
Abstract
In order to improve the knowledge of total mercury (THg) bioaccumulation in bioindicator species of sea catfish (Siluriformes, Ariidae) and taking into account the relatively recent approach with respect to estuarine fish on the Brazilian coast, 65 individuals were caught in the northern and southern regions of the Cananeia estuary to determine the concentration of the THg in muscles, gills, gonads and kidney of the Cathorops spixii and Genidens genidens specimens. The difference in the THg accumulation associated to the maturity of the catfish reflects a differential metabolism regarding THg bioaccumulation in adults (males and females) and juveniles. These observations reinforce the importance of considering the maturity of the individual in order to understand the bioaccumulation and metabolism of fish under different environmental stress and conditions. Furthermore, abiotic conditions such as salinity should be evaluated in association with metabolic/biological conditions of the fish’s bioindicators, especially in environments with large natural or anthropogenic transition gradients.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
In the bioaccumulation process, the chemical compounds are retained in different organs/tissues depending on their chemical form and biochemistry interactions. Therefore, bioaccumulation of metals such as mercury (Hg) can cause metabolic disruptions and acute and/or chronic consequences. Hg can occur in the environment in both organic and inorganic forms, in different proportions and exert potent neurotoxic effects on the biota (Heath 1990; Lacerda and Malm 2008). Therefore, this element has great capacity for bioaccumulation and biomagnification, to the marine environment (Dias et al. 2008).
The somatic indexes, the hepatosomatic index (HSI) in particular and condition factors (CF), are important to describe additional information concerning damage to the liver, hepatic atrophy and the degree of well-being (Adams and McLean 1985; Gomiero and Braga 2003). Besides evaluating biological changes, such as reproduction (Vazzoler 1996), it is possible to evaluate the impact of abiotic conditions of the aquatic systems in fish species since they are poikilothermic organisms.
To evaluate the bioavailability of metals in biota, it is important to use bioindicators that are relevant for ecological, environmental, and biodiversity importance. Sea catfish (Siluriformes, Ariidae) were suggested as a bioindicator species for metal contamination (Azevedo et al. 2012a) and is a regionally relevant representative of an ecological and environmental bioindicator.
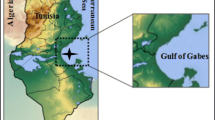
The Cananéia-Iguape estuarine-lagoon complex (CIELC) is an aquatic ecosystem subject to minimal anthropogenic influence, considered a Biosphere Reserve (UNESCO). Recently, it was included on the Ramsar list of wetlands of international importance (https://rsis.ramsar.org/ris/2310). Thus, the monitoring of Hg and its effect on the metabolism of bioindicator species is recommended in this area mainly because other studies have indicated the presence of Hg in sediments of the northern region of this system (Amorim et al. 2008). This area receives Ribeira de Iguape River inflow via Valo Grande (artificial channel) with materials from continental and anthropogenic origins. The southern region is more preserved than the northern area of the CIELC, since it receives major influence from the ocean, therefore presenting more estuarine features naturally attributed to the tides and the sea hydrodynamic (Fig. 1). Azevedo et al. (2012b) recommended the evaluation of biological variables such as the length, weight, and liver weight of the fish, enabling the calculation of the somatic indices; CF and HSI, in order to understand the total mercury (THg) bioaccumulation in fish subjected to anthropogenic and natural environmental conditions. This study aimed to improve the knowledge of the bioaccumulation process of THg in the muscles, gills, gonads and kidneys of the Ariidae catfish Cathorops spixii and Genidens genidens, observing HSI and CF indices with consideration towards a sexual pattern and the maturity phase, e.g. adults (male and female) and juvenile.

Sampling sites showing the southern and northern regions in the CIELC, São Paulo State, Brazil
Materials and Methods
The bottom water samples were obtained in the same sites of the catfish collection, but before the Ariidae catch. For the water sampling a van Dorn bottle was used. The salinity of the bottom water was determined by inductive method, using Beckman® salinometer, model RS-10, with an accuracy of ± 0.001. The equipment was calibrated using ampoules of standard seawater.
