
Abstract
Rationale
Discrete cues, such as drug-associated paraphernalia, play an important role in tobacco smoking and relapse, an effect that can be modelled in the nicotine-seeking behaviour of laboratory animals. However, the role of contextual stimuli (i.e. the drug taking environment) within nicotine dependence is less clear. The present study investigated the effects of contextual stimuli on nicotine detoxification and relapse.
Materials and methods
Male hooded Lister rats were trained to self-administer nicotine (0.03 mg/kg/infusion) in one of two distinct environmental contexts: transparent walls and rod floor or checkerboard walls and grid floor. Extinction of drug-seeking behaviour, either in the acquisition context or alternate context, was achieved by removing both nicotine infusions and response-contingent cues. The two contexts were then presented with or without nicotine priming and response-contingent cue presentation.
Results
The initial rate of extinction was quicker in a novel environment compared to in the same context as training, although similar low levels of responding were eventually reached. Nicotine priming and re-presentation of cues resulted in significant reinstatement of nicotine-seeking behaviour, but there was a trend towards a reduction in this effect when conducted in a novel environment. In addition, re-presentation of the acquisition context after extinction in the alternate context produced a significant reinstatement of nicotine-seeking behaviour without the need for nicotine priming and re-presentation of cues.
Conclusions
Contextual stimuli are capable of modulating the extinction and reinstatement of nicotine-seeking behaviour, and exposure to environments previously associated with smoking may lead to an increased risk of relapse. Context is an additional factor that could be targeted when developing smoking cessation strategies. For example, the long-term success of cue exposure might be improved by conducting treatment in multiple settings.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Tobacco smoking is the one of the most prevalent and persistent forms of drug addiction (Anthony et al. 1994). However, nicotine’s euphoric and reinforcing properties are weaker than those of other drugs (Goldberg et al. 1981; Rose and Corrigall 1997; Rose and Levin 1991), and it has been suggested that other factors may be involved in the maintenance and relapse of smoking behaviour. For example, non-pharmacological stimuli, in the form of discrete cues, appear to be involved in both animal and human nicotine self-administration (Caggiula et al. 2001, 2002a; Rose and Corrigall 1997).
Classical conditioning paradigms can be used to create a conditioned stimulus (CS) by pairing a neutral stimulus with the availability of a primary reinforcer (Pavlov 1927), and operant procedures can reveal the conditioned reinforcing properties of the CS. Studies with a variety of drugs of abuse have highlighted the importance of CS in animal self-administration (Arroyo et al. 1998; Goldberg and Gardner 1981; Markou et al. 1993; Schenk and Partridge 2001; See et al. 1999; Spealman et al. 1999) and drug dependence in humans (O’Brien et al. 1998).
Smoking, compared to other routes of nicotine administration, may be a particularly effective way of establishing CS. Smoking cues (e.g. a lit cigarette in an ashtray) increase craving in abstinent and non-abstinent smokers (Perkins et al. 1994), and neutral stimuli paired with availability of smoking can increase smoking urges (Lazev et al. 1999) and time spent smoking (Mucha et al. 1998). Conversely, blockade of smoking cues reduces subjective liking and satisfaction (Baldinger et al. 1995; Perkins et al. 2001; Rose et al. 1985). De-nicotinised cigarettes (i.e. cues) and nicotine cigarettes result in equal smoke intake and reductions in subjective craving and withdrawal (Butschky et al. 1995; Gross et al. 1997), and de-nicotinised cigarettes cause a greater increase in ratings of subjective ‘liking’ and ‘satisfaction’ than intravenous nicotine, although the greatest effects were observed when both cues and nicotine were combined (Rose et al. 2000). Smokers whose ‘liking’ and ‘satisfaction’ of de-nicotinised was similar to nicotine cigarettes tended to be more heavily dependent (Brauer et al. 2001); therefore, cues may be more important in maintaining, rather than establishing, smoking behaviour.
The two most widely used procedures to assess the reinforcing properties of drugs in animals, intravenous self-administration and conditioned place-preference, have been used to demonstrate the influence of environmental stimuli on nicotine reinforcement in the laboratory (Caggiula et al. 2001; Le Foll and Goldberg 2005). This effect was first reported with nicotine self-administration in squirrel monkeys, where omitting a brief light stimulus that had been associated with the drug reduced responding (Goldberg et al. 1981). The Caggiula laboratory has since conducted a series of rodent intravenous self-administration experiments demonstrating the interaction of nicotine’s primary reinforcing effects with environmental cues. Acquisition of nicotine self-administration behaviour is significantly enhanced by presentation of nicotine-related cues (Caggiula et al. 2002b), which also help maintain nicotine-seeking behaviour (Donny et al. 1999). Response-contingent, nicotine-associated visual cues retard extinction and are capable of maintaining behaviour at a similar level to nicotine alone although at lower levels than when cues and nicotine are combined (Caggiula et al. 2001). Following extinction by removal of drug cues, re-presentation of the cues can reinstate lever pressing behaviour, and this is even more effective when the cues are presented in combination with nicotine priming (Caggiula et al. 2001).
