred cocaine consumption (Q0) and IntA intake (R2 = 0.12, p = 0.03; Fig. 6c). In addition to the correlation, it can be seen that rats “load-up” to similar levels during each Drug-Available block of IntA as they do during the within-session threshold procedure. Further, when Q0 was taken from the 2nd probe test (post-30 IntA sessions) and correlated with intake from the last 3 IntA sessions, these measures were still significantly correlated (R2 = 0.19, p = 0.02; Fig. 6d), perhaps suggesting that these two measures of consumption reflect similar processes before and after escalation of cocaine intake.
There was no sex difference in drug- or cue-induced reinstatement
Following the 3rd and final probe test, the rats underwent a 2-day, drug-induced reinstatement test. The first day consisted of a 90-min extinction period followed by saline infusions, and the second day consisted of a 90-min extinction period followed by a series of cocaine infusions (see the “Materials and methods” section). Males and females did not differ in their extinction responding on the first day (effect of sex, F(2, 27.3) = 3.75, p = 0.063; sex × time interaction, F(17, 55.9) = 1.67, p = 0.08; Fig. 7a) or the extinction portion of the second day (data not shown). The cocaine-priming injections dose-dependently reinstated drug-seeking behavior in both males and females, as measured by responses in the previously active nose port (effect of dose, F(4, 26.4) = 8.34, p < 0.001; Fig. 7b) but not to a different extent in males and females (effect of sex, F(1, 25.0) = 2.89, p = 0.1). Notably, there was no effect of dose on responses at the inactive nose port (effect of dose, p > 0.1).

Cocaine- and cue-induced reinstatement of drug seeking. Males and females did not differ in responding during a within-session extinction procedure (see the “Materials and methods” section) (a) or in the extent to which multiple doses of cocaine (IV) reinstated drug-seeking (b). Males and females also did not differ in the number of responses made or the number of sessions required to reach extinction criteria during a between-session extinction procedure (c). Males and females did not differ in the number of responses made at the previously active port when a nose poke resulted in the presentation of the cue alone that had previously been paired with cocaine (d). Nose pokes at the inactive nose port are represented by the dashed lines. N = 18 males, 16 females. Values represent means ± SEMs
Following the drug-induced reinstatement test, rats underwent additional extinction training for at least 5 days and then a test for cue-induced reinstatement of cocaine-seeking behavior (conditioned reinforcement). Again, males and females did not differ in either the number of responses made during extinction or the number of sessions required to meet a predetermined extinction criteria (all p values > 0.1; Fig. 7c). Both males and females showed robust cue-induced drug-seeking at the previously active nose port, compared to the last day of extinction (effect of session, F(1, 23.0) = 106, p < 0.001; Fig. 7d). In addition, there was no effect of sex on cue-induced reinstatement (effect of sex, F(1, 23.0) = 0.58, p = 0.45; session × sex interaction, (F(1, 23.0) = 0.62, p = 0.44). Notably, there was no effect of session on responses directed toward the inactive nose port (effect of session, p > 0.1).
Discussion
IntA cocaine self-administration experience models intermittent patterns of cocaine use during the transition to addiction in humans (for review, see Allain et al. 2015). A history of IntA experience increases subsequent motivation for cocaine in male rats—that is, it produces incentive-sensitization (Zimmer et al. 2012; Kawa et al. 2016; Allain et al. 2017; Singer et al. 2018), and it does so more effectively than LgA self-administration experience (Zimmer et al. 2012; Allain et al. 2018; Allain and Samaha 2018; James et al. 2018). Given the clinical literature suggesting that women transition to addiction more readily than men (see the “Introduction” section), we asked whether there are sex differences in incentive-sensitization produced by IntA experience in rats.
Motivation for cocaine was assessed before, during, and after IntA experience, using behavioral economic procedures that allowed us to quantify changes in the “elasticity” of cocaine demand curves—that is, the degree to which rats would work to defend their preferred level of cocaine consumption as cost (in effort) progressively increased. The elasticity of a demand curve is quantified by a metric called α (see the “Materials and methods” section), which provides a measure of motivation for a commodity (cocaine in this case). Lower α values reflect less elasticity and, therefore, greater motivation. Relatively limited IntA self-administration experience decreased α more readily in females than males, and α was lower in females than males throughout IntA testing. This is consistent with a study using similar behavioral economic metrics to measure sex differences in the motivation to self-administer methamphetamine after a relatively limited number of 2-h self-administration sessions (Cox et al. 2017). Motivation for cocaine further increased in males and females after 2 weeks of drug abstinence but still remained higher in females. In addition, females made more responses during the No-Drug Available periods (especially toward the end of these periods)—suggesting greater anticipation of the next Drug Available period (see Kawa et al. 2016 discussion). These findings are consistent, therefore, with the hypothesis that females are more susceptible to incentive-sensitization than males.
