
Abstract
Rationale
Abnormalities of glutamatergic neurotransmission have been reportedly observed in psychiatric disorders. Previously, we demonstrated that (1R, 2R, 3R, 5R, 6R)-2-Amino-3-(3,4-dichlorobenzyloxy)-6-fluorobicyclo[3.1.0]hexane-2,6-dicarboxylic acid (MGS0039) is a selective antagonist for group II metabotropic glutamate receptors (mGluR2/3), and that it exerted antidepressant effects in some animal behavioral tests.
Objectives
In the present study, we provide additional evidence that MGS0039 exhibits antidepressant and anxiolytic effects in experimental rodent models, which are predictive of clinical efficacy.
Methods
The learned helplessness (LH) paradigm, which is a common model used to examine the depressive state, was used to assess antidepressant effects of MGS0039. Moreover, anxiolytic effects of MGS0039 were investigated in the conditioned fear stress (CFS) model, which represents emotional abnormality, including anxiety.
Results
Intraperitoneal administration of MGS0039 (10 mg/kg) to rats for 7 days elicited a significant reduction in escape failures in the LH paradigm. In addition, rats treated with MGS0039 (2 mg/kg) showed significantly attenuated freezing behavior in a CFS model, indicating the anxiolytic-like potential of MGS0039.
Conclusions
These results suggest that the blockade of mGluR2/3 with MGS0039 may be effective in the treatment of depressive and anxiety disorders.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Abnormalities of glutamatergic neurotransmission have been observed in several psychiatric disorders such as schizophrenia, attention deficit hyperactivity disorder, bipolar disorder, major depressive disorder, and obsessive–compulsive disorder (Clinton et al. 2003; Carlsson 2001; McCullumsmith and Meador-Woodruff 2002; Rosenberg et al. 2000). Glutamate receptors are classified into two major types: ionotropic glutamate receptors (iGluRs) and metabotropic glutamate receptors (mGluRs). Of these, mGluRs, a family of G-protein-coupled receptors, are divided into three groups based on sequence homologies, their ability to activate specific second messenger systems, and specific sensitivities for agonists (Conn and Pin 1997). Group II mGluRs such as mGluR2 and mGluR3 (mGluR2/3) share significant sequence homologies and are similarly distributed in many brain regions. Various pharmacological and histochemical studies have implicated that these mGluR2/3 play roles in emotional state. For example, mGluR2/3 are abundantly distributed in limbic structures, which participate in the control of emotional states. Moreover, it has been reported that mGluR2/3 agonists exhibited anxiolytic and antipsychotic-like activity in animal models (Helton et al. 1998; Cartmell et al. 1999).
Previously, we reported that (1R, 2R, 3R, 5R, 6R)-2-Amino-3-(3, 4-dichlorobenzyloxy)-6-fluorobicyclo[3.1.0]hexane-2, 6-dicarboxylic acid (MGS0039) is a potent and selective mGluR2/3 antagonist with low affinity for mGluR7, and that MGS0039 exhibited antidepressant effects in rats subjected to forced swimming tests and in mice subjected to tail suspension tests (Chaki et al. 2004). In addition to these studies in behavioral despair models, the chronic administration of MGS0039 promoted cell proliferation in the adult mouse hippocampus (Yoshimizu and Chaki 2004). Recent findings indicate that the behavioral effects of chronic treatment with antidepressants are mediated by the stimulation of neurogenesis in the hippocampus (Santarelli et al. 2003). As above, the blockade of mGluR2/3 might be an effective way of treating subjects suffering from clinical depression.
To ensure clinical efficacy in the treatment of depression, it would be more convincing to demonstrate antidepressant-like potential in additional models that are predictive of the clinical efficacy of compounds. One such model more closely aligned with the clinical aspects of depression is learned helplessness (LH), in which animals are conditioned to fail to escape from a noxious stimulus (Seligman 1978). The LH procedure is a well-established paradigm for studies on the depressive state inasmuch as the escape deficits evoked by inescapable shock are considered to reflect a state of decreased motivation, anhedonia, and psychomotor retardation, key elements of depressive conditions in humans (Seligman and Beagley 1975). Moreover, these escape failures are prevented after chronic, but not acute, treatment with antidepressants (Sherman et al. 1982). This time course of the onset of antidepressant effects in the LH paradigm resembles the actions of the drugs in clinical settings.
