
Abstract
The relationship between membrane permeabilization and loss of viability by chilling depending on the chilling rate was investigated in two bacterial models: one Gram-positive bacterium, Lactobacillus plantarum, and one Gram-negative bacterium, Escherichia coli. Cells were cold shocked slowly (2°C/min) or rapidly (2,000°C/min) from physiological temperature to 0°C and maintained at this temperature for up to 1 week. Loss of membrane integrity was assessed by the uptake of the fluorescent dye propidium iodide (PI). Cell death was found to be strongly dependent on the rate of temperature downshift to 0°C. Prolonged incubation of cells after the chilling emphasized the effect of treatment on the cells, as the amount of cell death increased with the length of exposure to low temperature, particularly when cells were rapidly chilled. More than 5 and 3-log reductions in cell population were obtained with L. plantarum and E. coli after the rapid cold shock followed by 7-day storage, respectively. A correlation between cell inactivation and membrane permeabilization was demonstrated with both bacterial strains. Thus, loss of membrane integrity due to the chilling treatments was directly involved in the inactivation of vegetative bacterial cells.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Application of physical stresses to microorganisms is the most widely used method for destruction or stabilization of microorganisms (Marechal et al. 1999). Optimization of such processes is the objective of both industry and scientists. Preservation of foods using low temperatures is popular and practiced worldwide, and examples of this are chilling and freezing, which are the most commonly used for food processing. Chilling generally refers to temperatures above 0°C, at which the rate of growth of microorganisms is slowed down. Exposure of microorganisms to low positive temperatures is usually associated with a cellular metabolic adaptive response (Jones and Inouye 1994), contrary to freezing temperatures that are well known to inhibit metabolic activity and cell growth for most organisms (Panoff et al. 1998). At present, the effects of the rates of cooling have been studied and applied only to the freezing process, for which very low (<5°C/min) and intermediate cooling rates (180–5,000°C/min) induced noticeable cell injuries or death, whereas low (5–180°C/min) or very high cooling rates (>5,000°C/min) were found to optimize cell survival (Dumont et al. 2004; Mazur 1970). With regard to chilling, the chilling rates in non-freezing temperature range have often been neglected and consequently remained unexplored fundamentally. It has been recently shown that chilling could induce cell inactivation if cells are rapidly chilled at critical chilling rate (Cao-Hoang et al. 2008a). Membrane alterations seemed to be the principal cause of the cold shock injury during the rapid temperature variation. The kinetics of temperature decrease to 0°C appears thus an important criterion for cell preservation or cell inactivation, depending on the degree of membrane damage. In this paper, we investigated the effect of a lethal chilling (2,000°C/min) and a slow chilling (2°C/min) followed by cold storage on the inactivation of the Gram-negative bacterium Escherichia coli TG 1 and the Gram-positive bacterium Lactobacillus plantarum 103151T. The relationship between cell death and membrane permeabilization induced by cold shock was also studied to better understand the mechanisms leading to the cell death, by measuring the uptake of the fluorescent probe propidium iodide (PI). It has been shown that this probe is a good indicator of membrane integrity as it is a nucleotide-binding probe excluded by intact cells (Wouters et al. 2001). Indeed, PI uptake was successfully used to membrane permeabilization measurement by flow cytometry in relation with inactivation kinetics of different microbial strains after various treatments (Aronsson et al. 2005; Pagan and Mackey 2000; Ulmer et al. 2000; Wouters et al. 2001).
Materials and methods
Microbial materials
Escherichia coli strain TG1 (K12 supEhsdΔ5 thi Δ (lac-proAB) F’[traD36 proAB+lacIqlacZΔM15]) was from the Microbiology Laboratory Culture Collection, ENSBANA, Dijon, France. L. plantarum strain 103151T was obtained from the Collection of Pasteur Institute, Paris, France.
E. coli was maintained on Luria Broth (LB) agar plates (Sigma–Aldrich, St. Louis, MO, USA) at 4°C. Subculture was prepared by transferring a colony from each plate into 10 ml of LB broth at water activity (a w ) equal to 0.992, followed by incubation for 18 h at 37°C. A culture was then prepared by inoculating 0.1 ml of bacterial suspension into 10 ml of the respective growth medium and incubated statically to mid-log growth phase (final population of 2 × 107 cells/ml).