Females also consumed more cocaine during IntA than males, similar to studies using other self-administration procedures (Lynch and Taylor 2004; Roth and Carroll 2004; Lynch et al. 2005; Smith et al. 2011) and other drugs of abuse (Cicero et al. 2003; Hwa et al. 2011). During IntA, most cocaine consumption occurs within the first minute of each 5 min Drug Available period (Kawa et al. 2016; Allain et al. 2018), and that was also the case here, for both males and females. Interestingly, there were no sex differences in cocaine intake during the first minute of drug availability. Furthermore, the escalation of intake seen with increasing IntA experience was confined to this first minute, as described previously (Allain et al. 2018), and there was no sex difference in the degree of escalation. The sex difference in total drug consumption was entirely due to females taking more cocaine than males in minutes 2–5 of drug availability, when no escalation of intake was observed. Of course, it is possible that the greater incentive-sensitization seen in females was a result of them consuming more cocaine. However, there was no correlation between the total amount of drug intake and motivation for cocaine (α), as has been reported by others for male rats (Allain et al. 2018), suggesting this is not the explanation.
In contrast with sex differences in motivation for cocaine, males and females did not differ in Q0, a measure of the preferred level of consumption when cost is low, nor did Q0 change in either sex with increasing IntA experience or following a 14-day abstinence period, consistent with previous reports using males (Kawa et al. 2016; Singer et al. 2018; James et al. 2018). This suggests that IntA produced robust changes in motivation for cocaine (“wanting”) without altering the desired effects produced by the preferred brain level of cocaine, which could in part reflect the degree of cocaine “liking.” If so, the absence of a sex difference in Q0 may be consistent with reports that in humans the degree of enjoyment derived from cocaine use does not differ between the sexes, despite other differences in addiction vulnerability (e.g., Kennedy et al. 2013). It is also worth noting, that unlike IntA, LgA cocaine self-administration experience produces a persistent increase in Q0 (Oleson and Roberts 2009; Bentzley et al. 2014; James et al. 2018).
Using the within-session threshold procedure to quantify cocaine consumption and motivation
Throughout the study, we relied on behavioral economic analysis of data derived from the within-session threshold procedure. Traditionally, manipulations of “unit price” can be achieved through two routes. One method is to change the number of responses required to obtain a fixed amount of the commodity (often referred to as a progressive-ratio test), and the other (used in this study) is to hold the response requirement the same throughout the session but change the amount (or dose) of commodity delivered per response. Both manipulations equivalently affect the “unit price” of a commodity, and generally, individuals will titrate their responding similarly in both cases (Collier et al. 1986; Hursh et al. 1988; Bickel et al. 1990; but see Gan et al. 2010). One advantage of using threshold procedures over PR procedures is that they differentiate between the preferred level of consumption and motivation to work for the commodity (Oleson and Roberts 2009; Oleson et al. 2011). That is, threshold procedures yield metrics for the preferred level of drug consumption when little, or no effort is required, and separately, the amount of effort a subject is willing to expend for drug when the amount of effort required is increased. Distinct psychological and neurobiological systems have been implicated in the preferred level of consumption and motivation (Brebner et al. 2000; Nicola and Deadwyler 2000; Sharpe and Samson 2001; España et al. 2010; Gan et al. 2010; Oleson et al. 2011; Guillem et al. 2014). For example, α and Q0 are dissociable metrics and have been shown to measure different processes (Bentzley et al. 2013, 2014; Kawa et al. 2016; James et al. 2018; Fig. 6). In addition, a number of studies have shown that the within-session threshold procedure is a valid technique for manipulating “unit price” including the following: studies that show manipulations similarly and predictably affect the “motivation” measures on the within-session threshold procedure and “breakpoint” from a PR test (Oleson et al. 2011); economic demand (but not preferred consumption) derived from the within-session threshold procedure predicts other addiction-like behaviors (Bentzley et al. 2014); and finally decreased responding during the within-session threshold procedure is the result of increased “unit price” and not session length or satiation—demonstrated by presenting doses in an ascending order or with a 20-min time-out (intermittent threshold procedure) between each unit price (Porter-Stransky et al. 2017).