In addition to antidepressant-like activity, we previously observed that mGluR2/3 antagonists including MGS0039 exhibited anxiolytic-like activity in a marble-burying model in mice, a model used to evaluate clinical potential in obsessive–compulsive disorder (Shimazaki et al. 2004). However, the anxiolytic-like potential of mGluR2/3 antagonists in animal models largely remains to be explored because MGS0039, like selective serotonin reuptake inhibitors (SSRIs), did not show appreciable effects in classical anxiety models in which only benzodiazepine (BZ) anxiolytics are effective (Chaki et al. 2004). It has been recognized that the conditioned fear stress (CFS) model reflects emotional abnormality including anxiety and depressive state without physical stimuli (Fanselow and Hemstetter 1988; Kalin et al. 1988). Furthermore, the CFS model has been established as a useful model in the prediction of clinical anxiolytic efficacy because not only BZs, but also antidepressants such as SSRIs and serotonin-norepinephrine reuptake inhibitors (SNRIs), are known to be effective. These antidepressants now form the first-line treatment for anxiety disorders (Hashimoto et al. 1996; Inoue et al. 1996).
In the present study, we investigated the effects of MGS0039 in animal models such as LH and CFS, which are predictive of clinical efficacy, to explore possibilities relating to the antidepressant and anxiolytic potential of mGluR2/3 antagonists.
Materials and methods
Experimental animals
Male Sprague–Dawley rats (Charles River, Yokohama, Japan), weighting 250–280 g (for the LH paradigm), or 270–300 g (for the CFS paradigm), were housed in groups of four per cage under standard conditions: room temperature (23±3°C); light–dark cycle (light phase 7:00–19:00). Food and water were available ad libitum. All studies were reviewed by the Taisho Pharmaceutical Animal Care Committee and met the Japanese Experimental Animal Research Association standards, as defined in Guidelines for Animal Experiments (1987).
Learned helplessness paradigm
LH rats were produced and their behaviors assessed using the Shuttle Avoidance System (CompACT SAS/W; Muromachi Kikai, Tokyo, Japan). For the LH paradigm, animals were placed in a shuttle-box (58×24×35.5 cm, with a grid floor) made of two equal-sized compartments divided by an aluminum steel partition fitted with a box, and exposed to 60 times of “inescapable” electric footshocks (intensity 0.4 mA, duration 10 s, interval 10 s) on day 1 of testing. A shock generator scrambler was used to deliver electric shocks to the grid floor. Nonshocked rats were placed in identical chambers for 20 min without receiving electric footshocks. On day 2 of testing, each rat was placed in the same shuttle-box, which was divided into two compartments by a movable partition, and was allowed to habituate to this situation for 5 min and then subjected to “escapable” electric footshocks. This selection session consisted of 40 trials in which electric footshocks (intensity 0.4 mA, duration 10 s, interval 10 s) were preceded by a tone signal (70–75 dB) for 5 s that remained on until the shock was terminated. For each trial, the rat could avoid the shock by crossing into the other compartment of the shuttle box during the tone signal, or terminate the shock by escaping into the other side. Rats received the shock for 10 s when they did not escape into the other compartment (escape failure). Shocks were terminated automatically after 10 s if there was no response within that time. Behaviors were measured and analyzed automatically by CompACT SAS Ver.2 software (Muromachi Kikai, Tokyo Japan). Rats with more than 28 escape failures in the 40 trails were regarded as LH rats. Approximately 75% of the rats fulfilled this criterion. After the selection session, the acceptable rats were grouped through averaging of the number of escape failures for drug administration. As in the case of day 1, nonshocked rats were placed in identical boxes for 20 min without receiving footshocks and tone signals. On day 3 to 9 (for 7 days), MGS0039 (5 or 10 mg/kg), imipramine (30 mg/kg), vehicle (for LH groups), or vehicle (for nonshocked groups) was administered intraperitoneally to rats. On day 9, the avoidance test session was performed under the same conditions as in the selection session on day 2, and the Shuttle Avoidance System recorded the numbers of escape failures. In the shocked group treated with vehicle, LH behavior lasted for 7 days under these conditions.
