
Abstract
Key message
In tomato, genotype by watering interaction resulted from genotype re-ranking more than scale changes. Interactive QTLs according to watering regime were detected. Differentially expressed genes were identified in some intervals.
Abstract
As a result of climate change, drought will increasingly limit crop production in the future. Studying genotype by watering regime interactions is necessary to improve plant adaptation to low water availability. In cultivated tomato (Solanum lycopersicum L.), extensively grown in dry areas, well-mastered water deficits can stimulate metabolite production, increasing plant defenses and concentration of compounds involved in fruit quality, at the same time. However, few tomato Quantitative Trait Loci (QTLs) and genes involved in response to drought are identified or only in wild species. In this study, we phenotyped a population of 119 recombinant inbred lines derived from a cross between a cherry tomato and a large fruit tomato, grown in greenhouse under two watering regimes, in two locations. A large genetic variability was measured for 19 plant and fruit traits, under the two watering treatments. Highly significant genotype by watering regime interactions were detected and resulted from re-ranking more than scale changes. The population was genotyped for 679 SNP markers to develop a genetic map. In total, 56 QTLs were identified among which 11 were interactive between watering regimes. These later mainly exhibited antagonist effects according to watering treatment. Variation in gene expression in leaves of parental accessions revealed 2259 differentially expressed genes, among which candidate genes presenting sequence polymorphisms were identified under two main interactive QTLs. Our results provide knowledge about the genetic control of genotype by watering regime interactions in cultivated tomato and the possible use of deficit irrigation to improve tomato quality.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Today, agriculture is one of the primary water users in many regions of the world, but global warming and drought risks are threatening plant growth and productivity. In particular, the Mediterranean region should experience more frequent drought episodes in the next decades (Gao and Giorgi 2008; Dai 2011). In this area, economic losses due to water limitation could be critical for the fruit and vegetable productions (Katerji et al. 2008). Thus, a better management of water resource for crop production is needed. A commonly accepted solution is to improve plant adaptation to low water availability.
Many studies have assessed plant response to different watering regimes in several species and shown the negative impact of water shortage on plant growth and yield. Reviews of the different morphological, physiological and molecular changes induced by water limitation are available (Chaves et al. 2003; Hirayama and Shinozaki 2010; Blum 2011; Farooq et al. 2012; Silva et al. 2013). In particular, these studies highlight the role of secondary metabolites and carbohydrates in plant protection against photo-oxidative stress induced by stomata closure and cell dehydration (Gershenzon 1984; Chaves et al. 2009; Shaar-Moshe et al. 2015). These drought induced secondary metabolites are also essential compounds for quality of plant food products. For instance, ascorbic acid (Vitamin C), an important antioxidant for human diet, is well known for its role in scavenging reactive oxygen species (ROS) in plant under water stress (Jiang and Zhang 2002; Stevens et al. 2008). Evidence of the crucial role of sugars in osmotic adjustment induced by drought has been obtained in several species such as tomato (Bertin et al. 2000), Arabidopsis thaliana (Anderson and Kohorn 2001) or white lupine (Chaves et al. 2002). Well-mastered water deficit can thus help to achieve a tradeoff between crop yield and quality, reducing non-beneficial water consumption in crop production at the same time. Such deficit irrigation strategies are particularly under consideration in fleshy fruits for which consumers are expecting healthier and tastier products (Chaves and Oliveira 2004; Nora et al. 2012; Ripoll et al. 2014). However, knowledge about the Quantitative Trait Loci (QTLs) and genes involved in plant response to water deficit and their interactions is still lacking (Shinozaki and Yamaguchi-Shinozaki 2007; Ashraf 2010; Tardieu et al. 2011). Despite the identification of hundreds of genes involved in response to drought by gene expression analysis associated or not to linkage mapping, their roles and modes of action are still poorly understood (Lovell et al. 2015; Shaar-Moshe et al. 2015). Besides, these genes were mainly identified in A. thaliana under laboratory conditions (Seki et al. 2002) or in cereals (Langridge 2006; Barnabas et al. 2007). Not all of them are involved in adaptation process (Chaves et al. 2003).
Understanding the genetic determinism of genotype by watering regime interactions will constitute a basis for crop improvement, allowing the identification of favorable alleles under drought conditions (Collins et al. 2008; Tardieu and Tuberosa 2010). The emergence of high-throughput genomic tools and the availability of genome sequences for many crops facilitate the decomposition of genotype by environment interactions into underlying QTLs and/or genes (Des Marais et al. 2013; El-Soda et al. 2014b). These approaches will provide a better understanding of the ability of an individual genotype to adapt its phenotype in response to environmental constraints, a phenomenon termed as ‘phenotypic plasticity’ (Via and Lande 1985; Schlichting 1986). In the context of multiple environments, two main approaches are applied to map QTL by environment (QTL × E) interactions. The first one, looking at the effects of a given QTL in each environment, identifies different interactive QTL types (Malosetti et al. 2007; Yang et al. 2008; Van Eeuwijk et al. 2010; Korte et al. 2012; Li et al. 2015). In most cases, QTLs have a strong effect in one environment, but lower effect in another (differential sensitivity effect). More rarely and mainly in wild species, QTLs can show opposite effects for a same trait in different environments (antagonist effect). The second strategy consists in constructing composite variables measuring phenotypic plasticity to deal with univariate QTL mapping models. These variables can be ratio or difference between the values of a trait measured in two environments or parameters from reaction norms (Tétard-Jones et al. 2011; El-Soda et al. 2014a; Coupel-Ledru et al. 2014). The two methods substantially overlap but the second one gives additional statistical power with more QTLs exceeding the threshold (Tétard-Jones et al. 2011; El-Soda et al. 2014a).
In cultivated tomato (Solanum lycopersicum L.), a water demanding crop extensively grown in Mediterranean region, QTLs for chemical and physical fruit quality were previously mapped, but no attention was paid to the interaction with abiotic factors (Causse et al. 2001; Saliba-Colombani et al. 2001; Pascual et al. 2015). In this species, genes involved in response to abiotic stress were mainly characterized by translational genetics and genetic engineering with genes identified in A. thaliana (Hsieh et al. 2002; Rai et al. 2013; Zhu et al. 2014). Studies of natural variability of the interactions with environmental constraints have focused on salt stress (Foolad et al. 2003; Foolad 2004; Uozumi et al. 2012; Kissoudis et al. 2015; Asins et al. 2015). In few accessions, authors have reported a positive effect of mild to moderate water deficit on tomato fruit quality, with an increased fruit soluble solids levels and an increased concentration of hexoses (Mitchell et al. 1991; Bertin et al. 2000; Patanè and Cosentino 2010; Zheng et al. 2013). Besides, Foolad et al. (2003) and Semel et al. (2007) have shown some genetic variability in response to water deficit at the seed and plant levels. However, to date, no QTL by watering regime interaction mapping studies were conducted in the cultivated tomato. More precisely, introgression line populations involving wild relative species (Solanum habrochaites and Solanum pennellii) were used to map QTLs and the large confidence intervals obtained made the transposition difficult into the cultivated tomato (Gur et al. 2011; Easlon et al. 2014).
In this context, the aims of the present study are to: (1) describe genotype by watering regime interactions for plant and fruit traits in cultivated tomato genotypes, (2) decipher the inheritance patterns of these interactions and (3) identify candidate genes as putative targets for breeding. We addressed these aims by phenotyping a population of recombinant inbreed lines (RILs), grown in greenhouse under two watering regimes (drought and control), in two locations (Morocco and France). Linkage mapping was conducted to identify QTLs controlling genotype by watering regime interactions. Microarray analysis of gene expression in young leaves from the parental genotypes grown under the two watering regimes was performed to identify differentially expressed genes between the watering conditions. Finally, gene expression data were used to identify candidate genes underlying two interactive QTLs. The genetic determinism of genotype by watering regime interactions in cultivated tomato and the possible use of water deficit to improve tomato fruit quality in future breeding programs are discussed.