A total of 65 fish were collected during the winter of 2014 (August 26) in the northern (coordinates: 24°54′S, 47°48′W) (n = 32) and southern (coordinates: 25°02′S, 47°54′W) (n = 33) regions of CIELC (SP, Brazil) (Fig. 1). The fish were caught using an “Otter Trawl” bottom trawls at a mean depth of 6 m for 10 min at 3 mph. Immediately, the catfish were separated and the species identified according to Figueiredo and Menezes (1978). The fish were transported to the laboratory in an icebox and the biometric data, such as total length (TL), total weight (TW), body weight (BW), and liver weight (LW) were obtained in accordance with Vazzoler (1996), with posterior dissection of each individual organism for extraction of muscle, gill, gonad and kidney. The obtained tissues were identified, packed in zippered bags and stored at − 4°C until the time of THg analysis.
TW, TL, BW and LW were used to calculate somatic indices: Fulton CF as BW/TL3 and HSI as LW/BW × 100.
Muscle, gill, gonad and kidney samples were stored at − 20°C until THg analysis. Each sample was weighed according to the type of tissue, for instance: gill: 0.1–0.2 g, gonads: 0.01–0.1 g, muscle: 0.8–1.0 g, and kidney: 0.1–0.2 g and placed in a nickel sample. The THg determination was made by solid thermal decomposition using a Direct Mercury elementary analyzer (DMA-80 Milestone). The analytical control was made by the simultaneous analysis of the certified material DORM-4 (National Research Council Canada – NRC). The obtained detection and quantification limits were 0.408 ng g−1 and 3.619 ng g−1, respectively, with circa 90% recovery compared with DORM-4, showing high precision and accuracy of the THg determination. Results of THg were expressed in wet weight.
Initially, species (C. spixii and G. genidens) were evaluated concerning differences in THg concentrations and somatic indices. However, as no statistically significant differences were obtained, species were then grouped in order to increase the sample number. No females were collected in the northern region. Differences in the somatic indices (HSI and CF) were tested by ANCOVA and TL were used as co-variable to CF. Differences in THg concentration in the catfish tissues were tested by the non-parametric median test using a Kruskal–Wallis test with Dunn’s multiple comparisons test using all groups available (males/females/juvenile, and Northern/Southern areas). Regarding the THg levels, principal components analysis (PCA) was used in order to verify the dependence of the each variable in each sampling site and the hierarchical clustering analysis was also used in order to classify THg in the tissues in accordance with the abiotic differences, by the salinity values, in the areas. All tests were evaluated for significance at α = 0.05 and the statistical calculations were executed by using GraphPad Prism and Past 3.0.
Results and Discussion
The northern region showed lower values of salinity, as a consequence of the freshwater inflow due to the proximity to the Ribeira de Iguape River input by Valo Grande channel. Meanwhile, the southern area showed a more hydrodynamic profile, with higher salinity due to the influence of the marine water and tidal cycle (Table 1). The biological data and THg contents in different tissues are shown in Table 1. HSI showed no significant differences between the sexes in the two areas (northern and southern) of the CIELC. CF values did not show significant sex differences, however, there were higher THg values in fish from the northern area. The lowest HSI values were observed in male Ariidae catfish from the northern regions of the CIELC compared to the southern region, likely. In general, results of the somatic indices suggest absence of hepatic stress in catfish from both areas, but a better health condition for catfish from the southern area than catfish from the northern region of the CIELC.