Conditioned fear stress paradigm
Rats were individually subjected to inescapable electric footshock for a total of 2.5 min [five footshocks (1.0 mA scrambled shock, 30 s duration) that were delivered at intershock mean intervals of 60 s] in a shock chamber with a grid floor of 29×24×30 cm (Neuroscience, Tokyo, Japan). Electric shocks were administered using Model SG-500 (Med Associater, Vermont, USA), which produces a high-voltage, high-resistance circuit with resistance controlled by dial settings calibrated by the manufacturer in a short circuit current. Twenty-four hours after a single footshock session, the rats were placed in a shock chamber without shocks and were observed for 5 min for conditioned fear. Conditioned fear, measured by freezing, develops from the contextual stimuli of the conditioned chamber with these procedures (Fanselow 1980). During the observation period, the duration of freezing behavior was recorded using a modified time-sampling procedure (Fanselow 1980). Freezing was defined as the absence of all observable movement of the skeleton and the vibrissae, except those related to respiration. All other behavior was scored as activity. The animal was classified as showing either freezing or active behavior accordingly.
Chemicals and drug administration
MGS0039 and fluvoxamine were synthesized at Taisho Medicinal Research Laboratories. For the behavioral study, MGS0039 and fluvoxamine were dissolved in 1/15 M phosphate buffer (pH 8.0). Imipramine (Sigma, Tokyo, Japan) was dissolved in saline. Diazepam (Sigma, Tokyo, Japan) was suspended with a 0.3% Tween80/saline solution. In the learned helplessness paradigm, rats were given daily administration of drugs for 7 days. The avoidance test session was carried out 1 h after the last administration. In the CFS paradigm, MGS0039 was administrated intraperitoneally 1 h before the test, and fluvoxamine or diazepam was administrated orally 1 h before the test. Of note, it has been reported that MGS0039 does not affect spontaneous locomotor activity in rats up to 10 mg/kg, and that MGS0039 demonstrates high plasma levels and reaches the brain at pharmacologically relevant concentrations after intraperitoneal administration (Chaki et al. 2004; Nakazato et al. 2004).
Statistics
All statistical analysis was carried out using the PC application SAS (SAS institute Japan, Tokyo, Japan). The statistical significances of differences were determined by Student’s t test or by Dunnett’s test and followed by one-way analysis of variance (ANOVA). The p values of less than 0.05 were considered to be statistically significant.
Results
Effect of MGS0039 on learned helplessness behavior
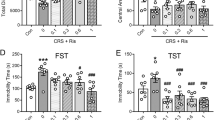
First, the efficacy of imipramine, a well-known antidepressant, in the LH paradigm was examined. Repeated administration of imipramine (10 or 30 mg/kg) for 7 days reduced the number of escape failures in LH rats. Significant effects were observed at 30 mg/kg of imipramine [F(2,23)=13.12, p<0.01]; (number of escape failures±SEM nonshock = 9.4±3.5; vehicle = 37.7±1.0; imipramine 10 mg/kg = 35.8±1.1; 30 mg/kg = 15.0±5.6) (Fig. 1a). After the study on the efficacy of imipramine, the effects of MGS039 on the escape failures in LH rats were examined (Fig. 1b). The number of escape failures in nonshocked rats that were treated with vehicle for 7 days was 15.2±3.4 in the avoidance test session. Compared with these nonshocked rats, shocked rats (vehicle-treated) manifested a significantly higher number of failures (39.0±0.6). In imipramine-treated animals, the number of escape failures was 11.7±3.5, which corresponds to a significant decrease—to the same level as nonshocked rats—compared with the number of escapes in the vehicle-treated group. As for MGS0039, the 3 and 10 mg/kg doses were used in LH rats because 0.3–3 mg/kg of MGS0039 has antidepressant effects in the rat forced swim test (Chaki et al. 2004), and MGS0039 exhibits brain concentrations enough to exert pharmacological effects in rats (Nakazato et al. 2004). Administration of MGS0039 at 10 mg/kg elicited a significant reduction in escape failures as with imipramine (29.5±2.9), though the lower dose of MGS0039 (3 mg/kg) did not exert a significant effect on escape behaviors, showing a tendency to reduce the number of escape failures (33.0±2.2); [F(2,42)=4.99, p<0.05].