Given the important role of discrete cues in smoking behaviour, it is possible that contextual cues may also contribute to nicotine’s addictive properties. It can be difficult to make a clear distinction between contexts and cues, as a context may also act as a CS. However, in the present paper, context will refer to the environment in which drug use takes place and proximal cues are paired with drug taking. It has previously been demonstrated that contextual stimuli are capable of modulating the behavioural and physiological effects of drugs of abuse in the laboratory (Crombag et al. 2000; Robinson et al. 1998) and environments previously associated with opiate intake can provoke drug relapse during abstinence in humans (O’Brien et al. 1992).
In studies using non-drug reinforcers, conditioning generalises readily, whereas the extinction and resumption of conditioned–unconditioned stimulus relationships are believed to be highly context dependent. This is highlighted by the renewal, reinstatement, spontaneous recovery and reacquisition phenomena (Bouton 2004). Renewal refers to a robust return of conditioned responding due to a change in context after extinction. In “ABA renewal” conditioning is conducted in one context (A), extinction in a second (B) and when the CS is returned to original conditioning context (A) responding to the CS returns (Bouton and Bolles 1979). Adapting this procedure for drug self-administration studies (Crombag and Shaham 2002) has demonstrated the reinstatement of heroin (Bossert et al. 2004), cocaine (Crombag et al. 2002; Fuchs et al. 2005) and alcohol seeking (Burattini et al. 2006) by the drug-associated context after extinction in an alternative context.
In regard to nicotine, human studies have documented strong subjective responding from abstinent smokers exposed to distal cues functioning as CS, such as the drug-taking environment, without the presence of proximal smoking cues (Conklin 2006). However, pre-clinically, there is limited information regarding nicotine’s interaction with contextual stimuli, although spontaneous recovery of nicotine-seeking behaviour was observed when rats were re-exposed to the drug-taking environment after a period of enforced abstinence (Shaham et al. 1997).
These findings suggest that contextual stimuli previously associated with nicotine delivery may contribute to smoking behaviour and warrants further investigation. The intravenous self-administration reinstatement procedure will be employed due to its face validity as a laboratory model of drug relapse (Shaham et al. 2003) and its possible predictive validity (Epstein et al. 2006). Altering the context during extinction and reinstatement of nicotine-seeking behaviour could elucidate the role of contextual stimuli in these processes.
Methods
Animals
Male hooded Lister rats (Harlan, UK), initially weighing 280–300 g, were pair housed with food and water available ad libitum, in a room maintained at 20–22°C, humidity controlled, and on a regular 12-h light/dark cycle (light from 7 am to 7 pm). All experiments complied with the local and national ethical requirements and were carried out according to the Animals (Experimental procedures) Act, 1986, under license from the UK Home Office.
Surgery
Rats were implanted with a chronic micro-renathane catheter into the external jugular vein, under surgical anaesthesia (a mixture of medetomidine 0.3 mg/kg and ketamine 70 mg/kg, IP), as previously described (Shoaib et al. 1997; Shoaib et al. 2003). The catheter was secured to the vein using a prolene suture and passed subcutaneously to the top of the skull. It was connected to an L-shaped connector (made in house) and mounted with dental acrylic and screws imbedded in the skull. Patency of the intravenous catheter was maintained by daily flushing with heparinised saline containing Baytril (enthroflaxin) to prevent infection. The self-administration sessions began once animals had returned to their pre-operative weights.
Apparatus
Standard operant chambers (MED-Associates) consisted of a Plexiglas enclosure with a stimulus light, two levers, one tether and fluid swivel connected to an infusion pump (MED-Associates). At the beginning of the study, levers were randomly assigned as active, with pressing resulting in fluid infusions and the termination of the stimulus light that signalled the start of a 20-s time-out period where nicotine is unavailable or as inactive where lever pressing had no programmed consequence. The operant chambers were surrounded by sound-attenuating ventilated cubicles (made in-house) and controlled by a computer using the MED-Associates MED-PC software package.