Independent of catfish sex, a decreasing pattern of accumulation was observed in this order: kidney > gonad > muscle > gill (Fig. 1). Taking into account the data from this study along with the results of THg contents in C. spixii collected in the southern area of the CIELC in 2009 (Azevedo et al. 2012b), it was possible observe a decrease in the THg contents in all tissues of catfish sampled in 2014. However, individual catfish collected in 2014 were substantially smaller than specimens collected in 2009 and thus differences may be attributed to length and age differences. Concerning the THg concentrations by sex of the catfish, absence of sexual differences were observed in catfish sampled in 2009 (Azevedo et al. 2012b) and in the present work (male and female from the southern region of the CIELC). However, differences were observed regarding adults male catfish and juvenile catfish from the northern region of the CIELC (Fig. 2). Comparing the areas, in general, the median THg levels in the tissues of the catfish showed higher concentrations in fish from the southern area, where they are metabolically more active with more energy expenditure and greater detoxification capacity, thus with a higher level of metal in detoxification structures such as liver and kidney; due to the hydrodynamic conditions and not specifically due to the greater anthropogenic impact (northern). There was sexual variation between male and females and higher accumulation in the gonad of males than females (Fig. 2). Juvenile/immature catfish showed a similar profile of detoxification with the males since the THg levels in kidneys were similar. Absence of immature catfish in the southern region is a weak aspect that should be inspected in the future. The highest THg concentrations were found in the kidneys with a decrease toward: gonad, muscle and gill; and is corroborated by previous studies with other fish species (Kojadinovic et al. 2007; Kasper et al. 2009). However, it is a relatively recent approach with respect to estuarine fish on the Brazilian coast. The difference in the Hg accumulation associated to the maturity of the fish reflects a differential metabolism regarding THg bioaccumulation with respect to males, females and juveniles.

Box and whiskers plot of concentrations of THg in different tissues from female, male and juvenile specimens of catfishes captured in the southern and northern regions of the CIELC. Box plots (Bars = 10 and 90 percentiles; boxes = 25 and 75 percentile; vertical line = median). Outliers are indicated as * above the boxplots. Different letters indicate statistical differences Kruskal–Wallis test at p < 0.05
Figure 3a shows the PCA for THg levels in gills, muscles, kidneys and gonads of adult male and female and juveniles catfis from the northern and southern regions of the CIELC. PC1 explained 84% of the variance and 13% was explained by PC2. Two groups were clearly separated in PC2, representing the Southern (THg in kidneys of adults male and female) and Northern area (THg in kidneys of juveniles) in the CIELC without direct dependence of the abiotic parameter as salinity. In order to observe the relationship between the salinity and the THg contents in different tissues of adults (male and female) and juvenile catfish, cluster analysis was applied (Fig. 3b). The salinity had associated only the THg contents in the gills of adults and juvenile catfish. The gills, feeding, water uptake and the epithelial tissue are the most common input route of the xenobiotic in the fish (Heath 1990), where the gills represent the specific target of adsorption and absorption of metals (Kalbassi et al. 2011). Fluctuations in salinity represent variations in the salt balance which can change the pH and consequently the solubility of elements like trace metals in the water and in the medium as a whole (Kennish 1991). This changes the pattern of bioaccumulation and biodistribution of metals such as THg in the fish. Therefore, the grouping observed among THg levels in gills of catfish and the salinity of the water is representative and abiotic conditions should be evaluated in association with metabolic/biological conditions, especially in environments with large natural or anthropogenic transition gradients.

Multivariate approach by a PCA of THg levels in different tissues such as the gill, kidney, muscle and gonad from adult (female – F and male – M) and juvenile (Juv) catfish captured in the southern and northern regions of the CIELC and; b dendogram illustrating similarity (Euclidean distance) as assessed by UPGMA among the THg concentrations in the gill, kidney, gonad and muscle of adults (male and female) and juvenile catfish from the Southern and Northern regions of the CIELC
These observations reinforce the importance of considering the stage of maturity of individual fish in order to understand the bioaccumulation and metabolism of fish under different environmental stress and conditions. THg concentrations in the catfish gonads may reflect a direct effect on reproduction but the integrated observation of the THg levels with the gonad integrity by microscopy analysis is interesting.
Although the THg concentrations are below the limit established for human consumption of 1.0 µg g−1 (ANVISA 2013; USEPA 1997), the observed variations among the tissues reflect a differential bioaccumulation process, as well as the possibility of anthropogenic input in the northern area close to the Ribeira de Iguape River. It can be transported in association with the organic matter of the water and deposited towards the southern area of the CELCI. In fact, even at low concentrations, THg levels in this region deserve attention, since parts of the CELCI are considered a biosphere reserve (UNESCO) and therefore, over time, chronic effects and damages may be observed in the local biota, with a future ecosystem imbalance.