Effects of MGS0039 and imipramine on the number of escape failures in learned helplessness rats. MGS0039 (3 or 10 mg/kg) or imipramine (10 or 30 mg/kg) was administrated intraperitoneally for 7 days. Data represent the mean±SEM of the number of escape failures n=14 to 16 per value. Two asterisks indicate p<0.01 vs vehicle treated group (Dunnett’s test). Number sign indicates p<0.01 vs nonshocked group (Student’s t test)
Effect of MGS0039 on conditioned fear stress behavior
The control group (receiving no electrical shock) normally exhibited low freezing behavior, while electrically shocked groups showed freezing behavior characterized by a complete suppression of motility, with occasional defecation and urination. Fluvoxamine, an SSRI, significantly reduced freezing behavior at a dose of 30 mg/kg [F(1,28)=10.48, p<0.01] in the same manner as 3 mg/kg diazepam-treatment [F(3,44)=4.29, p<0.01] (Fig. 2a). The 0.5–2 mg/kg doses of MGS0039 were selected in CFS paradigm based on the results of several behavioral experiments (Chaki et al. 2004; Shimazaki et al. 2004). As shown in Fig. 2b, rats treated with 2 mg/kg of MGS0039 significantly showed attenuated freezing behavior at the same level as a fluvoxamine [F(3,56)=3.29, p<0.05].

Effects of MGS0039, fluvoxamine, and diazepam on freezing in the conditioned fear stress model. Data represent the mean±SEM of freezing time for a 5-min observation period. All drugs were administered intraperitoneally (MGS0039) or orally (fluvoxamine, diazepam) 1 h prior to the test. Tween80 (0.3%)/saline solution and 1/15 M phosphate buffer (pH 8.0) were used as vehicle a and b, respectively. n=12 to 15 per value. Number sign indicates p<0.01 vs control group (Dunnett’s test). *p<0.05 and **p<0.01 vs vehicle treated group (Dunnett’s test)
Discussion
Previously, we showed that MGS0039 is a potent and selective antagonist for mGluR2/3 with low affinity for mGluR7, and that MGS0039 exerts antidepressant effects in behavioral despair models in acute treatment. Thus, MGS0039 can serve as a useful tool in elucidating the role of mGluR2/3 in emotional states. In the present study, we further demonstrated the antidepressant and anxiolytic effects of MGS0039 in LH and CFS models.
The LH model has been widely used for assessment of antidepressant effects of drugs. In this study, a 30-mg/kg dose of imipramine efficiently reduced the number of escape failures in the LH paradigm indicating antidepressant effects, but not a 10-mg/kg dose of imipramine. However, this high dose of imipramine markedly inhibited body weight gain during the drug treatment period (data not shown). As for MGS0039, the treatment at 10 mg/kg for 7 days elicited a significant reduction in escape failures. Although the efficacy of MGS0039 was apparently weaker compared to imipramine (Fig. 1b), MGS0039 had no effects on body weight gain (data not shown). Therefore, this finding suggests that MGS0039 may be effective in the treatment of depressive disorders at doses that are well tolerated.
To date, the involvement of several neurotransmitters in the development, maintenance, prevention, and reversal of LH has been studied. However, the neurochemical mechanisms in LH are complex, and may depend on the specific brain regions studied. Similar to human depression, LH has been associated with a defective serotonergic function. For example, LH rats have a reversible deficit in serotonin (5-HT) release in the prefrontal cortex (Petty and Sherman 1983). Thus, helpless behavior may be a consequence of low 5-HT levels in the brain induced by exposure to inescapable shocks. Chronic antidepressant treatment has been reported to restore this neurochemical imbalance, and to prevent the occurrence of escape failures in the LH model (Petty et al. 1992; Sherman and Petty 1982). This evidence is supported by findings in which repeated administration of SSRIs also prevents LH behavior (Martin et al. 1990), possibly by activation of postsynaptic 5-HT1A receptors (Gambarana et al. 1995). In accordance with this presumption, repeated administration of 5-HT1A-agonists also prevents LH behavior (Graeff et al. 1989). As for 5-HT, our previous studies have interestingly demonstrated that treatment with MGS0039 increased the activity of dorsal raphe nucleus (DRN) serotonergic neurons, and increased extracellular concentration of 5-HT in the medial prefrontal cortex (mPFC) (Kawashima et al. 2005; Karasawa et al. 2005). Therefore, activation of the serotonergic system with MGS0039 possibly contributes to recovery from LH behavior.