L. plantarum was maintained on Petri dishes with De Man, Rogosa, Sharpe (MRS) (Sigma–Aldrich) agar at 4°C. Cultures were grown in 10 ml of MRS broth (a w = 0.992) followed by incubation at 30°C. Inoculation was carried out by using 0.1 ml of bacterial suspension from a 18-h subculture grown under analogous conditions until the mid-log growth phase was reached (final population of 1 × 108 cells/ml). The growth phases of the microbial cultures were estimated from the scatter at 600 nm (A600).
Chilling treatments
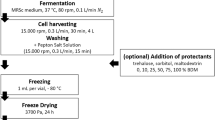
Chilling was conducted in phosphate-buffer saline (PBS) and performed as described previously (Cao-Hoang et al. 2008a). Cells were collected by centrifugation at 2,800g for 10 min. The pellets were washed and resuspended in PBS to a cell concentration of about 107 CFU/ml (A 600 = 0.6). Cells were then submitted to chilling from T i (growth temperature: T i = 37°C for E. coli or 30°C for L. plantarum) to 0°C at two chilling rates 2°C/min and 2,000°C/min as described in the previous paper (Cao-Hoang et al. 2008a). The temperature variation was followed using a T-thermocouple (TCSA, Dardilly, France) inserted into an uninoculated tube (blank) that was placed into the cryostat along with the inoculated samples. When the target temperature of 0°C was obtained, samples were either transferred directly to a water-bath at T i for warming at 2°C/min or incubated at 0°C for several days up to 1 week before viability determination. The experimental design is schematically represented in Fig. 1.

Schematic representation of the experimental design
Viable cell counts
Cell viabilities were determined immediately after chilling treatment or after 1-, 3- and 7-day incubation in PBS buffer at 0°C. Cell suspensions were adequately diluted then plated simultaneously onto nutritive media and incubated at 37°C for 24 h for E. coli and 30°C for 48 h for L. plantarum. After incubation, colony forming units (CFU) were counted. Viabilities were determined by comparing the experimental and control CFU counts. Each experiment was repeated at least three times.
Propidium iodide (PI) staining for cell membrane integrity by fluorescence measurement
Propidium Iodide (PI) from Molecular Probes (Invitrogen SARL, Cergy Pontoise, France) was used to evaluate the effect of chilling treatment on cell membrane integrity. This probe is a nucleotide-binding probe excluded by intact cells, and it cannot penetrate an intact cell membrane. It stains the DNA of the cells having lost their membrane integrity. Cells were stained with PI either before the treatment or after the treatment. Changes in membrane permeability were also followed during incubation period at 0°C. Before treatments, cells were harvested by centrifugation (2,800g, 10 min) and washed twice in PBS. The cell pellets were resuspended in the same buffer, and the cell concentration was adjusted to A 600 = 0.2. PI was prepared in distilled water and added to cell suspensions to a final concentration of 2.9 μM (Pagan and Mackey 2000). After 10-min incubation in the dark, cell suspensions were centrifuged and washed twice to ensure that no extracellular PI remained in the buffer. Fluorescence was measured using a Fluorolog-3 spectrofluorometer (Jobin–Yvon, Horiba Group, Edison, Inc., USA) using a 535-nm excitation filter and a 625-nm emission filter. The slit width was 10 nm. When cells were stained with PI after treatments, PI was added to cell suspensions to a final concentration of 2.9 μM, and fluorescence was measured. Changes in membrane permeability were expressed as PI uptake, so-called the degree of permeabilization, which corresponds to the ratio of fluorescent intensity value of treated and untreated cells. All fluorescence measurements were performed at least in triplicate from independent cultures. Means and 95% confidence intervals were calculated.
Results
Effect of chilling treatments on cell survival
The effect of different kinetics of chilling from optimal growth temperature to 0°C was assessed on the survival of two bacteria, one Gram-positive bacterium, L. plantarum, and one Gram-negative bacterium, E. coli. Figure 2a, b show the bacterial survival fraction after the slow chilling of 2°C/min and the rapid chilling of 2,000°C/min followed by 0°C-storage, respectively.