Drugs
Nicotine hydrogen tartrate salt (Sigma, Dorset) was dissolved in 0.9% physiological saline and the pH adjusted to 7.2 ± 2 with diluted NaOH. A concentration of approximately 0.03 mg kg−1 infusion−1 (expressed as free base weight) was used; this dose has been found to be at or near the peak of the dose–response curve for nicotine self-administration on a limited access fixed-ratio (FR) schedule (Cohen et al. 2002; Corrigall and Coen 1989; Corrigall et al. 1992; Donny et al. 1995, 2000).
Procedure
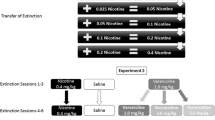
Figure 1 gives an overview of the experimental design. A between-session procedure was implemented, in which training for drug self-administration, extinction of the drug-reinforced behavior and reinstatement tests were conducted during sequential daily sessions.

The experimental design demonstrating the three phases of the protocol: acquisition, extinction (between-group) and reinstatement (within-group)
Two distinct contexts were utilised, a standard Plexiglas enclosure (transparent walls and barred floor) and an adapted Plexiglas enclosure (checkerboard walls and grid floor). To eliminate the factor of bias towards one particular context, the context used in the acquisition phase was counterbalanced across two groups of animals. During extinction, animals were further divided in two with extinction in a context that was the same or different from that during acquisition. If there was no significant effect of the particular context used as the acquisition environment, it was possible to pool the data, giving two groups, one with extinction taking place in the acquisition context (A) and one in the alternative context (B).
Acquisition phase
Rats were given the opportunity to lever press for intravenous infusions of nicotine solution, under an FR schedule, during 1-h daily limited access sessions. Response-contingent cues were presented in the form of termination of the stimulus light (which signals nicotine availability) and the sound of the infusion pump. Once rats showed a response accuracy of at least 80% of the responses being on the active lever and a stable level of nicotine infusions over 2 days, the number of responses required to produce an infusion was increased progressively up to FR-3. Animals entered the extinction phase when they exhibited stable levels of nicotine self-administration on a FR-3 schedule for 3 days (≤ ±20% variability from the mean number of infusions over three sessions).
Extinction phase
Extinction was conducted in the acquisition context (A) or an alternative context (B). During this phase, active lever pressing resulted in no programmed consequence, i.e. no infusion of nicotine and no cue presentation. Reinstatement tests commenced once animals’ responding had met the extinction criterion (≤20% of the average infusions in the three sessions before extinction) for three consecutive days.
Reinstatement phase
Four within-subject reinstatement tests were conducted in a randomised order. The reinstatement tests involved a priming injection of nicotine (0.2 mg/kg, SC) immediately before the self-administration session and presentation of response-contingent cues (stimulus light off) during the session or a saline (SC) injection and no cue presentation. Tests were conducted in both the acquisition and alternate context.
Statistical analysis
Data in the form of total active and inactive lever responses were analysed using the statistical software program SPSS, and the criterion for statistical significance was set at p < 0.05. All data (active and inactive lever presses) were first analysed with acquisition context treated as a separate factor; if this between-group factor was non-significant, the data were pooled to give two groups for further analysis, extinction in the same or different context as acquisition (AA and AB, respectively). Data from the acquisition phase of nicotine self-administration were analysed by a repeated measures one-way analysis of variance (ANOVA) with session as the within-subject variable. Extinction data were analysed by repeated measures two-way ANOVAs to examine the interaction of the within-subject measure of session with the between-subject variable of extinction context (A or B). A one-way ANOVA and Tukey’s post hoc tests were used to reveal differences between the two groups on specific extinction sessions. Lever pressing during the reinstatement tests was analysed using repeated measures three-way ANOVAs; with the between-subject factor of extinction context (A or B) and the within-subject factors of reinstatement context (A or B) and treatment (nicotine priming and cues or saline and no cues). One-way ANOVAs were followed by pairwise comparisons using the Bonferroni post hoc test to reveal significant within-subject differences and, therefore, the effects of reinstatement treatment and reinstatement context. There was one missing data point for which a harmonic mean was imputed.
Results
Acquisition
Data from the first 13 training sessions were included for analysis, as all animals were yet to enter the extinction phase. The particular context used during training had no significant effect on acquisition of nicotine-seeking behaviour [active lever presses: f(1,50) = 1.513, p = 0.224; inactive lever presses: f(1,50) = 0.152, p = 0.698, two-way ANOVA for repeated measures]; thus, data from the two groups could be combined. Figure 2 shows the acquisition profile for the 52 animals trained to lever press for nicotine, of which 37 successfully reached the stability criteria and thus entered the extinction phase. Over repeated sessions, animals learnt to distinguish the active from the inactive lever and self-administer nicotine, with an increase in active [f(12, 600) = 34.674, p < 0.001] and decrease in inactive [f(12,600) = 3,218, p < 0.001] lever presses.