It has been suggested that mGluR2/3 negatively regulates dopamine (DA) release in the nucleus accumbens (NAc) shell (Greenslade and Mitchell 2004), and we have recently observed that local application of MGS0039 into the rat NAc shell significantly increases extracellular DA levels in the NAc shell in freely moving rats (Karasawa et al. 2006), indicating that MGS0039 may activate accumbal dopaminergic activity. The involvement of activation of the accumbal dopaminergic system in alleviating the escape deficit in the LH paradigm has been suggested. It has been reported that long-term exposure to inescapable shock develops the escape deficit in rats and reduces DA levels in the NAc shell (Mangiavacchi et al. 2001). Thus, the dysfunction of dopaminergic activity in the NAc shell may be attributed to LH behavior. It has also been shown that local injection of a κ-opioid receptor antagonist into the NAc shell, which increases DA release in the NAc shell, produces antidepressant effects in the LH paradigm (Newton et al. 2002). Although the mechanism by which mGluR2/3 regulates DA release in the NAc shell is unclear, increased DA levels by MGS0039 in the NAc shell may be partly involved in the significant effect of MGS0039 on escape behaviors in the LH paradigm.
Recent findings indicate that the behavioral effects of chronic antidepressants are mediated by the stimulation of neurogenesis in the hippocampus (Santarelli et al. 2003). An intriguing explanation for the length of recovery may be found in the hypothesis that the proliferation and integration of proliferated cells in functional networks play an important role in recovery from depressive episodes. Malberg and Duman (2003) reported that exposure to inescapable shocks in the LH model results in a decrease in cell proliferation in the adult rat hippocampus, and that the administration of fluoxetine, an SSRI, reverses shock-induced reduction in cell proliferation, as well as the behavioral helplessness resulting from exposure to electric shocks. It is interesting to note that chronic administration of MGS0039 promoted cell proliferation in the adult hippocampus, as did administration of different classes of antidepressants (Yoshimizu and Chaki 2004). This effect of MGS0039 might be at least partially implicated in the significant reduction of escape failures in the LH paradigm.
In addition to their efficacious use in treating depressive disorder, SSRIs have been widely used for the treatment of anxiety disorders, for which these compounds have come to form the first-line treatment. Our observations that MGS0039 shares some of the neurochemical mechanisms of SSRIs prompted us to investigate the anxiolytic-like potential of MGS0039 in an animal model of anxiety. The CFS model is regarded as reflecting psychological stress without physical stimuli, and is useful in predicting clinical efficacy, as not only BZ anxiolytics but also SSRIs and SNRIs have been proven to be effective. In the present study, MGS0039 significantly decreased freezing behavior, as did diazepam and fluvoxamine, indicating the anxiolytic-like potential of MGS0039. The precise mechanisms underlying the diminished expression of freezing behavior with MGS0039 remain to be investigated. Freezing behavior induced by CFS has been shown to relate to concentrations of brain 5-HT, particularly in the mPFC and amygdala (Hashimoto et al. 1996, 1999; Inoue et al. 2004). Previously, we found that MGS0039 enhanced the firing rate of serotonergic neurons in the DRN and increased dialysate concentrations of 5-HT in the mPFC (Kawashima et al. 2005; Karasawa et al. 2005). Thus, it is likely that this activation of the serotonergic system may contribute to the anxiolytic-like effects of MGS0039 in the CFS model.
mGluR2/3 inhibit neurotransmitter release as autoreceptors located on glutamatergic terminals. Treatment with the mGluR2/3 antagonists such as MGS0039 in vivo leads to an increase in extracellular glutamate. Therefore, it is an intriguing hypothesis that the moderate elevation of glutamate levels in specific areas of the brain by MGS0039 may cause the anxiolytic-like effects seen in the CFS model. However, there are contradictory findings: decreasing glutamatergic transmission attenuates the expression of conditioned fear. For example, the infusion of N-methyl-d-aspartate, (NMDA) receptor antagonists, or α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) receptor antagonists into the amygdala diminishes the acquisition and expression of fear conditioning (Miserendino et al. 1990; Campeau et al. 1992; Kim et al. 1993; Walker and Davis 1997). In addition, the infusion of LY354740, an mGluR2/3 agonist, into the basolateral amygdala disrupts the fear-potentiated startle response in rats (Stanek et al. 2002). Thus, these findings suggest implication of blockade of glutamatergic transmission in the amygdala in the expression of conditioned fear. NMDA receptors in the amygdala play an important role in triggering the neuronal changes that support fear learning and also the loss of fear that accompanies extinction training, while AMPA receptors also participate in fear learning (Walker and Davis 2002). Given that long-term potentiation, a candidate cellular mechanism for learning and memory in fear conditioning, is mediated by these receptors, then it may be presumed that the impairment of fear-related memory is responsible for the effect of compounds that reduce glutamatergic transmission. Alternatively, given that NMDA receptor antagonists increase glutamate release in certain regions (Moghaddam et al. 1997; Moghaddam and Adams 1998), it could be hypothesized that increase in glutamate release might be responsible for the anxiolytic effects of NMDA receptor antagonists.