Are females especially susceptible to the effects of intermittent drug experience?
The “telescoping effect” describes the observation that women transition from first drug use to addiction more rapidly and to a more severe extent than men (Anglin et al. 1987a, b; Griffin et al. 1989; Kosten et al. 1993; Brady and Randall 1999). Our present findings show that after only 10 days of IntA experience, motivation for cocaine (α) increased more in female rats than in male rats, and starting from the first IntA session, females consumed more cocaine. These findings are both consistent with the telescoping effect and with several other pre-clinical studies (Lynch and Carroll 1999; Hu et al. 2004). It should be noted that females’ motivation for cocaine may also increase following other, less intermittent, self-administration procedures (Lynch and Taylor 2004; but also see Ramôa et al. 2013, 2014); however, it appears that IntA was especially effective in increasing motivation for cocaine in females, relative to males. Earlier studies involving experimenter-administered drug injections are also consistent with the idea that females are particularly susceptible to intermittent drug exposure. Females show greater psychomotor sensitization (Robinson 1984; Robinson and Becker 1986; van Haaren and Meyer 1991) and more rapidly induced conditioned place preference (Russo et al. 2003) following repeated, experimenter-administered psychomotor-stimulant drug injections.
A number of factors may contribute to the telescoping effect reported in women. As suggested previously, intermittent exposure to drugs may more readily produce sensitization-related changes in brain systems that mediate the psychomotor activating and incentive motivational effects of drugs in females, thus promoting a more rapid and pronounced transition from casual drug use to addiction. Another factor that might contribute to the telescoping effect is that it has been reliably shown that the reinforcing effects of cocaine vary across the estrous or menstrual cycle (for reviews, see Lynch 2018; Yoest et al. 2018). Thus, when females self-administer cocaine across multiple days of their cycle, the drug experience may be more variable than in males, and thus by definition, more intermittent. Given that intermittency promotes incentive-sensitization, this could contribute to the sex differences described here, and to the telescoping effect in humans.
Female behavior was not more variable than male behavior despite not measuring estrous phase
As noted in the “Materials and methods” section, we did not monitor the estrous cycle. A great deal of work has been done to elucidate how circulating hormones interact with the brain and the role this plays in addiction (for reviews, see Becker et al. 2012; Lynch 2018; Yoest et al. 2018). Based on this large body of literature, it is likely that our behavioral measures would have been affected by phase of the cycle. For example, it is possible that on the baseline probe test, females tested during estrus would have been more motivated than non-estrous females and males. However, in general, the magnitude of the effects of estrous cycle in females is small relative to the effects of sex and IntA training on motivation that we observed here. It is also important to note that at no point in our results was the variability of female behavior larger than the variability of male behavior, consistent with a recent meta-analysis of data collected from rats that concluded that female behavior was not more variable than male behavior, even when females are tested without regard for estrous phase (Becker et al. 2016).
Males and females did not differ in rate of acquisition or reinstatement
In this study, males and females did not differ in the rate that they acquired self-administration behavior nor in the number of responses made during acquisition (Fig. 2a,b). This is in contrast to several previous reports that females acquire self-administration faster than males at low doses of cocaine (Lynch and Carroll 1999) and when ovariectomized with or without estradiol treatment (Hu et al. 2004). However, this discrepancy is probably related to procedural differences in how the rats were trained to self-administer cocaine. The present study utilized an “infusion criteria” procedure that was developed to explicitly minimize individual differences (Saunders and Robinson 2010), by ensuring that all rats experience the same amount of drug and cue exposure prior to any subsequent testing.