Materials and methods
Plant material and experimental design
The RIL population consisted in 119 F7 recombinant inbred lines. This population was developed from an intraspecific cross between two inbred lines, Cervil and Levovil (described in Saliba-Colombani et al. 2000). Cervil is a cherry type tomato (S. lycopersicum cerasiforme) with small fruits (6–10 g), whereas Levovil (S. lycopersicum) is a large fruited accession (90–160 g). In 2013, the plants, including the 119 RILs and the two parents, were grown in a heated glasshouse in INRA Avignon (Avi, France) from Mars to July. Besides, from December 2013 to May 2014, plants were grown in an unheated plastic greenhouse in the experimental site of the company GAUTIER Semences in Agadir (Aga, Morocco). In the greenhouses, the mean air temperature was 23 and 26 °C during day, 16° and 18 °C during night, in France and Morocco, respectively. In each experiment, plants were grown in 4 litters (l) plastic pots filled with peat (Klasmann 165) and watered with nutritive solution (2, 4, 6 mmol l−1, N, P, and K, respectively).
In both locations, two watering regimes were applied to the plants in each trial: drought (D) and control (C). Control treatment was applied according to ETp climatic data and the cultural coefficient for tomato crop under greenhouse with a maximal drainage of 25 % and a relative humidity of the peat substrate of 65 %. The drought treatment was progressively applied after flowering of the second truss of Cervil (considered as a reference early genotype): water supply was reduced by 25 % compared to control for 1 week, then decreased by 60 % until the end of the experiment, aiming to exacerbate the contrast with the control irrigation. Throughout the experiment, relative humidity of the peat substrate was controlled with a GRODAN® moisture probe and monitored in drought pots between 25 and 30 %. Genotypes were randomized within rows and watering regime was applied by row. For each experiment, two plants per watering regime per genotype were placed side by side. To insure relatively homogenous environment in the greenhouses, trials were surrounded with one row of border tomato plants.
Plant and fruit phenotyping
In the two trials, under the two watering regimes, RIL plants were phenotyped for traits describing plant performance and fruit characteristics. Vegetative vigor and phenology were measured daily on every plant. Flowering date of the first flower from the 5th truss in Avignon (Flw.Avi) and 4th in Agadir (Flw.Aga) were assessed in number of days after sowing. The implantation height (Ht.Avi and Ht.Aga, in cm), stem diameter (Diam.Avi and Diam.Aga, in mm) and leaf length (Leaf.Avi and Leaf.Aga, in cm) under truss were measured on the 4th truss in Avignon and the 5th truss in Agadir. The number of fruits per plant (Nbfruits.Avi) was assessed in Avignon only by counting all the fruits from the second truss to the sixth truss.
Fruit measurements were conducted on tomatoes harvested daily on the basis of their red color to ensure a homogeneous ripening stage. At least, ten fruits per genotype per watering regime were harvested in the two trials on 3rd to 6th truss. For each fruit, fresh weight (FW.Avi and FW.Aga, in g) and firmness (FIR.Avi and FIR.Aga, in Durofel index) were measured. Besides, in Avignon only, harvested fruits were pooled in three groups of three to four fruits per watering regime. These pools constituted the three replicates for chemical analysis. In each pool, a quarter of fruit pericarp was sampled and dried in an oven at 60 °C for 4 days to measure dry matter content (DMW.Avi, in %). Then, half of each fruit pool was mixed in juice to measure pH (pH.Avi) and soluble solid content (SSC.Avi, with a refractometer, in °Brix). Pericarps were sampled from the remaining fruit of each pool, frozen with liquid nitrogen and ground into fine powder with an IKA® mill for total Vitamin C (VitCFM.Avi, in mg per 100 g of fresh matter) assessment according to Stevens et al. (2006). Average total Vitamin C per genotype per watering regime was also expressed in mg per 100 g of dry matter (VitCDM.Avi) using DMW.Avi.
The average yield per genotype in Avignon (Yield.Avi, in g fresh weight per plant from truss 2–6) was estimated in each watering regime as the product of the average fruit fresh weight (FW.Avi) by the average number of fruits (Nbfruits.Avi). Finally, a total of 19 traits were assessed, under two watering conditions each, considering as two separate traits a same phenotypic measurement carried out in the two locations. The phenotypic means in the RIL population are available in Supplemental Table 1.
Statistical analyses on phenotypic data
Statistical analyses were performed on RIL raw data of each trial separately (Avignon and Agadir) using R 3.2.0 (R Development Core Team 2012). Prior to any analysis of variance (ANOVA), data were corrected for normality deviation using Box and Cox transformations (Box and Cox 1964). Effect of watering regime and interaction with genotype were tested by the ANOVA model: Y ij = μ + G i + W j + G i * W j + e ij , where Y ij was the phenotypic value of genotype i in watering regime j, μ the overall mean, Gi the fixed effect of genotype i, W j the fixed effect of watering regime j, and e ij the residual error effect. Residuals were spatially plotted to control for a potential microenvironment effect due to side by side position of the two replicates of a given genotype in each watering regime in the experiments. No significant pattern was identified and we chose to not include a spatial effect in the ANOVA model. To further describe the genotype by watering regime interaction, the G × W sum of squares was partitioned into part associated with heterogeneous variance (scale change) and part due to imperfect correlation between genotypes (rank change) using the method 1 of Muir et al. (1992). For FW, Nbfruits, VitCFM and VitCDM, ecovalences were calculated according to Wricke (1964) to measure participation of independent genotypes in interaction.
Then, genetic variability expressed at a given watering regime was assessed using the following ANOVA model: Y ij = µ + G i + e ij. (G i and e ij as random). Restricted maximum likelihood estimates (REML) of variances of the random factors (\(\sigma_{G}^{2}\) and \(\sigma_{e}^{2}\)) were computed. Broad sense heritability was calculated in each watering regime as: H 2 = \(\sigma_{G}^{2}\) / \(\sigma_{\text{Total}}^{2}\), with \(\sigma_{\text{Total}}^{2}\) = \(\sigma_{G}^{2}\) + \(\sigma_{e}^{2}\). For the different traits in the two trials, correlations between H 2 and \(\sigma_{G}^{2}\) measured under drought and under control conditions were estimated by Spearman coefficient and declared significant when P value were below 0.05.
For subsequent analyses, for each watering regime and each trial, the average genotypic values over replicates were computed. For each phenotypic trait k in each trial, plasticity (∆k) was calculated on the mean of the trait under each watering regime (drought D k ; control C k ) as: ∆k = (D k − C k )/C k . In the different watering regimes and in the different trials, Pearson correlations between means of traits and between means and plasticity data were calculated. A Mantel test was performed to measure changes in correlation between traits, according to the watering regime, in the two trials. P value was calculated after 9999 permutations and an alpha threshold of 0.05 was considered to declare significance.
Plant genotyping and genetic map building
Genotyping and map construction are described in Pascual et al. (2015). Briefly, a set of 754 polymorphic markers between the two parents were genotyped in the RIL population: 679 are SNP markers derived from the re-sequencing of the parent genomes (see Causse et al. 2013), two are RAPD markers (random amplified polymorphic DNA) and 73 are RFLP markers (restriction fragment length polymorphism) present in a previous genetic map from this progeny (Saliba-Colombani et al. 2000). The Chi-square test (α = 0.0001 %) revealed that 98 % of the markers (739/754) did not show any segregation distortions and were used in genetic mapping as described in Pascual et al. (2015). When several markers colocalized, only the one with the lowest percentage of missing data was conserved. The final genetic map obtained included 501 loci (501/754) and was covering 1090 cM corresponding to 98 % of the assembled tomato genome (Tomato Genome Consortium 2012). Markers were named according to their positions on the tomato genome (assembly v2.5), as Y01_56000045 at position 56,000,045 pb on chromosome 1. The genotypic data of the RIL population are available in the Supplemental Table 2. The genetic map is available in Pascual et al. (2015).