Acquisition profile of animals (n = 52) trained to self-administer nicotine (data were pooled for those trained either context), showing active and inactive lever presses and infusions over the first 33 days. Days 1–13 (black symbols) include all animals before they reached the stability criteria and thus entered extinction, days 14–33 (white symbols) include the animals that had not yet entered extinction (n = 30–51)
Extinction
Included for analysis were the data from the first three extinction sessions, when all animals were still to reach the extinction criteria and had not yet entered the reinstatement tests. Acquisition context had no significant effect on lever presses [active: f(1,33) = 0.001, p = 0.981; inactive: f(1,33) = 2.548, p = 0.153, three-way ANOVA], and the data were pooled.
Figure 3 shows the 3 days of stable nicotine intake followed by the extinction of nicotine-seeking behaviour in either the acquisition context (n = 18) or alternative context (n = 19). A two-way ANOVA of active lever presses revealed a significant effect of the repeated within-subject measure of session [f(2,66) = 11.694, p < 0.001], the between subject factor of extinction context [f(1,33) = 21.371, p < 0.001] and a significant interaction [f(2,66) = 4.286, p = 0.018]. There was a significant effect of session on the active lever pressing in the three stable sessions of nicotine intake and first three extinction sessions in both groups [f(11,221) = 16.729, p < 0.001, one-way ANOVA]. There were no differences between the two groups in the level of responding in sessions before extinction. However, extinction was initially greater, with lever pressing on the first day being significantly lower (p = 0.006, Tukey’s honestly significantly different), when conducted in a novel environment compared to in the acquisition context, although there were no significant differences between the two groups by day 2 or in the mean number of sessions to reach the extinction criterion.

Extinction profiles (no nicotine infusion and removal of CS) for animals previously trained to self-administer nicotine, showing active and inactive lever presses and infusions. a Extinction in the same context as acquisition (n = 18) and b extinction in the alternative context (n = 19). The dashed line demonstrates commencement of extinction after the three stable days of nicotine self-administration. Black symbols represent the sessions where data from all animals are included, and white symbols represent the sessions in which some animals had reached extinction criteria and thus entered reinstatement tests. Between session differences (A or B; Tukey’s HSD post hoc tests) are represented by an ‘at’ symbol (p < 0.05)
Reinstatement
The reinstatement data showed no significant effect of acquisition context upon responding [active: f(1,29) = 1.086, p = 0.306; inactive: f(1,29) = 1.665, p = 0.207, four-way ANOVA], and the data were once again pooled. Figure 4 shows responses during the four reinstatement tests in the two groups, acquisition and extinction in the same (AA) or different context (AB).

Active and inactive lever presses during the four reinstatement tests: nicotine priming and cues or saline injection and no cues, in context A or B. a Responses in the group in which acquisition and extinction were in the same context (AA) and b responses in the group in which acquisition and extinction were in different contexts (AB). Within-group significant effects (one-way ANOVA and Bonferroni post hoc tests) due to nicotine priming and re-presentation of cues are represented by double asterisks (p < 0.01) and those due to context are represented by double number signs (p < 0.01)
Three-way ANOVAs, with the between-subject factor of extinction context (A or B), the within-subject factors of reinstatement context (A or B) and reinstatement treatment (nicotine with cues or saline without cues), were conducted. For active lever presses, there was a significant interaction between reinstatement context, reinstatement treatment and extinction context [f(1,29) = 4.226, p = 0.049] and significant effects of reinstatement context [f(1,29) = 9.886, p = 0.004] and treatment [f(1,329) = 73.968, p < 0.001], but no effect of the between-group factor of extinction context. No significant effects were found for inactive lever presses.
One-way ANOVAs showed a significant effect of the within-subject factor of reinstatement test on active, but not inactive, lever presses in both groups [AA: f(3,48) = 41.327, p < 0.001; AB: f(3,48) = 15.158, p < 0.001]. Pairwise comparisons using Bonferroni post hoc tests revealed a significant increases in responses due to nicotine priming and re-presentation of cues independent of context (saline − CS compared to nicotine + CS: AAA p < 0.001; AAB p < 0.001; ABA p = 0.007; ABB p = 0.005). However, there was a trend towards a reduction in the reinstatement by nicotine priming and cues when conducted in a novel environment (AAA compared to AAB nicotine + CS), although this did not reach significance in the post hoc tests. When extinction was conducted in a different environment to acquisition, placing animals once again in the acquisition context significantly increased active lever pressing compared to when placed in the extinction context without the need for nicotine priming and re-presentation of cues (ABA compared to ABB: saline − CS p < 0.001).