Another interesting finding is that males and females did not differ during extinction or on the drug- or cue-induced reinstatement tests (Fig. 7). The majority of extinction-reinstatement studies find that females respond more during the early stages of extinction but do not differ from males in the number of extinction sessions required to meet a predetermined criteria (Lynch et al. 2005; Fuchs et al. 2005; Kippin et al. 2005; Feltenstein et al. 2011). Similar to these studies, we did not see any sex difference in the number of sessions required to meet an extinction criteria. However, we did not see increased responding in females during early extinction. We believe that this is most likely due to different self-administration procedures used in these different studies. Here, we used the IntA procedure, and rats were well-accustomed to periods of No-Drug availability (essentially extinction conditions), and the finding that females responded more during the No-Drug periods of IntA is consistent with this interpretation (Fig. 3d). It is worth noting that males and females did not differ in learning to discriminate between the Drug Available periods and No-Drug periods of IntA, and by session 5 of IntA, both males and females could clearly discriminate between the two periods (Fig. 3a,b). Due to their experience with the No-Drug periods of IntA, both males and females stopped responding relatively quickly during the extinction training that preceded the reinstatement tests (i.e., Fig. 7a).
There is a lack of a consensus on whether females are more motivated by cocaine-paired cues following cocaine self-administration. There are several reports in humans that cocaine-associated cues induce stronger craving in females (Robbins et al. 1999; Elman et al. 2001; Kennedy et al. 2013), although this is not always the case (Negrete and Emil 1992; Avants et al. 1995). However, most preclinical studies report either no sex difference in cue-induced reinstatement (Lynch et al. 2005; Fuchs et al. 2005; Kerstetter et al. 2008; Feltenstein et al. 2011) or greater reinstatement in males dependent on the dose of cocaine used during training (Fuchs et al. 2005). Thus, the present results (Fig. 7d) add to the literature suggesting that female rats are not more susceptible to cue-induced reinstatement than male rats when testing occurs without respect to the estrous cycle.
Studies on cocaine-induced reinstatement more consistently demonstrate increased drug-seeking in females following a cocaine priming injection, particularly during estrus (Lynch and Carroll 2000; Kippin et al. 2005; Kerstetter et al. 2008). Given these results, we were surprised that there was no sex difference in cocaine-induced reinstatement (Fig. 7b). Further work is required to determine how different self-administration procedures affect the conditioned motivating effects of cues and non-contingent cocaine infusions. It is clear however that IntA experience results in very robust drug- and cue-induced reinstatement in both male and female rats, relative to what is typically reported following self-administration experience with other self-administration procedures (also see Kawa et al. 2016).
Conclusion
In summary, the present results add to a growing literature showing that IntA cocaine self-administration experience is especially effective in producing addiction-like behavior, despite resulting in much less total drug consumption than more widely used LgA procedures (Zimmer et al. 2012; Kawa et al. 2016; Allain et al. 2018; Allain and Samaha 2018; James et al. 2018). Furthermore, we not only extend this observation to female rats, but find that females are more susceptible to incentive-sensitization produced by intermittent exposure to cocaine. This sex difference may be related to the “telescoping effect” reported in women during the transition to addiction and may therefore provide a useful approach for studying the neural basis of this phenomenon.
References