QTL and QTL × watering regime mapping
In each watering regime and each trial, the plasticity data and average phenotypic values were used for QTL detection. When distributions were skewed, corrections for normality were applied: Log10(Ht.Avi); Log10(Nbfruits.Avi); Log10(FW.Avi); √(Diam.Aga); Log10(Leaf.Aga); Log10(Ht.Aga); Log10(FW.Aga) and Log10(Nbfruits.Avi). The QTL detection was performed by simple interval mapping (Lander and Botstein 1989) using the EM algorithm method implemented in R/QTL package (Broman et al. 2003). A 1000-permutation test was performed to estimate significant threshold. LOD threshold was 3.08, corresponding to a genome wide significance level of α = 0.05. For each detected QTL, position, LOD score, marker at the LOD score peak, confidence interval (genetic CI, LOD decrease of one unit), average phenotypic values of the two parental alleles and percentage of phenotypic variation explained (PVE) were displayed. QTL effects were calculated as: (Cervil mean allele − Levovil mean allele)/2. The genetic CI was translated into physical intervals (Physical-CI in Mbp) onto the tomato genome (assembly v2.5). When a QTL was detected in one watering regime, the effect and PVE were also calculated in the second watering regime. Then, to test for watering regime and interaction with marker, two different ANOVA tests were developed in R 3.2.0 (R Development Core Team 2012):
-
1.
A “watering regime effect test” (W test) that compares a model with marker genotype and watering regime effect, to a model without watering regime effect;
-
2.
An “interaction effect test” (G × W test) that compares the full model, including the effect of the marker genotype and its interaction with the watering regime, to the one that doesn’t include interaction.
This testing method is inspired from the multi-trait mixed model (MTMM) developed by Korte et al. (2012) for association analysis, considering only fixed effects. To correct for multiple testing, significance thresholds P value corresponding to a genome wide significance level of α = 0.05 were computed by a 1000-permutation test (P value Wtest = 2.21 × 10−4; P valueG × Wtest = 1.93 × 10−4). This procedure allowed displaying P value for watering regime and interaction effect for marker at the QTL LOD score peak and to identify interactive markers not identified in the QTL mapping step.
Microarray experiment on parental accessions
Microarray analyses were performed on an Agilent four-plex arrays at IPS2 Transcriptomic Platform (INRA, France). For each of the 34,727 tomato genes (assembly v2.4, Tomato Genome Consortium 2012), a set of ten 60-mer probes were designed using the eArray Agilent software. Considering melting temperature and specificity criteria, the best probe for each gene was chosen and synthesized in forward and reverse sense. The array contained 33,913 forward and 33,913 reverse probes representing 98 % of the known tomato genes, each printed in technical duplicate and 18 controls in triplicate. The array design is available through the GEO at NCBI (GPL20224) and on the CATdb database (Gagnot et al. 2008): array ‘4PLEX_TOMATO’.
Samples of young leaves of Cervil and Levovil, grown under the two watering regimes in Avignon, were harvested, immediately frozen and ground in liquid nitrogen with an IKA® mill. RNA was extracted using the RNeasy Plant Mini Kit (Qiagen) following the manufacturer’s protocol, with DNase I treatment. RNA quality was assessed on Agilent 2100 Bioanalyser using Nano 6000 kit. Two independent biological replicates per genotype were produced. The labeling of cRNAs with Cy3-dUTP or Cy5-dUTP was randomly performed as described in Two-Color Microarray-Based Gene Expression Analysis Low Input Quick Amp Labeling manual (© Agilent Technologies, Inc.). Hybridization and washing were performed according to Agilent Microarray Hybridization Chamber User Guide instructions (© Agilent Technologies, Inc.). A two microns resolution scanning was performed using InnoScan900 scanner (InnopsysR, Carbonne, France) and raw data were extracted using the MapixR software (InnopsysR, Carbonne, France). In total, eight hybridizations were carried out comparing Cervil Control vs Cervil Drought and Levovil Control vs Levovil Drought. For each comparison, one technical replicate with dye swap was performed for each biological replicate (i.e., four hybridizations per comparison).
Microarray statistical analyses
Analysis was conducted with the R 3.2.0 software (R Development Core Team 2012). For each array, the raw data comprised the logarithm of the median feature pixel intensity at wavelengths 635 nm (red) and 532 nm (green). A global intensity-dependent normalization using the loess procedure (Yang et al. 2002) was performed to correct the dye bias. The differential analysis was based on the log10 of the fold changes between watering regimes averaging over the duplicate probes and over the technical replicates. Hence, the numbers of available data for each gene equals the number of biological replicates. Empirical Bayes posterior means were computed to smooth the specific variances and used to calculate the moderated t test (function SqueezeVar of the library limma, Smyth 2005). Under the null hypothesis, no evidence that the specific variances changed between probes was highlighted and consequently the moderated t-statistic was assumed to follow a standard normal distribution. To control the false discovery rate, P values were adjusted with the Bonferroni approach (Storey 2007) using the R library kerfdr (Guedj et al. 2009). We considered as being differentially expressed the probes with a Bonferroni-adjusted P values below 0.05. A Venn diagram was drawn to indicate genes differentially expressed in Cervil and/or in Levovil. Gene Ontology (GO) terms were associated with the differentially expressed genes using genome annotation v2.4 (Tomato Genome Consortium 2012). A maximum of seven GO terms were associated to each gene. A total of 33 % of the differentially expressed genes (DEG) were not associated to any GO term due to a lack in the genome annotation. Identification of GO terms related to biological process that were significantly enriched within the differentially expressed genes compared to the tomato genome was achieved using the ‘GO term enrichment analysis’ tool (http://bioinfo.bti.cornell.edu/tool/GO/GO_enrich.html) based on the ‘GO::TermFinder’ program described in Boyle et al. (2004). GO terms were declared significantly enriched when Bonferroni-corrected P value was below 0.05. Separate analyses for up and down regulated genes in Cervil only, in Levovil only and common to the two accessions were conducted. All raw and normalized data are available through the GEO at NCBI (GSE69898) and through the CATdb database (Gagnot et al. 2008): project ‘4PLEX_TOMATO_2013_03′. The list of genes differentially expressed and the GO data are available in the Supplemental Table 3.
Candidate gene selection under interactive QTLs
Microarray data on young leaves were used to identify candidate genes under interactive QTLs for plant traits potentially controlled at leaf level. We focused on two QTLs with short physical intervals: a QTL for flowering time on chromosome 2 (3.23 Mbp) and a QTL for stem diameter on chromosome 4 (2.55 Mbp). Within the QTL confidence intervals, differentially expressed genes between watering regimes in Cervil and/or in Levovil were selected (adjusted P values below 0.05). Among these genes, polymorphism data obtained through the re-sequencing of parental accessions (Causse et al. 2013) was screened to identify nucleotide variants between Cervil and Levovil (SNPs and Indels). Re-sequencing depth was 19.6× for Cervil (covering 88.8 % of the genome with a minimum depth of 4×) and 9.2× for Levovil (72.7 % of the genome with a minimal depth of 4×). Variants were classified in four categories as specified in Causse et al. (2013): ‘High’ for polymorphisms which modified splice sites or start/stop codons (loss or gain); ‘Moderate’ for non-synonymous polymorphisms in coding regions, ‘Low’ for variants in coding regions which do not change the amino acid sequence and ‘Modifier’ for polymorphisms located in upstream and downstream regions or in UTR or intergenic regions.