Discussion
The results presented here indicate involvement of contextual stimuli in both the extinction and reinstatement of nicotine-seeking behaviour. Compared to when conducted in the same context as the self-administration training, extinction was found to be quicker in a novel environment, and there was a trend towards a reduction in the reinstatement of nicotine seeking-behaviour by nicotine priming and re-presentation of cues. In addition, after extinction in an alternative context, re-presentation of the acquisition context associated with nicotine delivery, without nicotine priming or cues, resulted in a significant reinstatement effect.
Extinction training was previously believed to result in unlearning leading to a weakening of the conditioned CS–US association (Mackintosh 1974). This notion has now been replaced by the concept that extinction is a form of new learning in which new associations to the original CS form, i.e. that previously reinforced behaviour is no longer reinforced under the conditions of extinction (Robbins 1990). This is further highlighted by extinction training leading to reduced reinstatement of drug-seeking behaviour compared to an equivalent period of enforced abstinence (Kelamangalath et al. 2007). In the situation presented here, the rate of extinction is likely to have been facilitated by removal of the cues alongside the removal of nicotine (Caggiula et al. 2001). Furthermore, a novel environment rather than the context in which animals had learnt to self-administer nicotine initially resulted in a greater reduction in the level of nicotine-seeking behaviour, although similar levels were reached in subsequent sessions. This may be due to the animal being able to more quickly discriminate when its behaviour is and is not reinforced. These observations suggest that human smokers may find it easier to quit when in a new environment or one not associated with smoking, but as discussed below, this may not be as effective in achieving long-term abstinence.
Reinstatement of nicotine-seeking behaviour, as demonstrated in previous studies (LeSage et al. 2004; Shoaib 2006), was achieved by nicotine priming combined with presentation of cues. There was a trend towards a less robust effect when this reinstatement was conducted in a novel environment compared to the acquisition context, indicating contextual stimuli can interact with drug priming and discrete cues and modify the nicotine-seeking response to these stimuli. The importance of contextual stimuli is further highlighted by the reinstatement of nicotine-seeking behaviour produced by re-presentation of the context in which animals learnt to self-administer nicotine without the need for nicotine priming or cues. The magnitude of responding produced by contextual stimuli is small in comparison to the effects produced by combined nicotine-priming and cue presentation although could probably be made more robust by increasing the number of contextual stimuli. An enhancement of the reinstatement of nicotine-seeking due to nicotine priming and cues in the acquisition context compared to the extinction context was not observed, which may be due to a ceiling effect of the large reinstatement effect produced by nicotine priming and re-presentation of cues.
Spontaneous recovery of nicotine-seeking behaviour has previously been observed when animals were re-exposed to the operant boxes after a period of enforced abstinence (Shaham et al. 1997), and clinical studies have reported craving in abstinent smokers when exposed to environments associated with smoking (Conklin 2006). We have demonstrated a reinstatement of responding by contextual stimuli, suggesting quitting smoking in a novel environment may lead to future vulnerability to relapse when abstinent smokers re-enter a situation in which they previously smoked. These data are similar to reports with other drugs of abuse where exposure to the drug-associated context, after extinction in a different context, renewed drug seeking to a heroin–cocaine mixture (Crombag and Shaham 2002). Furthermore, due the paradoxical nature of nicotine addiction, where reinforcing properties do not fully account for the highly addictive nature of tobacco smoking, environmental stimuli including context may play a particularly important role. Recently, it has been demonstrated that nicotine, in addition to having reinforcing properties, is also capable of enhancing the primary reinforcing value of environmental stimuli in the form of stimulus lights (Donny et al. 2003; Olausson et al. 2004) and it is possible that nicotine may also act to increase the value of more distal cues such as the drug-taking environment.
It must be noted that other studies examining the role of contextual stimuli in relapse (Crombag and Shaham 2002) employ the renewal procedure established by learning laboratories in which CS are presented throughout the extinction process (Bouton and Ricker 1994). In the present paradigm, extinction is achieved by removal of both the unconditioned stimuli and CS. It has been previously stated that contexts alone may not be capable of evoking conditioned responses but are necessary for the expression of learned responses thus setting the stage for the type of responding exhibited (Bouton 1993). However, the data presented here suggest that contextual stimuli are themselves capable of reinstating nicotine-seeking behaviour in the absence of proximal stimuli. Nevertheless, the contextual reinstatement effect using this model is less robust than that produced in studies with other drugs of abuse where CS are also presented. Hence, context is only a contributing factor to the reinstatement of nicotine-seeking behaviour and increased risk of relapse.