Ahmed SH (2012) The science of making drug-addicted animals. Neuroscience 211:107–125. https://doi.org/10.1016/j.neuroscience.2011.08.014
Ahmed SH, Koob GF (1998) Transition from moderate to excessive drug intake: change in hedonic set point. Science 282:298–300. https://doi.org/10.1126/science.282.5387.298
Allain F, Samaha A-N (2018) Revisiting long-access versus short-access cocaine self-administration in rats: intermittent intake promotes addiction symptoms independent of session length. Addict Biol in press. https://doi.org/10.1111/adb.12629
Allain F, Minogianis E-A, Roberts DCS, Samaha A-N (2015) How fast and how often: the pharmacokinetics of drug use are decisive in addiction. Neurosci Biobehav Rev 56:166–179. https://doi.org/10.1016/j.neubiorev.2015.06.012
Allain F, Roberts DCS, Lévesque D, Samaha A-N (2017) Intermittent intake of rapid cocaine injections promotes robust psychomotor sensitization, increased incentive motivation for the drug and mGlu2/3 receptor dysregulation. Neuropharmacology 117:227–237. https://doi.org/10.1016/j.neuropharm.2017.01.026
Allain F, Bouayad-Gervais K, Samaha A-N (2018) High and escalating levels of cocaine intake are dissociable from subsequent incentive motivation for the drug in rats. Psychopharmacology 235:317–328. https://doi.org/10.1007/s00213-017-4773-8
Anglin MD, Hser YI, Booth MW (1987a) Sex differences in addict careers. 4. Treatment. Am J Drug Alcohol Abuse 13:253–280. https://doi.org/10.3109/00952998709001512
Anglin MD, Hser YIL, Mcglothlin WH (1987b) Sex differences in addict careers. 2. Becoming addicted. Am J Drug Alcohol Abuse 13:59–71. https://doi.org/10.3109/00952998709001500
Avants SK, Margolin A, Kosten TR, Cooney NL (1995) Differences between responders and nonresponders to cocaine cues in the laboratory. Addict Behav 20:215–224. https://doi.org/10.1016/0306-4603(94)00066-2
Becker JB (2016) Sex differences in addiction. Dialogues Clin Neurosci 18:395–402. https://doi.org/10.1016/B978-0-12-802114-9/00006-8
Becker JB, Koob GF (2016) Sex differences in animal models: focus on addiction. Pharmacol Rev 68:242–263. https://doi.org/10.1124/pr.115.011163
Becker JB, Perry AN, Westenbroek C (2012) Sex differences in the neural mechanisms mediating addiction: a new synthesis and hypothesis. Biol Sex Differ 3:14. https://doi.org/10.1186/2042-6410-3-14
Becker JB, Prendergast BJ, Liang JW (2016) Female rats are not more variable than male rats: a meta-analysis of neuroscience studies. Biol Sex Differ 7:34. https://doi.org/10.1186/s13293-016-0087-5
Bentzley BS, Fender KM, Aston-Jones G (2013) The behavioral economics of drug self-administration: a review and new analytical approach for within-session procedures. Psychopharmacology 226:113–125. https://doi.org/10.1007/s00213-012-2899-2
Bentzley BS, Jhou TC, Aston-Jones G (2014) Economic demand predicts addiction-like behavior and therapeutic efficacy of oxytocin in the rat. Proc Natl Acad Sci 111:11822–11827. https://doi.org/10.1073/pnas.1406324111
Berridge KC, Robinson TE (2016) Liking, wanting, and the incentive-sensitization theory of addiction. Am Psychol 71:670–679. https://doi.org/10.1037/amp0000059
Bickel WK, DeGrandpre RJ, Higgins ST, Hughes JR (1990) Behavioral economics of drug self-administration. I. Functional equivalence of response requirement and drug dose. Life Sci 47:1501–1510. https://doi.org/10.1016/0024-3205(90)90178-T
Bowman BP, Vaughan SR, Walker QD, Davis SL, Little PJ, Scheffler NM, Thomas BF, Kuhn CM (1999) Effects of sex and gonadectomy on cocaine metabolism in the rat. J Pharmacol Exp Ther 290:1316–1323
Brady KT, Randall CL (1999) Gender differences in substance use disorders. Psychiatr Clin North Am 22:241–252. https://doi.org/10.1016/S0193-953X(05)70074-5
Brebner K, Phelan R, Roberts DCS (2000) Effect of baclofen on cocaine self-administration in rats reinforced under fixed-ratio 1 and progressive-ratio schedules. Psychopharmacology 148:314–321. https://doi.org/10.1007/s002130050056