Results
To study the genetic variability of tomato plant and fruit response to water deficit, we mapped QTLs in a population derived from a cross between a cherry tomato and a large fruit accession, grown under two watering regimes (control and drought), in two locations (Agadir and Avignon). Phenotypic data from the two locations were analyzed separately because fewer and/or different phenotypic traits were measured in Morocco. Thus, a total of 19 traits were assessed in each watering condition, considering as two separate traits a same phenotypic measurement carried out in the two locations. However, for the six common traits, correlations between the two experiments under the two watering regimes were highly significant (P value <0.001), suggesting a good repeatability of the measurements through the experiments (Supplemental Table 4). Significant watering regime by location interactions were detected for these six traits (data not shown), which may reflect the consequences of differences in temperature and day length between locations.
Phenotypic variability and genotype by watering regime interactions
Both parental accessions were impacted by the drought treatment. From 1.1 to 33.1 times significantly higher percentages of phenotypic changes due to drought were observed in Levovil than in Cervil for 15 of the 19 measured traits (excepted for Diam.Aga, Leaf.Aga, FW.Aga and pH.Avi) (Fig. 1 and Supplemental Fig. 1). It suggested a higher susceptibility to water deficit in the large fruit accession. In particular, under drought, SSC was increased by 114.6 % and FW.Avi decreased by 71.8 % in Levovil, whereas the SSC gain was only 11.7 % and the FW loss 33.3 % in Cervil.

Distribution of the average plant and fruit traits in the recombinant inbred lines (RILs) grown under two watering regimes in Avignon. Opaque color indicates trait values under control treatment and transparent color trait values under drought treatment (green plant traits, red fruit traits). The parental mean values are indicated: full red line for Cervil in control treatment, dashed red line for Cervil in drought treatment, full black line for Levovil in control treatment and dashed black line for Levovil in drought treatment. The black arrows represent the RIL population means: dashed arrow for drought and full arrow for control treatment (color figure online)
In the RILs, we surveyed ample phenotypic variation for the plant and fruit traits, under the two watering regimes, in the two locations (coefficient of variation ranking from 2.48 to 46.27 %; average CV = 17.68 %) (Fig. 1 and Supplemental Fig. 1). Transgressions beyond parental values in the two directions were observed for all traits in the two watering regimes, except for Flw.Aga, FW.Aga, FW.Avi and Nbfruits.Avi for which RIL phenotypic means were comprised between Cervil and Levovil means. At plant level, in average in the RILs and in the two locations, the drought treatment tended to reduce stem diameter (Avi: −20.7 %; Aga: −30.3 %), leaf length (Avi: −13.4 %; Aga: −25.8 %) and fruit number (Avi: −21.7 %). At fruit level, drought treatment reduced FW (Avi: −37.7 %; Aga: −25.4 %) and yield (Avi: −50.3 %), but increased SSC (Avi: +26.3 %) and DMW (Avi: +30.7 %). The average plant height, flowering time, fruit firmness and pH were poorly affected by watering deficit whatever the location (Ht.Avi = −5.6 %; Ht.Aga = 2.4 %; Flw.Avi = −0.2 %; Flw.Aga = +0.6 %; FIR.Avi = +3.4 %; FIR.Aga = +0.8 %; pH.Avi = −3.2 %). Vitamin C was differently impacted by drought depending on the unit in which it was expressed: in average increased when expressed relatively to fresh matter (+26.3 %) and reduced when expressed relatively to dry matter (−8.9 %).
For all the traits measured with replicates, in the two locations, genotype by watering regime interaction was significant (P value <0.01), except for pH, which was also poorly variable in the population (CVdrought = 2.80 %; CVcontrol = 3.34 %) (Table 1). These interactions represented between 5.70 (FW.Avi) and 16.86 % (Diam.Avi) of the total sum of square, a proportion lower than the one due to the genotype (between 23.96 and 85.87 %) for all traits and lower than the one due to the watering regime (between 14.66 and 49.62 %) for ten of the seventeen traits. Interaction partitioning according to method 1 from Muir et al. (1992) showed that the observed interactions were mainly due to genotype re-ranking across watering regimes (84.94–100 %) and poorly to scale changes (0–15.6 %, Table 1).
The broad sense heritabilities were comprised between 0.30 (FIR.Avi under drought) and 0.90 (DMW.Avi under drought), with consistence between experiments for the six common traits in the two watering regimes (P valuedrought = 0.03 and r drought = 0.85; P valuecontrol = 0.03 and r control = 0.86) (Table 1, Supplemental Fig. 2). Correlations between heritability and genetic variance were significant in both conditions (for H 2: P value <2.2 × 10−16 and r = 0.92; for varG: P value = 8.56 × 10−11 and r = 0.97). Thus, genetic variability was conserved across watering treatments, interactions being associated with re-ranking among genotypes more than heterogeneous variance between watering regimes (Table 1).
Changes in correlations according to fruit weight and watering regime
We observed a negative linear relationships between FW in control condition and FW plasticity (∆FW) in the two experiments (rAvi = −0.51 and P valueAvi = 2.08 × 10−09; r Aga = −0.44 and P valueAga = 9.69 × 10−07) (Fig. 2a and Supplemental Fig. 3). As ∆FW measures the percentage of FW gain or loss due to the drought treatment, these negative relationships indicated higher fresh weight loss for lines with large fruits. On another side, correlation between FW in control condition and vitamin C plasticity when expressed relatively to fresh matter (VitCFM) was not significant (P value = 0.29 and r = 0.09), whereas this correlation was significantly negative (P value = 1.05 × 10−03 and r = −0.35) when vitamin C plasticity was expressed relatively to dry matter (VitCDM) (Fig. 2c, d). Together, these results suggest important water losses in large fruits under drought, responsible for a FW decrease and concentration of vitamin C in fruit, without clear increase of vitamin C synthesis or accumulation in fruit.

Linear relationships between FW in control condition and plasticity for (a) fresh weight, (b) fruit number, (c) vitamin C content relatively to fresh weight and (d) vitamin C content relatively to dry weight in Avignon. Equation and R 2 of the linear regression lines are displayed. Color indicates relative ecovalence classes: blue <0.1, 0.1 < cyan < 0.5; 0 .5 < black < 1; green >1. The gray areas indicate small fruit accessions (FW < 25 g) (color figure online)
Among small fruit accessions (FW below 25 g), ∆FW was comprised between −0.6 and 0.1 in Avignon and between −0.6 and 0.5 in Agadir (Fig. 2a and Supplemental Fig. 3). Plasticity for fruit number was comprised between −0.6 and 0.4 (Fig. 2b). Thus, part of the small fruit accessions had positive delta value for FW.Avi (1), FW.Aga (8) and Nbfruits.Avi (12), indicating improved yield components under drought. Besides, in Agadir, two accessions showed, at the same time, a positive delta value for FW and fruit number (SSD168 and SSD172) (Supplemental Table 1). Ecovalence measurement is a method developed by Wricke (1964) to partition the sum of square of the interaction term and measure participation to individual genotype or environment to the genotype by environment interaction. In the small fruit accessions, the median values of the ecovalence distributions were 0.30, 0.18 and 0.25 for FW.Avi, FW.Aga and Nbfruits.Avi, respectively (blue and cyan colors in the gray area on Fig. 2a, b; Supplemental Fig. 3). These low values observed for 50 % of the small fruit accessions indicated a relatively stable FW and fruit number for these genotypes, whatever the watering regime.
For all phenotypic traits, correlations between mean values under control and drought were highly significant (P value <0.01), correlation coefficients ranking from 0.26 (Yield.Avi) to 0.88 (Flw.Aga) (Supplemental Table 5). Nonetheless, we exhibited a significant change in correlation network between phenotypic traits according to the watering treatment, in the two experiments (P valueAvi = 4.0 × 10−4 and P valueAga = 9.7 × 10−3) (Fig. 3, Supplemental Fig. 4). Part of the correlations observed under control treatment was reinforced under water deficit, as for the positive correlation between yield and fruit number (r control = 0.31 and r drought = 0.49) (Fig. 3). Others correlations were reduced under drought treatment. FW was slightly less positively correlated with yield under drought (0.28) than under control watering (0.47) (Fig. 3). Together, these changes in correlation between FW and yield and between fruit number and yield suggested that fruit number was a major yield component under drought. Only one situation of correlation reversal between watering regimes was observed, between flowering date and DMW (Fig. 3).