Cue exposure, in which addicts are exposed to personally relevant drug cues with the aim of extinguishing learned responses through repeated non-reinforced exposure, is a potentially effective treatment for drug addiction. However, due to its limited clinical success, it has been suggested that efficacy could be improved by employing protocols similar to those used in animal extinction studies. Extinguishing cues in multiple contexts has been found to decrease the context specificity thus increasing the generalisability (Gunther et al. 1998); thus, conducting cue exposure in multiple settings could reduce the association between the unique aspects of the therapy context and extinction and increase long-term abstinence rates (Conklin and Tiffany 2002). The data here suggest that this approach could also improve the long-term success rates of smoking abstinence.
There has been recent investigation into the neurobiology underlying the contextual-reinstatement of drug-seeking, and it is likely that similar brain systems are involved across drugs of abuse. For example, the mesocorticolimbic dopamine system has been implicated in studies using cocaine (Crombag et al. 2002) and heroin (Bossert et al. 2007). However, given the different pharmacological actions of drugs of abuse, specific investigation of contextual modulation of nicotine-seeking behaviour is warranted, and a model such as the one presented in this study could be used to identify the neurotransmitter systems and brain regions involved.
In conclusion, the data here further highlight the important role that environmental stimuli have in nicotine addiction. The drug-taking environment was able to modulate both the extinction and reinstatement of nicotine-seeking behaviour in rodents, and these effects are likely to also occur in human smokers. Thus, context is an additional factor to be targeted when developing smoking cessation strategies.
References
Anthony JC, Warner LA, Kessler RC (1994) Comparative epidemiology of dependence on tobacco, alcohol, controlled substances, and inhalants: basic findings from the national comorbidity survey. Exp Clin Psychopharm 2:244–268
Arroyo M, Markou A, Robbins TW, Everitt BJ (1998) Acquisition, maintenance and reinstatement of intravenous cocaine self-administration under a second-order schedule of reinforcement in rats: effects of conditioned cues and continuous access to cocaine. Psychopharmacology 140:331–344
Baldinger B, Hasenfratz M, Battig K (1995) Switching to ultralow nicotine cigarettes: effects of different tar yields and blocking of olfactory cues. Pharm Biochem Behav 50:233–239
Bossert JM, Liu SY, Lu L, Shaham Y (2004) A role of ventral tegmental area glutamate in contextual cue-induced relapse to heroin seeking. J Neurosci 24:10726–10730
Bossert JM, Poles GC, Wihbey KA, Koya E, Shaham Y (2007) Differential effects of blockade of dopamine D1-family receptors in nucleus accumbens core or shell on reinstatement of heroin seeking induced by contextual and discrete cues. J Neurosci 27:12655–12663
Bouton ME (1993) Context, time, and memory retrieval in the interference paradigms of Pavlovian learning. Psychol Bull 114:80–99
Bouton ME (2004) Context and behavioral processes in extinction. Learn Mem 11:485–494
Bouton ME, Bolles RC (1979) Role of conditioned contextual stimuli in reinstatement of extinguished fear. J Exp Psychol: Animal Behav Proc 5:368–378
Bouton ME, Ricker ST (1994) Renewal of extinguished responding in a second context. Animal Learn Behav 22:317–324
Brauer LH, Behm FM, Lane JD, Westman EC, Perkins C, Rose JE (2001) Individual differences in smoking reward from de-nicotinized cigarettes. Nicotine Tobacco Res 3:101–109
Burattini C, Gill TM, Aicardi G, Janak PH (2006) The ethanol self-administration context as a reinstatement cue: acute effects of naltrexone. Neuroscience 139:877–887
Butschky MF, Bailey D, Henningfield JE, Pickworth WB (1995) Smoking without nicotine delivery decreases withdrawal in 12-hour abstinent smokers. Pharm Biochem Behav 50:91–96
Caggiula AR, Donny EC, White AR, Chaudhri N, Booth S, Gharib MA, Hoffman A, Perkins KA, Sved AF (2001) Cue dependency of nicotine self-administration and smoking. Pharm Biochem Behav 70:515–530