Cicero TJ, Aylward SC, Meyer ER (2003) Gender differences in the intravenous self-administration of mu opiate agonists. Pharmacol Biochem Behav 74:541–549. https://doi.org/10.1016/S0091-3057(02)01039-0
Collier GH, Johnson DF, Hill WL, Kaufman LW (1986) The economics of the law of effect. J Exp Anal Behav 46:113–136. https://doi.org/10.1901/jeab.1986.46-113
Cox BM, Bentzley BS, Regen-Tuero H, See RE, Reichel CM, Aston-Jones G (2017) Oxytocin acts in nucleus accumbens to attenuate methamphetamine seeking and demand. Biol Psychiatry 81:949–958. https://doi.org/10.1016/j.biopsych.2016.11.011
Crombag HS, Badiani A, Maren S, Robinson TE (2000) The role of contextual versus discrete drug-associated cues in promoting the induction of psychomotor sensitization to intravenous amphetamine. Behav Brain Res 116:1–22. https://doi.org/10.1016/S0166-4328(00)00243-6
Cummings JA, Gowl BA, Westenbroek C, Clinton SM, Akil H, Becker JB (2011) Effects of a selectively bred novelty-seeking phenotype on the motivation to take cocaine in male and female rats. Biol Sex Differ 2:3. https://doi.org/10.1186/2042-6410-2-3
Deroche V, Le Moal M, Piazza PV (1999) Cocaine self-administration increases the incentive motivational properties of the drug in rats. Eur J Neurosci 11:2731–2736. https://doi.org/10.1046/j.1460-9568.1999.00696.x
Elman I, Karlsgodt KH, Gastfriend DR (2001) Gender differences in cocaine craving among non-treatment-seeking individuals with cocaine dependence. Am J Drug Alcohol Abuse 27:193–202. https://doi.org/10.1081/ADA-100103705
España RA, Oleson EB, Locke JL, Brookshire BR, Roberts DCS, Jones SR (2010) The hypocretin-orexin system regulates cocaine self-administration via actions on the mesolimbic dopamine system. Eur J Neurosci 31:336–348. https://doi.org/10.1111/j.1460-9568.2009.07065.x
Fattore L, Altea S, Fratta W (2008) Sex differences in drug addiction: a review of animal and human studies. Women's Health (Lond Engl) 4:51–65. https://doi.org/10.2217/17455057.4.1.51
Feltenstein MW, Henderson AR, See RE (2011) Enhancement of cue-induced reinstatement of cocaine-seeking in rats by yohimbine: sex differences and the role of the estrous cycle. Psychopharmacology 216:53–62. https://doi.org/10.1007/s00213-011-2187-6
Fuchs RA, Evans KA, Mehta RH, Case JM, See RE (2005) Influence of sex and estrous cyclicity on conditioned cue-induced reinstatement of cocaine-seeking behavior in rats. Psychopharmacology 179:662–672. https://doi.org/10.1007/s00213-004-2080-7
Gan JO, Walton ME, Phillips PEM (2010) Dissociable cost and benefit encoding of future rewards by mesolimbic dopamine. Nat Neurosci 13:25–27. https://doi.org/10.1038/nn.2460
Griffin ML, Weiss RD, Mirin SM, Lange U (1989) A comparison of male and female cocaine abusers. Arch Gen Psychiatry 46:122–126. https://doi.org/10.1001/archpsyc.1989.01810020024005
Guillem K, Ahmed SH, Peoples LL (2014) Escalation of cocaine intake and incubation of cocaine seeking are correlated with dissociable neuronal processes in different accumbens subregions. Biol Psychiatry 76:31–39. https://doi.org/10.1016/j.biopsych.2013.08.032
van Haaren F, Meyer ME (1991) Sex differences in locomotor activity after acute and chronic cocaine administration. Pharmacol Biochem Behav 39:923–927. https://doi.org/10.1016/0091-3057(91)90054-6
Hu M, Crombag HS, Robinson TE, Becker JB (2004) Biological basis of sex differences in the propensity to self-administer cocaine. Neuropsychopharmacology 29:81–85. https://doi.org/10.1038/sj.npp.1300301
Hursh SR, Silberberg A (2008) Economic demand and essential value. Psychol Rev 115:186–198. https://doi.org/10.1037/0033-295X.115.1.186
Hursh SR, Raslear TG, Shurtleff D, Bauman R, Simmons L (1988) A cost-benefit analysis of demand for food. J Exp Anal Behav 50:419–440. https://doi.org/10.1901/jeab.1988.50-419
Hwa LS, Chu A, Levinson SA, Kayyali TM, DeBold JF, Miczek KA (2011) Persistent escalation of alcohol drinking in C57BL/6J mice with intermittent access to 20% ethanol. Alcohol Clin Exp Res 35:1938–1947. https://doi.org/10.1111/j.1530-0277.2011.01545.x