Effect of the chilling at a 2°C/min and b 2,000°C/min followed by prolonged incubation at 0°C on bacterial survival of E. coli and L. plantarum. Error bars show the standard deviation of the mean of at least three replicates
The slow chilling alone, i.e. without low temperature storage, did not affect cell survival for both bacterial strains, as only about 13 and 3% of loss of viability were recorded for E. coli and L. plantarum, respectively (Fig. 2a). However, when cells were incubated at 0°C after the slow chilling treatment, the total bacterial population decreased progressively during the first 3-day incubation to reach 55% for E. coli and 26% for L. plantarum, thereafter remaining stable at 0°C until day 7. The rapid chilling at 2,000°C/min was more effective compared with the slow chilling with regard to E. coli, as a 1-log reduction was observed with this bacterium immediately after the shock. However, no significant reduction in bacterial counts was noticed for L. plantarum (Fig. 2b). Interestingly, cell viability decreased rapidly upon storage at 0°C, and the inactivation rate of L. plantarum cells was higher than that of E. coli cells. About 5.5-log and 3.8-log reductions in the number of survivors were observed within 7 days at 0°C with L. plantarum and E. coli, respectively (Fig. 2b).
Bacterial membrane permeabilization as a function of chilling rate during treatment
From the previous results, it appeared that chilling to 0°C sensitized both E. coli and L. plantarum, and the greater the chilling rate was, the higher the percentage of cell death obtained. Furthermore, incubation at 0°C after chilling seemed to increase the effect of chilling on cell survival, as loss of viability occurred during low temperature storage. In order to evaluate the effect of chilling on the membrane physical state, membrane permeabilization of both bacterial strains was evaluated by measuring the PI fluorescence and the amount of PI uptake by spectrofluorometer. Table 1 shows the effect of the slow chilling (2°C/min) and the rapid chilling (2,000°C/min) on PI fluorescence of treated cells when PI was added before or immediately after the chilling treatments. The slow chilling did not cause uptake of PI into both cells of E. coli and L. plantarum in comparison with untreated cells, as evaluated by PI fluorescence, suggesting that the bacterial membrane remained intact after this treatment. Concerning the rapid chilling, significant increase in PI fluorescence was observed with both bacterial strains, particularly with L. plantarum, where PI fluorescence of treated cells was about 20-fold and 15-fold higher than that of untreated cells when PI was present during or after the chilling, respectively, confirming membrane damage due to the rapid cold shock (Table 1).
Changes in membrane permeability of treated cells were also followed during the incubation period at 0°C. Figure 3a, b show, respectively, the membrane permeabilization of E. coli and L. plantarum, as evaluated by PI uptake, after the slow chilling at 2°C/min followed by incubation at 0°C. The figures also include the percentage of cell survival upon the treatments. A Slight increase in PI fluorescence value was observed with L. plantarum when the cells were maintained at 0°C after the slow chilling (Fig. 3b), whereas cells of E. coli maintained a low degree of membrane permeabilization upon storage (Fig. 3a). The relationship between membrane permeabilization and cell inactivation of E. coli and L. plantarum after the rapid chilling at 2,000°C/min followed by storage at 0°C is illustrated in Fig. 3c, d, respectively. Both cells of E. coli and L. plantarum showed a gradual loss of membrane integrity, as reflected by the increase in PI uptake (Fig. 3c, d) when maintained at 0°C after the rapid chilling. Interestingly, the PI uptake was much greater for L. plantarum, and the fluorescence intensity remained approximately 120-fold higher than that of untreated control at day 7 of storage (Fig. 3d), whereas for E. coli, it was 30-fold higher than that of untreated cells (Fig. 3c).

Relationship between membrane permeabilization and microbial survival after chilling treatments. a Chilling E. coli at 2°C/min; b Chilling L. plantarum at 2°C/min; c Chilling E. coli at 2,000°C/min; d Chilling L. plantarum at 2,000°C/min
Bacterial membrane permeabilization was plotted against log reduction in order to investigate whether the observed decrease in viability of two strains was directly caused by membrane permeabilization (Fig. 4a, b). We found that increase in cell inactivation did not necessarily imply greater PI uptake in the case of the slow chilling (Fig. 4a), while there appeared to be a linear relationship between the number of inactivated cells and the number of permeabilized cells when cells were cold shocked rapidly following by incubation at 0°C, and it was for both E. coli and L. plantarum (Fig. 4b). These results showed a positive correlation between membrane damage, as measured by PI uptake, and the loss of viability induced by the rapid cold shocked cells.