Caggiula AR, Donny EC, Chaudhri N, Perkins KA, Evans-Martin FF, Sved AF (2002a) Importance of nonpharmacological factors in nicotine self-administration. Physiol Behav 77:683–687
Caggiula AR, Donny EC, White AR, Chaudhri N, Booth S, Gharib MA, Hoffman A, Perkins KA, Sved AF (2002b) Environmental stimuli promote the acquisition of nicotine self-administration in rats. Psychopharmacology 163:230–237
Cohen C, Perrault G, Voltz C, Steinberg R, Soubrie P (2002) SR141716, a central cannabinoid (CB(1)) receptor antagonist, blocks the motivational and dopamine-releasing effects of nicotine in rats. Behav Pharmacol 13:451–463
Conklin CA (2006) Environments as cues to smoke: implications for human extinction-based research and treatment. Exp Clin Psychopharm 14:12–19
Conklin CA, Tiffany ST (2002) Applying extinction research and theory to cue-exposure addiction treatments.[see comment]. Addiction 97:155–167
Corrigall WA, Coen KM (1989) Fixed-interval schedules for drug self-administration in the rat. Psychopharmacology 99:136–139
Corrigall WA, Franklin KB, Coen KM, Clarke PB (1992) The mesolimbic dopaminergic system is implicated in the reinforcing effects of nicotine. Psychopharmacology 107:285–289
Crombag HS, Badiani A, Maren S, Robinson TE (2000) The role of contextual versus discrete drug-associated cues in promoting the induction of psychomotor sensitization to intravenous amphetamine. Behav Brain Res 116:1–22
Crombag HS, Grimm JW, Shaham Y (2002) Effect of dopamine receptor antagonists on renewal of cocaine seeking by reexposure to drug-associated contextual cues. Neuropsychopharmacology 27:1006–1015
Crombag HS, Shaham Y (2002) Renewal of drug seeking by contextual cues after prolonged extinction in rats. Behav Neurosci 116:169–173
Donny EC, Caggiula AR, Knopf S, Brown C (1995) Nicotine self-administration in rats. Psychopharmacology 122:390–394
Donny EC, Caggiula AR, Mielke MM, Booth S, Gharib MA, Hoffman A, Maldovan V, Shupenko C, McCallum SE (1999) Nicotine self-administration in rats on a progressive ratio schedule of reinforcement. Psychopharmacology 147:135–142
Donny EC, Caggiula AR, Rowell PP, Gharib MA, Maldovan V, Booth S, Mielke MM, Hoffman A, McCallum S (2000) Nicotine self-administration in rats: estrous cycle effects, sex differences and nicotinic receptor binding. Psychopharmacology 151:392–405
Donny EC, Chaudhri N, Caggiula AR, Evans-Martin FF, Booth S, Gharib MA, Clements LA, Sved AF (2003) Operant responding for a visual reinforcer in rats is enhanced by noncontingent nicotine: implications for nicotine self-administration and reinforcement [see comment]. Psychopharmacology 169:68–76
Epstein DH, Preston KL, Stewart J, Shaham Y (2006) Toward a model of drug relapse: an assessment of the validity of the reinstatement procedure. Psychopharmacology 189:1–16
Fuchs RA, Evans KA, Ledford CC, Parker MP, Case JM, Mehta RH, See RE (2005) The role of the dorsomedial prefrontal cortex, basolateral amygdala, and dorsal hippocampus in contextual reinstatement of cocaine seeking in rats. Neuropsychopharmacology 30:296–309
Goldberg SR, Gardner ML (1981) Second-order schedules: extended sequences of behavior controlled by brief environmental stimuli associated with drug self-administration. NIDA Res Monograph 37:241–270
Goldberg SR, Spealman RD, Goldberg DM (1981) Persistent behavior at high rates maintained by intravenous self-administration of nicotine. Science 214:573–575
Gross J, Lee J, Stitzer ML (1997) Nicotine-containing versus de-nicotinized cigarettes: effects on craving and withdrawal. Pharm Biochem Behav 57:159–165
Gunther LM, Denniston JC, Miller RR (1998) Conducting exposure treatment in multiple contexts can prevent relapse. Behav Res Therapy 36:75–91
Kelamangalath L, Swant J, Stramiello M, Wagner JJ (2007) The effects of extinction training in reducing the reinstatement of drug-seeking behavior: involvement of NMDA receptors. Behav Brain Res 185:119–128
Lazev AB, Herzog TA, Brandon TH (1999) Classical conditions of environmental cues to cigarette smoking. Exp Clin Psychopharm 7:56–63
Le Foll B, Goldberg SR (2005) Nicotine induces conditioned place preferences over a large range of doses in rats. Psychopharmacology 178:481–492