James MH, Stopper CM, Zimmer BA, Koll NE, Bowrey HE, Aston-Jones G (2018) Increased number and activity of a lateral subpopulation of hypothalamic orexin/hypocretin neurons underlies the expression of an addicted state in rats. Biol Psychiatry. https://doi.org/10.1016/j.biopsych.2018.07.022
Kawa AB, Bentzley BS, Robinson TE (2016) Less is more: prolonged intermittent access cocaine self-administration produces incentive-sensitization and addiction-like behavior. Psychopharmacology 233:3587–3602. https://doi.org/10.1007/s00213-016-4393-8
Kennedy AP, Epstein DH, Phillips KA, Preston KL (2013) Sex differences in cocaine/heroin users: drug-use triggers and craving in daily life. Drug Alcohol Depend 132:29–37. https://doi.org/10.1016/j.drugalcdep.2012.12.025
Kerstetter KA, Aguilar VR, Parrish AB, Kippin TE (2008) Protracted time-dependent increases in cocaine-seeking behavior during cocaine withdrawal in female relative to male rats. Psychopharmacology 198:63–75. https://doi.org/10.1007/s00213-008-1089-8
Kippin TE, Fuchs RA, Mehta RH, Case JM, Parker MP, Bimonte-Nelson HA, See RE (2005) Potentiation of cocaine-primed reinstatement of drug seeking in female rats during estrus. Psychopharmacology 182:245–252. https://doi.org/10.1007/s00213-005-0071-y
Kosten TA, Gawin FH, Kosten TR, Rounsaville BJ (1993) Gender differences in cocaine use and treatment response. J Subst Abus Treat 10:63–66. https://doi.org/10.1016/0740-5472(93)90100-G
Lynch WJ (2018) Modeling the development of drug addiction in male and female animals. Pharmacol Biochem Behav 164:50–61. https://doi.org/10.1016/j.pbb.2017.06.006
Lynch WJ, Carroll ME (1999) Sex differences in the acquisition of intravenously self-administered cocaine and heroin in rats. Psychopharmacology 144:77–82. https://doi.org/10.1007/s002130050979
Lynch WJ, Carroll ME (2000) Reinstatement of cocaine self-administration in rats: sex differences. Psychopharmacology 148:196–200. https://doi.org/10.1007/s002130050042
Lynch WJ, Taylor JR (2004) Sex differences in the behavioral effects of 24-h/day access to cocaine under a discrete trial procedure. Neuropsychopharmacology 29:943–951. https://doi.org/10.1038/sj.npp.1300389
Lynch WJ, Mangini LD, Taylor JR (2005) Neonatal isolation stress potentiates cocaine seeking behavior in adult male and female rats. Neuropsychopharmacology 30:322–329. https://doi.org/10.1038/sj.npp.1300594
Negrete JC, Emil S (1992) Cue-evoked arousal in cocaine users: a study of variance and predictive value. Drug Alcohol Depend 30:187–192. https://doi.org/10.1016/0376-8716(92)90025-8
Nicola SM, Deadwyler SA (2000) Firing rate of nucleus accumbens neurons is dopamine-dependent and reflects the timing of cocaine-seeking behavior in rats on a progressive ratio schedule of reinforcement. J Neurosci 20:5526–5537 doi: 20/14/5526
Ojeda SR, Andrews WW, Advis JP, White SS (1980) Recent advances in the endocrinology of puberty. Endocr Rev 1:228–257. https://doi.org/10.1210/edrv-1-3-228
Oleson EB, Roberts DCS (2009) Behavioral economic assessment of price and cocaine consumption following self-administration histories that produce escalation of either final ratios or intake. Neuropsychopharmacology 34:796–804. https://doi.org/10.1038/npp.2008.195
Oleson EB, Richardson JM, Roberts DCS (2011) A novel IV cocaine self-administration procedure in rats: differential effects of dopamine, serotonin, and GABA drug pre-treatments on cocaine consumption and maximal price paid. Psychopharmacology 214:567–577. https://doi.org/10.1007/s00213-010-2058-6
Piazza NJ, Vrbka JL, Yeager RD (1989) Telescoping of alcoholism in women alcoholics. Subst Use Misuse 24:19–28. https://doi.org/10.3109/10826088909047272
Porter-Stransky KA, Bentzley BS, Aston-Jones G (2017) Individual differences in orexin-I receptor modulation of motivation for the opioid remifentanil. Addict Biol 22:303–317. https://doi.org/10.1111/adb.12323
Ramôa CP, Doyle SE, Naim DW, Lynch WJ (2013) Estradiol as a mechanism for sex differences in the development of an addicted phenotype following extended access cocaine self-administration. Neuropsychopharmacology 38:1698–1705. https://doi.org/10.1038/npp.2013.68