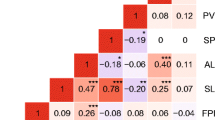
Changes in phenotypic correlation network between the two watering regimes. The figure displays Pearson correlation coefficients between average phenotypic values measured in control and drought treatment, in Avignon. Only coefficients higher than 0.2 are shown (P value <0.05). The line width is proportional to correlation coefficient value. The line color indicates direction of the correlation: green for positive correlations and red for negative correlations. ‘Flw’ for Flw.Avi; ‘Hgh’ for Ht.Avi; ‘Dmt’ for Diam.Avi; ‘Lef’ for Leaf.Avi; ‘Nbf’ for Nbfruits.Avi; ‘FW’ for FW.Avi; ‘FIR’ for FIR.Avi; ‘pH’ for pH.Avi; ‘DMW’ for DMW.Avi; ‘SSC’ for SSC.Avi; ‘VCF’ for VitCFM.Avi; ‘VCD’ for VitCDM.Avi and ‘Yld’ for Yield.Avi (color figure online)
QTLs and QTL by watering regime interactions
A total of 56 QTLs were mapped and 44 of them colocalized within five clusters on chromosomes 2, 3, 4, 6 and 11 (Table 2; Fig. 4). The 56 QTLs explained more than 5 % of the total phenotypic variance (PVE), with a median value of 14 % and a maximum of 41 % for FW.Avi in control treatment (Supplemental Table 6, Supplemental Table 7). Eight QTLs were detected both in Avignon and Agadir experiments (33 % of the QTLs detected on the six common traits between the two locations). The confidence intervals were smaller than 11 Mbp for 88 % of the QTLs. Seven QTLs mapped around the centromeres encompassing more than 30 Mbp. Besides, the size of the confidence intervals in cM and in Mbp were poorly correlated (P value = 0.01 and r = 0.38), due to differences in recombination rates along the genome. Pascual et al. (2015) and Sim et al. (2012) reported similar results in tomato and explained these findings by large genomic regions around the centromeres with roughly no recombination.

Overview of plant and fruit QTL identified on the tomato genome by QTL analysis in RILs. At the top of the panels, lines are representing tomato chromosomes where the lengths are proportional to chromosome physical sizes in million base pairs (Mbp). Centromeric regions with low recombination frequency are indicated in grey and peripheral parts in black (according to Sim et al. 2012). QTL are represented by square. Color codes correspond to the QTL types: constitutive (commons to control and drought treatment) in orange; detected only in control treatment in blue; detected only in drought treatment in red; interactive between the two watering regimes in purple. When an interactive QTL is colocalized with a non-interactive one, the interactive QTL is represented in the first plan in purple, surrounded by square with the color of the colocalized QTL. Positions of five major FW QTLs (: fw2.1, fw2.3, fw2.2, fw3.2, fw11.2 and fw11.3) are indicated (color figure online)
Twenty QTLs were detected only under the control conditions and 12 QTLs only under drought. Thirteen QTLs were constitutive as they were detected under both watering regimes. Distinguishing between specific and constitutive QTLs is not straightforward as it depends on the magnitude of the effect and the chosen detection threshold. Thus, we calculated the effects and PVE for all QTLs in the two watering regimes (Supplemental Table 6, Supplemental Table 7). On chromosomes 2 and 11, constitutive QTLs for FW colocalized with FW QTLs previously fine-mapped or cloned (Frary 2000; Lecomte et al. 2004; Huang and van der Knaap 2011; Illa-Berenguer et al. 2015; Fig. 4).
Eleven QTLs were significantly interactive between watering treatments, with two of them mapped both with the ANOVA testing procedure and with the plasticity data. The plasticity data gave more power to detect QTL by watering regime interaction, mapping ten interactive QTLs against three for the ANOVA testing procedure. One more interactive QTL, for yield on chromosome 8, was just below the threshold according to the ANOVA procedure (P value = 0.005) and was not detected with the plasticity data. Among interactive QTLs detected, four were associated with plant traits and seven to fruit quality traits. Seven antagonist QTLs had opposite allelic effects when comparing both watering treatments and four differential QTLs had effect intensity changed according to treatment (Table 2; Fig. 4). In average, the interactive QTLs explained 14 % of the phenotypic variance (SD = 5 %). One of them, controlling variation in FW, was in the centromeric region of chromosome 11 and covered 46 Mbp. The ten others encompassed in average 4 Mpb (SD = 3 Mbp) and genomic regions carrying between 221 and 1009 genes (Supplemental Table 8).
Among the interactive QTLs, the differential QTL for SSC on chromosome 2 mapped in the same genomic region as ssc2.2, a SSC QTL fine-mapped between two FW QTLs by Lecomte et al. (2004). The differential QTL for FW on chromosome 3 overlapped with fw3.2 which was recently cloned (Chakrabarti et al. 2013; Fig. 4). This QTL was shown to control the increase in cell layers, the delay of fruit ripening and the decrease in fruit number as well. Besides, this QTL was reported to have a minor effect on fruit shape and to be sensitive to the growing environment (Zhang et al. 2012). In this same region at the extreme end of chromosome 3, we mapped also three antagonist QTLs for SSC, DMW and VitCDM. An antagonist QTL for pH mapped at 38 Mbp on chromosome 6 could be related to detected associations for organic acid content in an unrelated tomato population. These QTLs/associations are close from two putative malate transporters (Solyc06g072910 and Solyc06g072920) identified in a previous study (Sauvage et al. 2014). The differential QTL for FW on chromosome 11 mapped 2 Mbp ahead of the fine-mapped QTLs fw11.2 and fw11.3 (Huang and van der Knaap 2011; Illa-Berenguer et al. 2015). No interaction with the environmental condition was demonstrated for these two QTLs until now.
Three examples of interactive QTLs are displayed in Fig. 5. On chromosome 3, an interactive QTL had antagonist effect on DMW.Avi: in control treatment Cervil allele increased the trait value of 0.4 units whereas under drought Cervil allele reduced DMW of 0.3 units (Fig. 5a). On chromosome 4, an interactive QTL with antagonist effect on stem diameter was mapped: in control treatment Cervil allele decreased stem diameter of 0.7 units whereas under drought Cervil allele increased Diam.Avi of 0.1 units (Fig. 5b). On chromosome 2, an interactive QTL was detected with changes in effect intensity according to watering regime for flowering time (Fig. 5c). Cervil allele effect was increased (+1.2 units) under drought (meaning an earlier flowering).

Examples of interactive QTL effects (a) ‘Antagonist’ interactive QTL on chromosome 3 for fruit DMW measured in Avignon (marker at the LOD peak: Y03_64701243). b ‘Antagonist’ interactive QTL on chromosome 4 for stem diameter measured in Avignon (marker at the LOD peak: Y04_63370382). c ‘Differential’ interactive QTL on chromosome 2 for flowering time measured in Avignon (marker at the LOD peak: Y02_38601550)
Genes differentially expressed and candidate gene identification under interactive QTLs
To go further in the understanding of the genetic control of tomato response to water deficit, gene expression was measured in young leaves of Cervil and Levovil, grown under the two watering regimes. Among the 33,913 tomato genes carried on the microarrays, 2259 were differentially expressed between watering treatments in young leaves of Cervil and/or Levovil (Fig. 6; Supplemental Table 3). More genes were differentially expressed in Levovil (1911), than in Cervil (786). Roughly identical counts of up (Cer: 43 % and Lev: 44 %) and down (Cer: 57 % and Lev: 57 %) regulated genes were observed in the two accessions. A total of 438 genes were differentially expressed in both parental accessions and 405 of them showed regulation in the same direction in Levovil and Cervil.