Bacterial membrane permeabilization as a function of inactivation of E. coli and L. plantarum after a slow chilling at 2°C/min and b rapid chilling at 2,000°C/min. N 0 , number of cells before treatment; N t , number of cells after treatment
Discussion
In this work, we have investigated the effect of slow and rapid chilling followed by low temperature storage on the survival of one Gram-positive bacterium (L. plantarum) and one Gram-negative bacterium (E. coli), as well as the relationship between cell inactivation and membrane permeabilization due to the treatments. Numerous works have reported the occurrence of membrane permeabilization under different treatments such as pulsed electric fields (Aronsson et al. 2005; Garcia et al. 2007; Pakhomov et al. 2007; Unal et al. 2001; Wouters et al. 2001), high hydrostatic pressure (Benito et al. 1999; Manas and Mackey 2004; Moussa et al. 2007; Pagan and Mackey 2000), osmotic pressure (Simonin et al. 2007) or freezing (Kaidi et al. 2001; Medrano et al. 2002; Reid et al. 1999) using PI as detector of membrane damage. However, little was known about the effect of chilling and chilling rates on loss of microbial viability and its relation with loss of membrane integrity. The relationship between irreversible membrane permeabilization and inactivation by pulsed electric fields has been observed in both Gram-positive and Gram-negative bacteria (Garcia et al. 2007; Unal et al. 2001; Wouters et al. 2001). Garcia et al. (2007) have correlated loss of viability with a permanent loss of membrane integrity of pulsed electric field-treated cells. Leslie et al. (1994), Simonin et al. (2007) have related the cell death during dehydration to changes in plasma membrane permeability. In our present investigation, it was shown that the slow chilling of 2°C/min did not significantly affect cell viability of both L. plantarum (Gram positive) and E. coli (Gram negative) (c.f. Fig. 2a, day 0), and neither renders the membrane permeabilized to PI. Gradual loss in cell viability occurred when treated cells were maintained at low temperature after the slow chilling; approximately, 45 and 75% reductions in cell population were achieved after 7-day incubation at 0°C with E. coli and L. plantarum, respectively (Fig. 2a). However, cell membrane remained apparently intact as very little permeabilization assessed by PI uptake was observed (Fig. 3a, b). As the cells were maintained in buffer after chilling, this decrease in viability might thus simply be attributed to the lack of nutrients in the incubation medium.
With regard to the rapid chilling of 2,000°C/min, response of cells of E. coli and of L. plantarum appeared to be different depending on whether the cells were hold at 0°C after the rapid chilling or not. The rapid chilling alone resulted in 1-log decrease in E. coli population; then, cell death increased as the period of incubation at 0°C was extended, reaching more than 3-log inactivation at day 7 (Fig. 2b). Contrary to E. coli cells, cells of L. plantarum seemed resistant to the rapid cold shock without post-incubation, as no loss of viability occurred at day 0; however, they showed high sensitivity to the rapid chilling followed by low temperature storage as more than 5-log reductions in bacterial counts were obtained at day 7 (Fig. 2b). Interestingly, whether PI was added before or after the treatment, greater PI uptake was seen in L. plantarum than in E. coli at day 0 (Table 1). These results suggest that cells of L. plantarum had not necessarily lost their ability to multiply after the rapid chilling to 0°C if cells were immediately returned to initial conditions, although their cell membranes were permeabilized, whereas cells of E. coli did not have this capacity. However, when maintaining L. plantarum and E. coli treated cells at 0°C after the rapid cold shock, both showed gradual loss of viability upon incubation, which correlated with the loss of membrane integrity as evaluated by PI uptake (Fig. 3c, d). This indicated that (1) this rate of temperature decrease may be critical for cell viability as well as membrane permeability during chilling and (2) membrane was permanently permeabilized, as cells were stained with PI after treatment. It was shown previously that rapid temperature downshift could induce phase separation of phospholipids, leading to increase in membrane permeability (Cao-Hoang et al. 2008a). The instantaneous chilling could lead to crystallization of some phospholipid-containing compounds in the bacterial membranes, with formation of crystals that puncture the membranes with increase in membrane permeability and death as a subsequent possibility. Furthermore, by plotting PI uptake versus bacterial log reduction induced by the rapid chilling, we obtained a linear relationship between increased PI uptake and an increased level of inactivation (Fig. 4b). These results are in agreement with those obtained previously (Cao-Hoang et al. 2008a, b), confirming the destructive effect of the rapid chilling on the cell membrane and that the membrane permeabilization observed could be critical for bacterial survival. From the results of the present study, we propose a mechanism leading to the cell death during chilling followed by incubation at reduced temperatures as follows: when cells are chilled at a critical rate, their membrane is altered and undergoes structural changes such as phospholipid phase separation, leading to increase in membrane permeability and consequently leading to immediate cell mortality or time-induced death during prolonged incubation period at low temperature if the cells are not able to survive permeabilization. Following incubation, these cells, suffering membrane damage, lose cellular materials (e.g. leakage of ATP, UV-absorbing substances) as their membrane cannot reseal, consequently leading to increased cell inactivation.