Ramôa CP, Doyle SE, Lycas MD, Chernau AK, Lynch WJ (2014) Diminished role of dopamine D1-receptor signaling with the development of an addicted phenotype in rats. Biol Psychiatry 76:8–14. https://doi.org/10.1016/j.biopsych.2013.09.028
Robbins SJ, Ehrman RN, Childress AR, O’Brien CP (1999) Comparing levels of cocaine cue reactivity in male and female outpatients. Drug Alcohol Depend 53:223–230. https://doi.org/10.1016/S0376-8716(98)00135-5
Roberts DCS, Bennett SAL, Vickers GJ (1989) The estrous cycle affects cocaine self-administration on a progressive ratio schedule in rats. Psychopharmacology 98:408–411. https://doi.org/10.1007/BF00451696
Robinson TE (1984) Behavioral sensitization: characterization of enduring changes in rotational behavior produced by intermittent injections of amphetamine in male and female rats. Psychopharmacology 84:466–475. https://doi.org/10.1007/BF00431451
Robinson TE, Becker JB (1986) Enduring changes in brain and behavior produced by chronic amphetamine administration: a review and evaluation of animal models of amphetamine psychosis. Brain Res Rev 11:157–198
Robinson TE, Berridge KC (1993) The neural basis of drug craving: an incentive-sensitization theory of addiction. Brain Res Rev 18:247–291. https://doi.org/10.1016/0165-0173(93)90013-P
Roth ME, Carroll ME (2004) Sex differences in the escalation of intravenous cocaine intake following long- or short-access to cocaine self-administration. Pharmacol Biochem Behav 78:199–207. https://doi.org/10.1016/j.pbb.2004.03.018
Russo SJ, Jenab S, Fabian SJ, Festa ED, Kemen LM, Quinones-Jenab V (2003) Sex differences in the conditioned rewarding effects of cocaine. Brain Res 970:214–220. https://doi.org/10.1016/S0006-8993(03)02346-1
Saunders BT, Robinson TE (2010) A cocaine cue acts as an incentive stimulus in some but not others: implications for addiction. Biol Psychiatry 67:730–736. https://doi.org/10.1016/j.biopsych.2009.11.015
Sharpe AL, Samson HH (2001) Effect of naloxone on appetitive and consummatory phases of ethanol self-administration. Alcohol Clin Exp Res 25:1006–1011. https://doi.org/10.1111/j.1530-0277.2001.tb02309.x
Singer BF, Fadanelli M, Kawa AB, Robinson TE (2018) Are cocaine-seeking “habits” necessary for the development of addiction-like behavior in rats? J Neurosci 38:60–73. https://doi.org/10.1523/JNEUROSCI.2458-17.2017
Sisk CL, Zehr JL (2005) Pubertal hormones organize the adolescent brain and behavior. Front Neuroendocrinol 26:163–174. https://doi.org/10.1016/j.yfrne.2005.10.003
Smith MA, Walker KL, Cole KT, Lang KC (2011) The effects of aerobic exercise on cocaine self-administration in male and female rats. Psychopharmacology 218:357–369. https://doi.org/10.1007/s00213-011-2321-5
Vezina P (2004) Sensitization of midbrain dopamine neuron reactivity and the self-administration of psychomotor stimulant drugs. Neurosci Biobehav Rev 27:827–839. https://doi.org/10.1016/j.neubiorev.2003.11.001
West BT, Welch KB, Ga AT, Crc H (2007) Linear mixed models: a practical guide using statistical software
Yoest KE, Quigley JA, Becker JB (2018) Rapid effects of ovarian hormones in dorsal striatum and nucleus accumbens. Horm Behav. https://doi.org/10.1016/j.yhbeh.2018.04.002
Zimmer BA, Oleson EB, Roberts DC (2012) The motivation to self-administer is increased after a history of spiking brain levels of cocaine. Neuropsychopharmacology 37:1901–1910. https://doi.org/10.1038/npp.2012.37
Funding
This research was supported by grants PO1 DA031656 and T32 DA007281 from NIDA to TER.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Rights and permissions
About this article

Cite this article
Kawa, A.B., Robinson, T.E. Sex differences in incentive-sensitization produced by intermittent access cocaine self-administration. Psychopharmacology 236, 625–639 (2019). https://doi.org/10.1007/s00213-018-5091-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00213-018-5091-5