Venn diagram of differentially expressed genes between watering regimes in Cervil and/or Levovil. Genes were considered as being differentially expressed when the Bonferroni-adjusted P value was below 0.05. Blue and red colors indicate gene differentially expressed in Levovil and Cervil, respectively. Up and down arrows show genes up and down regulated under water deficit, respectively (color figure online)
The enrichment analysis of GO terms related to biological process was achieved on the differentially expressed genes. The 405 genes differentially expressed in the same direction in Cervil and Levovil contained more genes associated with microtubule process, to lipid metabolism and response to wound stress than the proportions observed in the whole tomato genome. Among the genes differentially expressed only in Cervil, processes associated with the defense against biotic stress and cell-wall process were significantly overrepresented compared to the tomato genome (Supplemental Fig. 5a). On the other hand, the list of genes differentially expressed only in Levovil was significantly enriched in genes related to cellular homeostasis, oxidation–reduction and metabolic process (Supplemental Fig. 5b). The differences between Levovil and Cervil in the enriched functions of the differentially expressed genes supported the differences observed between small and large fruit accessions at the phenotypic level. Finally, among the 33 genes regulated in different direction between Cervil and Levovil, three genes were associated with response to stress stimulus (Supplemental Fig. 5d): Solyc11g028060 (‘defensin-like protein’), Solyc06g009140 (‘Late embryogenesis abundant protein 3′) and Solyc07g006380 (‘defensin-like protein’). Interestingly, two of these genes were located in close vicinity of interactive QTLs (Fig. 4): Solyc06g009140 was located 2 Mbp above a differential QTL for fruit number on chromosome 6, whereas Solyc11g028060 was located in the interval of the differential QTL for FW on chromosome 11. They could be related to the phenotypic difference observed between Levovil and Cervil under drought and represent candidate genes for future studies, although these results should be taken cautiously as their expression was studied in leaves and related to fruit traits.
Then, we focused on two short genomic regions where interactive QTLs for stem diameter (chromosome 4, antagonist QTL, detected in Agadir and Avignon) (Fig. 5b) and for flowering time (chromosome 2, differential QTL, Agadir) (Fig. 5c) were mapped to look deeper at the differentially expressed genes (Supplemental Table 6, Supplemental Table 7). The interactive QTL for Flw.Aga corresponded to a genomic region carrying 357 genes, whereas the QTL for stem diameter encompassed 289 genes (Tomato Genome Consortium’ 2012) (Supplemental Table 8). Selecting the differentially expressed genes in Cervil and/or Levovil in these intervals reduced the candidate gene list to 24 and 29 genes, for Flw QTL and Diam QTL, respectively (Table 3).
The re-sequencing of Cervil and Levovil genomes identified polymorphisms between these accessions and constituted a powerful tool to further reduce the differentially expressed gene lists (Causse et al. 2013). Under the interactive QTL for stem diameter, 24 genes differentially expressed were polymorphic between Cervil and Levovil. Among them, seventeen genes exhibited moderate effect polymorphism and two genes had a polymorphism with a high impact on the protein sequence: Solyc04g077640 coding for a ‘serine carboxypeptidase 1’ (splice site donor) and Solyc04g079080 coding for a ‘calmodulin’ (frame shift) (Table 3). Among the 24 differentially expressed genes under the interactive QTL for Flw, eleven were polymorphic between Cervil and Levovil. Five genes presented moderate effect polymorphisms (non-synonymous variants in coding region) and only one had a polymorphism with a high impact on the protein sequence: Solyc02g069060 coding for a ‘phloem lectin’ (loss of a stop codon) (Table 3). These moderate to high effect polymorphisms and differentially expressed genes constitute putative candidates for the genetic control of tomato response to water deficit and have to be further investigated. Nevertheless, others polymorphic genes in these QTL intervals represent others putative candidates. Apart from the polymorphisms described above and in Table 3, we identified 17 genes with high effect variants under the Diam QTL (Solyc04g076410, Solyc04g076840, Solyc04g076940, Solyc04g077050, Solyc04g077330, Solyc04g077630, Solyc04g077700, Solyc04g077710, Solyc04g077920, Solyc04g078080, Solyc04g078180, Solyc04g078230, Solyc04g078260, Solyc04g078350, Solyc04g078360, Solyc04g078660 and Solyc04g078910) and 4 genes (Solyc02g065250, Solyc02g068970, Solyc02g069140 and Solyc02g069270) under the Flw QTL (polymorphism details in Causse et al. 2013).
Discussion
The aims of the study were (1) to outline genotype by watering regime interactions for 19 plant and fruit traits in 119 RILs from a cross between a small fruit accession and a large fruit accession, grown under two watering regimes, (2) to elucidate the inheritance patterns of these interactions and (3) to identify candidate genes as putative targets for tomato breeding under deficit irrigation. The results provided a basis for improving the use of deficit irrigation strategies for tomato production.
Genotype by watering regime interaction at the phenotypic level
At the phenotypic level, we identified significant genotype by watering regime interactions for most of the traits evaluated (except pH). The importance of the interaction with respect to watering and genotype factors may depend on the phenotypic traits and the genotypes studied, as well as the plant developmental stage and the intensity/duration of the water deficit suffered by the plants. In our study, the interactions, although marginal in regard to the magnitude of the effect of the genotype factor (24–86 %), represented up to 17 % of the total sum of squared deviations. They were essentially due to genotype re-ranking (85–100 %), and poorly to heterogeneous variance between watering conditions. In a previous publication on leaf water content in six cultivated tomato accessions grown under two watering regimes, the authors reported a significant genotype by watering treatment interaction representing 16 % of the total sum of square, against 5 and 72 % for the genotype and watering factor, respectively (Jureková et al. 2011). On the other hand, in thirty wheat lines phenotyped for traits measuring vigor of seeds and seedlings under two watering conditions, genotype by watering interactions represented between 8 and 39 % of the total sum of square and their weight was always equal or lower than the weight of the genotype factor (Dhanda et al. 2004).
Heritabilities and genetic variance were highly correlated between the two watering treatments, in the two locations. These results are contrasting with the only detailed study of genotype by watering regime interaction in tomato reported by Gur et al. (2011) in a set of introgression lines (ILs) derived from a cross between S. pennellii and S. lycopersicum. The authors described decreased genetic variances and heritabilities under drought. The discrepancy with our results could be caused by the different genetic basis of the populations, a lower number of tested lines and/or a more drastic drought treatment (no water supply) in Gur et al. (2011). In rice, intermediate results on 151 lines were reported, with a conserved heritability under arid conditions for some traits (grain and biomass yield), decreased (plant height, 1000-grain-weight) or increased (harvest index) for others, with differences according to water deficit intensity (Babu et al. 2003).
The pattern of genotype by environment interaction and the level of heritability under stress conditions are important features to consider when choosing a breeding strategy. When correlations are imperfect between environmental conditions (revealed through re-ranking of genotypes) and if there is genetic variability under stress, selecting one genotype for a specific environmental condition seems to be the best strategy. However, such breeding strategy is limited by the variability across years in the intensity and frequency of the drought episodes. Alternative approaches could be to improve drought adaptation in elite varieties by incorporating morphological and physiological mechanisms maintaining genotype performances under drought or to improve yield potential in already drought adapted accessions (Mitra 2001).
Interaction between genotype and the watering regime at the genotypic level
For the first time, in a cross between two cultivated tomato accessions, we identified QTL by watering regime interaction, deciphering the genetic architecture of tomato response to water deficit. Such an approach for dissecting G × E interaction into underlying genetic loci is not new and was already performed in numerous plant species. In the first QTL by environment studies, authors performed independent QTL mapping in each environment and compared the QTLs obtained through the experiments (Paterson et al. 1991 in tomato; Jansen et al. 1995 in A. thaliana; Lu et al. 1997 in rice). Today, more complex mapping strategies are undertaken and allow a more refined understanding of QTL × E interactions. Models can test for the presence of QTL whose effect vary between environments or plasticity QTLs can be mapped using composite traits measuring genotypic response to environmental constraints (Van Eeuwijk et al. 2010; Des Marais et al. 2013; Li et al. 2015).