The membrane of bacterial cells, particularly of the Gram-positive bacterium L. plantarum, was shown to be drastically permeabilized after the rapid chilling followed by 7-day incubation, as PI fluorescence intensities of treated cells were 120-fold higher than those of untreated cells (Fig. 3d). Besides, cells of L. plantarum seemed to exhibit higher degree of membrane permeabilization than that of E. coli for a same percentage of inactivated cells obtained (Fig. 4b). In other words, E. coli was found to be more chilling resistant and showed less membrane permeabilization than the more chilling-sensitive bacterium, L. plantarum. Thus, membrane permeabilization is probably important in determining chilling inactivation. These results were in agreement with those observed with cells treated by pulsed electric fields that Gram-positive bacterial membrane was more permeabilized than Gram-negative bacterial membrane when submitted to the same treatment (Garcia et al. 2007). Indeed, Gram-negative bacteria are bounded by two membranes, outer membrane and cytoplasmic membrane, while Gram-positive bacteria are surrounded only by one single cytoplasmic membrane (Clark 2005). Thus, the outer membrane of Gram-negative bacteria must be first permeabilized to allow molecules such as PI to gain access to the cytoplasmic membrane. This could explain the difference in PI fluorescence intensities obtained with two bacterial species.
In conclusion, we have described for the first time important differences about the way that chilling affects the cellular membrane and that membrane damage may depend on the intensity and the duration of the chilling treatment. Loss of viability appears to be directly related to a permanent loss of membrane integrity induced by the rapid cold shock. It is remarkable that cold-shocked cells maintained at low temperatures lost viability during storage time as a function of the chilling rate. Recently, combination of rapid chilling with nisin, a polypeptide produced by lactic acid bacteria that is recognized as natural, toxicologically safe, antimicrobial food additive (Cleveland et al. 2001), was shown to be effective against the Gram-negative bacterium E. coli as it resulted in more than 6-log reduction in bacterial population (Cao-Hoang et al. 2008b). Thus, utilization of chilling in adequate conditions such as in combination with nisin or other bacteriocins or following by low temperature incubation in food processing treatments could increase the efficiency of the process, enhancing microbiological safety and stability. Finally, more knowledge is required about the mechanisms underlying chilling sensitivity, so that conditions under which microorganisms can be optimally inactivated can be designed.