Early clinical results with mGluR2/3 agonist LY354740 and its prodrug LY544344 have demonstrated anxiolytic-like efficacy in a model of panic disorder (panic provocation induced by CO2 challenge in subjects with panic disorder) (Schoepp et al. 2003) and in a fear-potentiated startle reflex paradigm in healthy volunteers (Grillon et al. 2003). However, recently, it has been reported that LY354740 failed to give any treatment effects compared to placebo, while there was a clear trend to improvement with paroxetine-treated patients with panic disorder in a multicenter phase II efficacy study (Bergink and Westenberg 2005). Moreover, it has also been reported that LY544344 did not show significant effects on CCK-4-induced anxiety/panic symptoms in entire sample of healthy volunteers (Kellner et al. 2005). Therefore, further studies and discussion may be needed to clarify the potential of mGluR2/3 agonists or antagonists.
In the present study, we demonstrated that MGS0039, an mGluR2/3 antagonist, exhibited antidepressant and anxiolytic effects in rodent models, which can be predictive of clinical therapeutic efficacy. Taking our present and previous findings into consideration, it can be suggested that mGluR2/3 is deeply involved in the etiology of depression and anxiety, and that mGluR2/3 antagonists including MGS0039 may be effective in the treatment of these disorders.
References
Bergink V, Westenberg HG (2005) Metabotropic glutamate II receptor agonists in panic disorder: a double blind clinical trial with LY354740. Int Clin Psychopharmacol 20:291–293
Campeau S, Miserendino MJ, Davis M (1992) Intra-amygdala infusion of the N-methyl-d-aspartate receptor antagonist AP5 blocks acquisition but not expression of fear-potentiated startle to an auditory conditioned stimulus. Behav Neurosci 106:569–574
Carlsson ML (2001) On the role of prefrontal cortex glutamate for the antithetical phenomenology of obsessive compulsive disorder and attention deficit hyperactivity disorder. Prog Neuropsychopharmacol Biol Psychiatry 25:5–26
Cartmell J, Monn JA, Schoepp DD (1999) The metabotropic glutamate 2/3 receptor agonists LY354740 and LY379268 selectively attenuate phencyclidine versus d-amphetamine motor behaviors in rats. J Pharmacol Exp Ther 291:161–170
Chaki S, Yoshikawa R, Hirota S, Shimazaki T, Maeda M, Kawashima N, Yoshimizu T, Yasuhara A, Sakagami K, Okuyama S, Nakanishi S, Nakazato A (2004) MGS0039: a potent and selective group II metabotropic glutamate receptor antagonist with antidepressant-like activity. Neuropharmacology 46:457–467
Clinton SM, Haroutunian V, Davis KL, Meador-Woodruff JH (2003) Altered transcript expression of NMDA receptor-associated postsynaptic proteins in the thalamus of subjects with schizophrenia. Am J Psychiatry 160:1100–1109
Conn PJ, Pin JP (1997) Pharmacology and functions of metabotropic glutamate receptors. Annu Rev Pharmacol Toxicol 37:205–237
Fanselow MS (1980) Conditioned and unconditioned components of postshock freezing. Pavlovian J Biol Sci 15:177–182
Fanselow MS, Hemstetter FJ (1988) Conditional analgesia, defensive freezing, and benzodiazepines. Behav Neurosci 102:233–243
Gambarana C, Ghiglieri O, Taddei I, Tagliamonte A, De Montis MG (1995) Imipramine and fluoxetine prevent the stress-induced escape deficits in rats through a distinct mechanism of action. Behav Pharmacol 6:66–73
Graeff EO, Hunziker MH, Graeff FG (1989) Effects of ipsapirone and BAYR1531 on learned helplessness. Braz J Med Biol Res 22:1141–1144