Applying these complementary strategies, we identified 56 QTLs with moderate (10 %) to high (40 %) percentage of phenotypic variance explained. Among them, 13 were constitutive, 20 were control specific, 12 were drought-specific and 11 were interactive between watering regimes. Excepted the seven QTLs encompassing centromeric regions with low recombination frequency (Sim et al. 2012; Pascual et al. 2015), we mapped QTLs with relatively small confidence intervals covering 0.33 to 11 Mbp. The others reported QTL × watering regime studies in tomato used introgression lines and reported QTLs with large confidence intervals covering up to an entire arm of chromosome (Gur et al. 2011; Easlon et al. 2014). The plasticity data gave more power to detect QTL by watering regime interaction, mapping ten interactive QTLs against three for the ANOVA testing procedure (with two common between methods). Tétard-Jones et al. (2011) and El-Soda et al. (2014a) obtained such proportion when comparing both mapping methods and proposed that mapping QTL using directly phenotypic difference or ratio could give additional statistical power by exacerbating contrasts between two environmental conditions. Alternatively, when more than two environments are compared, parameters from reaction norms can be used as plasticity variable.
Although it is difficult to make a precise comparison because the authors did not exactly test QTL × E interaction, the relative proportion of different QTL types was relatively similar in the study of Gur et al. (2011) on tomato introgression lines. In this latter study on tomato yield and quality traits, a majority of the QTLs was constitutive (45 %) and control-specific (39 %) and few drought-specific QTLs (16 %) were mapped. Part of our constitutive QTLs confirmed the constitutive loci identified by these authors, in particular for the FW QTLs located on chromosomes 2 and 11 (Fig. 4). These QTLs were colocalized with cloned or fine-mapped genes controlling tomato FW: fw2.1 and fw2.3 (fine-mapped in Lecomte et al. 2004), fw2.2 (cloned in Frary 2000), fw11.2 and fw11.3 (fine-mapped in Huang and van der Knaap 2011; Illa-Berenguer et al. 2015). Their constitutive feature is promising in regard to tomato yield improvement for diverse environments. From these results, Gur et al. (2011) proposed that tomato yield under drought conditions would be mostly controlled by QTLs determining the productivity of the plant, rather than QTLs providing a physiological improvement for drought tolerance. However, we moderate this hypothesis arguing that distinguishing between specific and constitutive QTLs is not straightforward as it depends on the magnitude of the effect and the chosen detection threshold. We agree with Des Marais et al. (2013) who encourage authors to display effects and PVE for all QTLs in the different environmental conditions to make fair comparison between studies. Besides, quantification of QTL × E interactions appears necessary to be able to detect QTLs with varying effects depending on the environmental conditions and to identify QTLs/genes potentially involved in tolerance mechanisms against abiotic stress.
On average, the eleven interactive QTLs detected in our study explained 14 % of the phenotypic variance for the respective traits. Among them, a majority had antagonistic effects according to watering regime (seven QTLs) and four QTLs showed a decreased effect under drought. To date, only four studies have reported QTLs with changing direction of allelic effects according to the environmental constraints (Des Marais et al. 2012). Such antagonist QTLs were described in rice in response to planting density (Liu et al. 2012) and between different water regimes in A. thaliana (Hausmann et al. 2005; El-Soda et al. 2014a) and sorghum (Sabadin et al. 2012). In our study, the detection of loci with antagonist effect could result from a drastic drought treatment and testing different levels of watering in a next QTL study may give different results. Negative correlation between water deficit intensity and fruit fresh weight decrease was already observed in tomato (Durán Zuazo et al. 2011), but the effect of different water reduction intensity on the genetic determinants of tomato plant response to drought has been poorly investigated. In A. thaliana, the expression patterns of 6180 genes that were differentially expressed under severe drought was not significantly changed under moderate water deficit (Harb et al. 2010). Besides, 18 A. thaliana mutants that behaved better under severe water stress did not present any superiority under moderate stress (Skirycz et al. 2011). Such results are in line with a different genetic determinism depending on the level of water deficit suffered by the plant. Skirycz et al. (2011) proposed that severe drought may be favorable for detecting QTLs relative to limitation of water depletion in plant tissues (referred as avoidance strategy) whereas mild stress would allow to identify loci responsible for maintening growth, photosynthesis and metabolism during water deficit (tolerance strategy).
Knowledge of the interactive QTLs is crucial in breeding programs because the presence of such QTLs can limit breeding efficiency if the favorable alleles do not have the same effect under different environmental conditions. We identified antagonist QTLs for DMW, SSC, stem diameter, fruit number and pH on chromosomes 3, 4 and 6. Luckily, tomato and others horticultural crops are widely grown under irrigation and the level of water deficit imposed to the plants can be controlled and used to manage fruit quality, yield and water consumption. Furthermore, the antagonist QTLs identified could be used in marker-assisted selection (MAS) to build either genotypes for growth specifically under deficit irrigation or genotypes to cultivate under well-watered conditions. However, specific consideration should be given to the overlap of interactive QTLs at the terminal end of chromosome 3. In this region, we mapped one differential QTL for FW and three antagonist QTLs for DMW, SSC and VitCDM. Under control, Levovil alleles increased FW and VitCDM, whereas it decreased SSC and DMW. In drought conditions, the Levovil alleles increased FW in a lower extend, but decreased VitCDM and increased SSC and DMW. This cluster of QTL colocalized with the cloned QTL fw3.2 which was shown to be environmentally sensitive and to have pleiotropic effects on fruit cell numbers, fruit shape, ripening date and fruit number (Zhang et al. 2012; Chakrabarti et al. 2013). Besides, previous mapping studies in tomato grown under well watered condition identified QTLs for sugar content and titrable acidity in the same genomic region (Saliba-Colombani et al. 2001; Pascual et al. 2015). It is difficult to determine if the interactive QTLs correspond to a unique QTL with pleiotropic effect on the different traits whatever the watering regime or to different QTLs controlling the different traits under the two watering treatments. If there are several QTLs, recombination events could help to gather the most favorable alleles (or less unfavorable) to build a genotype suitable for growing under deficit irrigation.
Could we stand on water management to improve tomato fruit quality?
Deficit irrigation and partial root drying are watering strategies under consideration in fruit crops, aiming to reduce non-beneficial water consumption in horticultural production while maintaining the economic feasibility of the cropping systems (Kirda et al. 2004; Cui et al. 2008; Zheng et al. 2013). Major fruit species are highly water demanding crops often cultivated with abundant irrigation, which can lead to overuse of groundwater and environmental degradations. Beyond the concerns of gaining in water productivity and controlling yield losses, these practices may also contribute to improve fruit flavour and nutritional quality. In tomato, flavour perception is an important criteria for genetic improvement since consumers started complaining about lack of taste and aroma in the new long shelf life varieties (Kader et al. 1977; Bruhn et al. 1991; Ratanachinakorn et al. 1997). Tomato flavour results from complex interactions between sugars, organic acids and tens of volatile aromas (Stevens 1972; Yilmaz 2001). Besides, an abundant scientific literature have reported the favorable effects of tomato consumption on human health (Giovannucci 1999; Khachik et al. 2002; Giovannucci 2002), in particular through its content in ascorbic acid (vitamin C) and carotenoids which are among the most effective antioxidant in plants (Smirnoff 1996). Sugars, acids and antioxidants are also involved in plant response against stressing factors and their contents in fruits may be improved by the application of abiotic stress (in particular drought; Lester 2006; Dorais et al. 2008).