LeSage MG, Burroughs D, Dufek M, Keyler DE, Pentel PR (2004) Reinstatement of nicotine self-administration in rats by presentation of nicotine-paired stimuli, but not nicotine priming. Pharmacol Biochem Behav 79:507–513
Mackintosh NJ (1974) The psychology of animal learning. Academic, London
Markou A, Weiss F, Gold LH, Caine SB, Schulteis G, Koob GF (1993) Animal models of drug craving. Psychopharmacology 112:163–182
Mucha RF, Pauli P, Angrilli A (1998) Conditioned responses elicited by experimentally produced cues for smoking. Can J Physiol Pharm 76:259–268
O’Brien CP, Childress AR, McLellan AT, Ehrman R (1992) Classical conditioning in drug-dependent humans. Ann NY Acad Sci 654:400–415
O’Brien CP, Childress AR, Ehrman R, Robbins SJ (1998) Conditioning factors in drug abuse: can they explain compulsion? J Psychopharm 12:15–22
Olausson P, Jentsch JD, Taylor JR (2004) Nicotine enhances responding with conditioned reinforcement. Psychopharmacology 171:173–178
Pavlov PA (1927) Conditioned reflexes: an investigation of the physiological activity of the cerebral cortex. Oxford Univ. Press, London
Perkins KA, Grobe JE, Fonte C, Goettler J, Caggiula AR, Reynolds WA, Stiller RL, Scierka A, Jacob RG (1994) Chronic and acute tolerance to subjective, behavioral and cardiovascular effects of nicotine in humans. J Pharm Exp Ther 270:628–638
Perkins KA, Gerlach D, Vender J, Grobe J, Meeker J, Hutchison S (2001) Sex differences in the subjective and reinforcing effects of visual and olfactory cigarette smoke stimuli. Nicotine Tobacco Res 3:141–150
Robbins S (1990) Mechanisms underlying spontaneous recovery in autoshaping. J Exp Psychol Anim Behav Proc 16:235–249
Robinson TE, Browman KE, Crombag HS, Badiani A (1998) Modulation of the induction or expression of psychostimulant sensitization by the circumstances surrounding drug administration. Neurosci Biobehav Rev 22:347–354
Rose JE, Levin ED (1991) Inter-relationships between conditioned and primary reinforcement in the maintenance of cigarette smoking. Br J Addiction 86:605–609
Rose JE, Corrigall WA (1997) Nicotine self-administration in animals and humans: similarities and differences. Psychopharmacology 130:28–40
Rose JE, Tashkin DP, Ertle A, Zinser MC, Lafer R (1985) Sensory blockade of smoking satisfaction. Pharm Biochem Behav 23:289–293
Rose JE, Behm FM, Westman EC, Johnson M (2000) Dissociating nicotine and nonnicotine components of cigarette smoking. Pharmacol Biochem Behav 67:71–81
Schenk S, Partridge B (2001) Influence of a conditioned light stimulus on cocaine self-administration in rats. Psychopharmacology 154:390–396
See RE, Grimm JW, Kruzich PJ, Rustay N (1999) The importance of a compound stimulus in conditioned drug-seeking behavior following one week of extinction from self-administered cocaine in rats. Drug Alcohol Depend 57:41–49
Shaham Y, Adamson LK, Grocki S, Corrigall WA (1997) Reinstatement and spontaneous recovery of nicotine seeking in rats. Psychopharmacology 130:396–403, erratum appears in Psychopharmacology (Berl) 1997 Sep;133(1):106
Shaham Y, Shalev U, Lu L, De Wit H, Stewart J (2003) The reinstatement model of drug relapse: history, methodology and major findings [see comment]. Psychopharmacology 168:3–20
Shoaib M (2006) Effects of isoarecolone, a nicotinic receptor agonist in rodent models of nicotine dependence. Psychopharmacology 188:252–257
Shoaib M, Schindler CW, Goldberg SR (1997) Nicotine self-administration in rats: strain and nicotine pre-exposure effects on acquisition. Psychopharmacology 129:35–43
Shoaib M, Sidhpura N, Shafait S (2003) Investigating the actions of bupropion on dependence-related effects of nicotine in rats. Psychopharmacology 165:405–412
Spealman RD, Barrett-Larimore RL, Rowlett JK, Platt DM, Khroyan TV (1999) Pharmacological and environmental determinants of relapse to cocaine-seeking behavior. Pharmacol Biochem Behav 64:327–336
Acknowledgement
We thank the University of Newcastle and Merck, Sharp and Dohme for supporting this research.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Wing, V.C., Shoaib, M. Contextual stimuli modulate extinction and reinstatement in rodents self-administering intravenous nicotine. Psychopharmacology 200, 357–365 (2008). https://doi.org/10.1007/s00213-008-1211-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00213-008-1211-y