Greenslade RG, Mitchell SN (2004) Selective action of (−)-2-oxa-4-aminobicyclo[3.1.0]hexane-4,6-dicarboxylate (LY379268), a group II metabotropic glutamate receptor agonist, on basal and phencyclidine-induced dopamine release in the nucleus accumbens shell. Neuropharmacology 47:1–8
Grillon C, Cordova J, Levine LR, Morgan CA 3rd (2003) Anxiolytic effects of a novel group II metabotropic glutamate receptor agonist (LY354740) in the fear-potentiated startle paradigm in humans. Psychopharmacology 168:446–454
Hashimoto S, Inoue T, Koyama T (1996) Serotonin reuptake inhibitors reduce conditioned fear stress-induced freezing behavior in rats. Psychopharmacology 123:182–186
Hashimoto S, Inoue T, Koyama T (1999) Effects of conditioned fear stress on serotonin neurotransmission and freezing behavior in rats. Eur J Pharmacol 378:23–30
Helton DR, Tizzano JP, Monn JA, Schoepp DD, Kallman MJ (1998) Anxiolytic and side-effect profile of LY354740: a potent, highly selective, orally active agonist for group II metabotropic glutamate receptors. J Pharmacol Exp Ther 284:651–660
Inoue T, Tsuchiya K, Koyama T (1996) Serotonergic activation reduces defensive freezing in the conditioned fear paradigm. Pharmacol Biochem Behav 53:825–831
Inoue T, Li XB, Abekawa T, Kitaichi Y, Izumi T, Nakagawa S, Koyama T (2004) Selective serotonin reuptake inhibitor reduces conditioned fear through its effect in the amygdale. Eur J Pharmacol 497:311–316
Kalin NH, Sherman JE, Takahashi LK (1988) Antagonism of endogenous CRH systems attenuates stress-induced freezing behavior in rats. Brain Res 457:130–135
Karasawa J, Shimazaki T, Kawashima N, Chaki S (2005) AMPA receptor stimulation mediates the antidepressant-like effect of a group II metabotropic glutamate receptor antagonist. Brain Res 1042:92–98
Karasawa J, Yoshimizu T, Chaki S (2006) A metabotropic glutamate 2/3 receptor antagonist, MGS0039, increases extracellular dopamine levels in the nucleus accumbens shell. Neurosci Lett 393:127–130
Kawashima N, Karasawa J, Shimazaki T, Chaki S, Okuyama S, Yasuhara A, Nakazato A (2005) Neuropharmacological profiles of antagonists of group II metabotropic glutamate receptors. Neurosci Lett 378:131–134
Kellner M, Muhtz C, Stark K, Yassouridis A, Arlt J, Wiedemann K (2005) Effects of a metabotropic glutamate(2/3) receptor agonist (LY544344/LY354740) on panic anxiety induced by cholecystokinin tetrapeptide in healthy humans: preliminary results. Psychopharmacology 179:310–315
Kim M, Campeau S, Falls WA, Davis M (1993) Infusion of the non-NMDA receptor antagonist CNQX into the amygdala blocks the expression of fear-potentiated startle. Behav Neural Biol 59:5–8
Malberg JE, Duman RS (2003) Cell proliferation in adult hippocampus is decreased by inescapable stress: reversal by fluoxetine treatment. Neuropsychopharmacology 28:1562–1571
Mangiavacchi S, Masi F, Scheggi S, Leggio B, De Montis MG, Gambarana C (2001) Long-term behavioral and neurochemical effects of chronic stress exposure in rats. J Neurochem 79:1113–1121
Martin P, Soubrie P, Puech AJ (1990) Reversal of helpless behavior by serotonin uptake blockers in rats. Psychopharmacology 101:403–407
McCullumsmith RE, Meador-Woodruff JH (2002) Striatal excitatory amino acid transporter transcript expression in schizophrenia, bipolar disorder, and major depressive disorder. Neuropsychopharmacology 26:368–375
Moghaddam B, Adams BW (1998) Reversal of phencyclidine effects by a group II metabotropic glutamate receptor agonist in rats. Science 281:1349–1352
Moghaddam B, Adams B, Verma A, Daly D (1997) Activation of glutamatergic neurotransmission by ketamine: a novel step in the pathway from NMDA receptor blockade to dopaminergic and cognitive disruptions associated with the prefrontal cortex. J Neurosci 17:2921–2927