References
Adams SM, McLean RB (1985) Estimation of largemouth bass, Micropterus salmoides lacépède, growth using the liver somatic index and physiological variables. J Fish Biol 26:111–126
Agência Nacional de Vigilância Sanitária (ANVISA) (2013) Ministério da Saúde, Brasil. Resolução nº 42, de 29 de agosto de 2013. Dispõe sobre o Regulamento Técnico MERCOSUL sobre Limites Máximos de Contaminantes Inorgânicos em Alimentos. Available http://bvsms.saude.gov.br/bvs/saudelegis/anvisa/2013/rdc0042_29_08_2013.html. Accessed August 2018
Amorim EP, Fávaro DIT, Berbel GBB, Braga ES (2008) Assessment of metal and trace element concentrations in the Cananéia estuary, Brazil, by neutron activation and atomic absorption techniques. J Radioanal Nucl Chem 278(2):485–489
Azevedo JS, Sarkis JES, Hortellani MA, Ladle RJ (2012a) Are Catfish (Ariidae) effective bioindicators for Pb, Cd, Hg, Cu and Zn? Water Air Soil Pollut 223:3911–3922
Azevedo JS, Sarkis JES, Oliveira TA, Ulrich JC (2012b) Tissue-specific mercury concentrations in two catfish species from the Brazilian coast. Braz J Oceanogr 60(2):211–219
Dias ACL, Guimarães JRD, Malm O, Costa PAS (2008) Mercúrio total em músculo de cação Prionace glauca (Linnaeus, 1758) e de espadarte Xiphias gladius Linnaeus, 1758, na costa sul- sudoeste do Brasil e suas implicações para a saúde pública. Cad Saude Publica 24(9):2063–2070
Figueiredo JL, Menezes NA (1978) Manual de peixes marinhos do sudeste do Brasil. II. Teleostei (1). Museu de Zoologia da Universidade de São Paulo, São Paulo, 110
Gomiero LM, Braga FMS (2003) Relação peso-comprimento e fator de condição para Cichla cf. ocellaris e Cichlamonoculus (Perciformes, Cichlidae) no reservatório de Volta Grande, rio Grande-MG/SP. Acta Sci 25(1):79–86
Heath AG (1990) Water pollution and fish physiology. 2nd edn. CRC Press, London p 245
Kalbassi MR, Salari-Joo H, Johari A (2011) Toxicity of silver nanoparticles in aquatic ecosystems: salinity as the main cause in reducing toxicity. Iran J Toxicol 5(12):436–443
Kasper D, Palermo EFA, Dias ACMI, Ferreira GL, Leitão RP, Branco CWC, Malm O (2009) Mercury distribution in different tissues and trophic levels of fish from a tropical reservoir, Brazil. Neotrop Ichthyol 7(4):751–758
Kennish MJ (1991) Ecology of estuaries: anthropogenic effects. CRC Press, London
Kojadinovic J, Potier M, Le Corre M, Cosson RP, Bustamante P (2007) Bioaccumulation of trace elements in pelagic fish from the Western Indian Ocean. Environ Pollut 146:548–566
Lacerda LD, Malm O (2008) Contaminação por mercúrio em ecossistemas aquáticos: uma análise das áreas críticas. Estud Av 22(63):173–190
USEPA (1997) Guidance for assessing chemical contaminant data for use in fish advisories. Volume 2Risk Assessment and Fish Consumption LimitsSecond Edition. U.S. Environmental Protection Agency. Office of Water, Washington, DC
Vazzoler AEA De (1996) Biologia da reprodução de peixes teleósteos: Teoria e Prática. Maringá. EDUEM. 169
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical Approval
All applicable international, national, and/or institutional guidelines for the care and use of animals were followed
Research Involving Animal and Human Participants
This article does not contain any studies with human participants performed by any of the authors.
Additional information
The original version of this article was revised: the units in Materials and methods section was corrected.
Rights and permissions
About this article

Cite this article
Pecoraro, G.D., Hortellani, M.A., Hagiwara, Y.S. et al. Bioaccumulation of Total Mercury (THg) in Catfish (Siluriformes, Ariidae) with Different Sexual Maturity from Cananéia-Iguape Estuary, SP, Brazil. Bull Environ Contam Toxicol 102, 175–179 (2019). https://doi.org/10.1007/s00128-018-2485-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00128-018-2485-3