References
Aronsson K, Rönner U, Borch E (2005) Inactivation of Escherichia coli, Listeria innocua and Saccharomyces cerevisiae in relation to membrane permeabilization and subsequent leakage of intracellular compounds due to pulsed electric field processing. Int J Food Microbiol 99(1):19–32
Benito A, Ventoura G, Casadei M, Robinson T, Mackey B (1999) Variation in resistance of natural isolates of Escherichia coli O157 to high hydrostatic pressure, mild heat, and other stresses. Appl Environ Microbiol 65(4):1564–1569
Cao-Hoang L, Dumont F, Marechal PA, Le-Thanh M, Gervais P (2008a) Rates of chilling to 0 degrees C: implications for the survival of microorganisms and relationship with membrane fluidity modifications. Appl Microbiol Biotechnol 77(6):1379–1387
Cao-Hoang L, Marechal P, Le-Thanh M, Gervais P (2008b) Synergistic action of rapid chilling and nisin on the inactivation of Escherichia coli. Appl Microbiol Biotechnol 79(1):105–109
Clark D (2005) Molecular biology. Academic Press, Sandiego, p 784
Cleveland J, Montville TJ, Nes IF, Chikindas ML (2001) Bacteriocins: safe, natural antimicrobials for food preservation. Int J Food Microbiol 71(1):1–20
Dumont F, Marechal PA, Gervais P (2004) Cell size and water permeability as determining factors for cell viability after freezing at different cooling rates. Appl Environ Microbiol 70(1):268–272
Garcia D, Gomez N, Manas P, Raso J, Pagan R (2007) Pulsed electric fields cause bacterial envelopes permeabilization depending on the treatment intensity, the treatment medium pH and the microorganism investigated. Int J Food Microbiol 113(2):219–227
Jones PG, Inouye M (1994) The cold-shock response: a hot topic. Mol Microbiol 11(5):811–818
Kaidi S, Bernard S, Lambert P, Massip A, Dessy F, Donnay I (2001) Effect of conventional controlled-rate freezing and vitrification on morphology and metabolism of bovine blastocysts produced in vitro. Biol Reprod 65(4):1127–1134
Leslie SB, Teter SA, Crowe LM, Crowe JH (1994) Trehalose lowers membrane phase transitions in dry yeast cells. Biochim Biophys Acta 1192(1):7–13
Manas P, Mackey BM (2004) Morphological and physiological changes induced by high hydrostatic pressure in exponential- and stationary-phase cells of Escherichia coli: relationship with cell death. Appl Environ Microbiol 70(3):1545–1554
Marechal PA, Martinez de Marañon I, Porrier I, Gervais P (1999) The importance of the kinetics of application physical stresses on the viability of microorganisms: significance for minimal food processing. Trends Food Sci Tech 10:15–20
Mazur P (1970) Cryobiology: the freezing of biological systems. Science 168(934):939–949
Medrano A, Watson PF, Holt WV (2002) Importance of cooling rate and animal variability for boar sperm cryopreservation: insights from the cryomicroscope. Reproduction 123(2):315–322
Moussa M, Perrier-Cornet J-M, Gervais P (2007) Damage in Escherichia coli cells treated with a combination of high hydrostatic pressure and subzero temperature. Appl Environ Microbiol 73(20):6508–6518
Pagan R, Mackey B (2000) Relationship between membrane damage and cell death in pressure-treated Escherichia coli cells: differences between exponential- and stationary-phase cells and variation among strains. Appl Environ Microbiol 66(7):2829–2834
Pakhomov AG, Shevin R, White JA, Kolb JF, Pakhomova ON, Joshi RP, Schoenbach KH (2007) Membrane permeabilization and cell damage by ultrashort electric field shocks. Arch Biochem Biophys 465(1):109–118
Panoff J-M, Thammavongs B, Gueguen M, Boutibonnes P (1998) Cold stress responses in mesophilic bacteria. Cryobiology 36(2):75–83
Reid TJ, LaRussa VF, Esteban G, Clear M, Davies L, Shea S, Gorogias M (1999) Cooling and freezing damage platelet membrane integrity. Cryobiology 38(3):209–224
Simonin H, Beney L, Gervais P (2007) Sequence of occurring damages in yeast plasma membrane during dehydration and rehydration: mechanisms of cell death. Biochim Biophys Acta 1768(6):1600–1610
Ulmer HM, Ganzle MG, Vogel RF (2000) Effects of high pressure on survival and metabolic activity of Lactobacillus plantarum TMW1.460. Appl Environ Microbiol 66(9):3966–3973
Unal R, Kim JG, Yousef AE (2001) Inactivation of Escherichia coli O1 57:H7, Listeria monocytogenes, and Lactobacillus leichmannii by combinations of ozone and pulsed electric field. J Food Prot 64(6):777–782
Wouters PC, Bos AP, Ueckert J (2001) Membrane permeabilization in relation to inactivation kinetics of Lactobacillus species due to pulsed electric fields. Appl Environ Microbiol 67(7):3092–3101
Acknowledgments
This work was supported in part by the Agence Universitaire de la Francophonie (AUF), the Vietnamese and French Ministries of Education and Training and the Bourgogne region.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Erko Stackebrandt.
Rights and permissions
About this article
Cite this article
Cao-Hoang, L., Dumont, F., Marechal, P.A. et al. Inactivation of Escherichia coli and Lactobacillus plantarum in relation to membrane permeabilization due to rapid chilling followed by cold storage. Arch Microbiol 192, 299–305 (2010). https://doi.org/10.1007/s00203-010-0555-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00203-010-0555-y