Miserendino MJ, Sananes CB, Melia KR, Davis M (1990) Blocking of acquisition but not expression of conditioned fear-potentiated startle by NMDA antagonists in the amygdala. Nature 345:708–716
Nakazato A, Sakagami K, Yasuhara A, Ohta H, Yoshikawa R, Itoh M, Nakamura M, Chaki S (2004) Synthesis, in vitro pharmacology, structure-activity relationships, and pharmacokinetics of 3-alkoxy-2-amino-6-fluorobicyclo[3.1.0]hexane-2,6-dicarboxylic acid derivatives as potent and selective group II metabotropic glutamate receptor antagonists. J Med Chem 47:4570–4587
Newton SS, Thome J, Wallace TL, Shirayama Y, Schlesinger L, Sakai N, Chen J, Neve R, Nestler EJ, Duman RS (2002) Inhibition of cAMP response element-binding protein or dynorphin in the nucleus accumbens produces an antidepressant-like effect. J Neurosci 22:10883–10890
Petty F, Sherman AD (1983) Learned helplessness induction decreases in vivo cortical serotonin release. Pharmacol Biochem Behav 18:649–650
Petty F, Kramer G, Wilson L (1992) Prevention of learned helplessness: in vivo correlation with cortical serotonin. Pharmacol Biochem Behav 43:361–367
Rosenberg DR, MacMaster FP, Keshavan MS, Fitzgerald KD, Stewart CM, Moore GJ (2000) Decrease in caudate glutamatergic concentrations in pediatric obsessive–compulsive disorder patients taking paroxetine. J Am Acad Child Adolesc Psychiatry 39:1096–1103
Santarelli L, Saxe M, Gross C, Surget A, Battaglia F, Dulawa S, Weisstaub N, Lee J, Duman R, Arancio O, Belzung C, Hen R (2003) Requirement of hippocampal neurogenesis for the behavioral effects of antidepressants. Science 301:805–809
Schoepp DD, Wright RA, Levine LR, Gaydos B, Potter WZ (2003) LY354740, an mGlu2/3 receptor agonist as a novel approach to treat anxiety/stress. Stress 6:189–197
Seligman ME (1978) Learned helplessness as a model of depression. Comment and integration. J Abnorm Psychol 87:165–179
Seligman ME, Beagley G (1975) Learned helplessness in the rat. J Comp Physiol Psychol 88:534–541
Sherman AD, Petty F (1982) Additivity of neurochemical changes in learned helplessness and imipramine. Behav Neural Biol 35:344–353
Sherman AD, Sacquitne JL, Petty F (1982) Specificity of the learned helplessness model of depression. Pharmacol Biochem Behav 16:449–454
Shimazaki T, Iijima M, Chaki S (2004) Anxiolytic-like activity of MGS0039, a potent group II metabotropic glutamate receptor antagonist, in a marble-burying behavior test. Eur J Pharmacol 501:121–125
Stanek L, Walker DL, Davis M (2002) Amygdala infusion of LY354740, a group II metabotropic receptor agonist, blocks fear-potentiated startle in rats. Soc Neurosci Abstr 26:2020
Walker DL, Davis M (1997) Double dissociation between the involvement of the bed nucleus of the stria terminalis and the central nucleus of the amygdala in startle increases produced by conditioned versus unconditioned fear. J Neurosci 17:9375–9383
Walker DL, Davis M (2002) The role of amygdala glutamate receptors in fear learning, fear-potentiated startle, and extinction. Pharmacol Biochem Behav 71:379–392
Yoshimizu T, Chaki S (2004) Increased cell proliferation in the adult mouse hippocampus following chronic administration of group II metabotropic glutamate receptor antagonist, MGS0039. Biochem Biophys Res Commun 315:493–496
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Yoshimizu, T., Shimazaki, T., Ito, A. et al. An mGluR2/3 antagonist, MGS0039, exerts antidepressant and anxiolytic effects in behavioral models in rats. Psychopharmacology 186, 587–593 (2006). https://doi.org/10.1007/s00213-006-0390-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00213-006-0390-7