Due to the application of a 60 % water deficit, we observed reduced plant vigor (stem diameter, leaf length and plant height) and productivity (fruit number and FW). On average, in the RIL population, the yield decrease reached the substantial value of 50 %, hardly compatible with a sustainable production system. However, at the same time, tomato fruit-soluble solid and dry matter content were increased by 26 and 31 % on average, respectively. These results confirmed decreased yield and improved fruit quality previously reported in many fruit crop species cultivated under water limitation, among which peach tree (Mirás-Avalos et al. 2013), grape wine (Santesteban and Royo 2006) and tomato (Guichard et al. 2005; Zheng et al. 2013; Ripoll et al. 2014). In accordance with others studies (Zairi et al. 2003; Kirda et al. 2004), we support that a limited water deficit could be the best compromise between crop yield and fruit quality. Response of large genotypic sets to low and mild water stress should be studied to assess the optimal stress level.
Concerning vitamin C content in fruit, results were different depending if concentration were reported on the basis of fresh weight or dry weight. When expressed relatively to fresh weight, vitamin C content was increased by 26 % in average in the RIL population, whereas when expressed relatively to dry weight it was decreased by 9 %. In their review, Ripoll et al. (2014) pointed out such discrepancy for acid and sugar content in many fruits. Regarding how metabolite contents are measured, it is thus difficult to define if an increase in concentration results from a dilution/concentration effect, solute accumulation or synthesis in the fruit. Recent studies in tomato showed increased concentration of vitamin C with various extend depending on genotypes and water stress level, but the contents were expressed relatively to fruit fresh weight only (Favati et al. 2009; Murshed et al. 2013). More refined study of various metabolite contents in tomato fruits grown under drought should be achieved to clearly state on the effect of water limitation on the nutritional value of tomato.
The highlight of our study is that the response to water stress depends on fruit size leading to different water management strategies. Large fruits suffered from a greater loss of FW due to the water deficit, but they were also those whose dry matter and soluble solid contents most increased. Growing large fruit tomatoes under a slight water deficit could improve tomato flavour and limit water consumption. To compensate the yield loss for growers, the fruits of plants grown with a slight water deficit could be marketed as ‘tasty and environment friendly food’ (Dorais et al. 2008). On the other hand, in small fruit accessions, we observed a large genotypic variability for fresh weight and fruit number plasticity under drought. In particular, a dozen lines showed stable or increased FW and/or fruit number under drought. They represent good candidates for tomato yield improvement under drought. Genotypic variability in small fruit lines could be further explored to limit water consumption in cherry tomato greenhouses. In the long term, interesting ‘plasticity alleles’ identified in small fruit genotypes could be introgressed in large fruit plants provided they have no pleiotropic effect on fruit size.
Studying G × E interactions in the omic era
Genetic and genomic methods are now available to accelerate the identification of candidate genes and polymorphisms, through fine-mapping and functional genomic studies. It gives opportunity to further decipher the genetic basis of phenotypic plasticity. Here, we combined QTLs and genomic approaches to gain knowledge on the genetic architecture of tomato response to water deficit. The re-sequencing of the RIL parents identified thousands of SNPs to build a new genetic map covering fully the tomato genome (Causse et al. 2013; Pascual et al. 2015). The availability of the reference genome sequence and its functional annotation allowed the projection of the QTL confidence intervals onto the tomato physical map, identifying hundreds of genes located in these intervals (Tomato Genome Consortium 2012). Then, we focused on two interactive QTLs potentially controlled at leaf level and with short confidence intervals: an antagonist QTL for stem diameter on chromosome 4 and a differential QTL for flowering time on chromosome 2. Combining the results of QTL mapping to the analysis of gene expression of the RIL parents drastically reduced the list of candidate genes under these two interactive QTLs by targeting differentially expressed genes between watering regimes. The catalog of polymorphisms between the parental accessions gained from their re-sequencing reduced further the putative gene list. Under the two QTLs, three promising candidate genes with differential expression and high impact polymorphisms were identified: Solyc02g069060 was coding for a ‘phloem lectin’ (loss of a stop codon), Solyc04g077640 for a ‘serine carboxypeptidase 1′ (splice site donor) and Solyc04g079080 for a ‘calmodulin’ (frame shift) (Table 3). The involvement of genes of these three protein families in plant protection and/or signaling in response to biotic and abiotic stress was reported in many studies and strengthened the interest paid to these three loci. Lectins constitute a class of carbohydrate-binding proteins with a known role in plant protection against cold, drought, salinity and biotic stress. They seem to be involved in cellular regulation and signaling in many plants (Van Damme et al. 2004; Jiang et al. 2010). Calmodulin is involved in plant response to abiotic stress through the involvement in osmotic adjustment and stress signaling in interaction with cellular calcium (Gong et al. 1997; Perruc et al. 2004; Reddy et al. 2011). A serine carboxypetidase type 1 was identified for its role in response to wound stress in tomato (Moura et al. 2001) and a rice serine carboxypeptidase-like gene was shown to be involved in response to biotic and oxidative stress (Liu et al. 2008). However, numerous serine carboxypeptidases genes are present in tomato genome and more precise study of these specific genes should be done.
On the other hand, others polymorphic genes in the QTL intervals remain equally good candidates as differential expression is not always a requisite and differences in stability and activity of proteins may not be related to differences in mRNA production. Besides, polymorphisms in non-coding regions may also affect gene expression or protein stability. Furthermore, we identified 17 and 4 genes not differentially expressed but with high effect variants under the stem diameter and the flowering time QTL, respectively. In particular, one of the polymorphic genes under the interactive QTL for flowering time (Solyc02g069270 coding for ‘SlAGO2b’—frame shift) belonged to the argonaute (AGO) gene family known to be involved in RNA silencing pathways and interaction with microRNAs in plants (Vaucheret 2008). This gene is located in a region of chromosome 2 including two others AGO genes (SIAGO2a and SIAGO3), the first (Solyc02g069260 in Table 3) presenting moderate effect polymorphisms and being differentially expressed depending on the watering regime in our experiments. In a previous study on tomato, this set of genes was shown to be up-regulated under tomato yellow leaf curl virus (TYLCV) infection and under several abiotic stress, including drought stress (Bai et al. 2012). The argonaute genes may play an important role in reproductive development of tomato plant subjected to biotic and abiotic stressors by involving miRNA, in line with several results obtained in A. thaliana (Vaucheret 2004; Lee et al. 2010; Westwood et al. 2013).
A pioneer publication on A. thaliana provided another more advanced strategy to use gene expression data as clues for the identification of candidate genes involved in plant response to drought (Lovell et al. 2015). The authors proposed to use gene expression as covariate in a QTL model to link markers, RNA expression and phenotypes. They selected genes with significant cis-eQTLs and tested the effect of their transcript abundance on the effect of QTLs for phenotypic plasticity traits. Efficiency of the method was proved by recovering the causal locus FRIGIDA (previously cloned by Lovell et al. 2013) among 92 cis-regulated genes in the confidence interval of a QTL for water use efficiency (WUE). However, a limited number of studies have explored the genetic variation in transcriptome response to environmental constraints in large populations (Des Marais et al. 2013). The development of new sequencing technologies at a reduced cost may help to improve the quality of genome sequences, produce marker and gene expression datasets and allow the automatic functional annotation in many crops, which is an essential condition to implement strategies combining gene expression and QTL analysis in future research.
Genome wide association studies (GWAs) are another possible framework for the dissection of the genetic basis of phenotypic plasticity in diverse plant populations. GWA approaches benefit from the many recombination events experienced in natural populations to achieve QTL mapping leading to a few candidate genes or even to the identification of the causal polymorphisms. Besides, GWAs access to a larger genetic diversity, with more alleles than in a bi-parental cross. GWA models testing G × E interactions have been developed by plant biologists (Korte et al. 2012; Saïdou et al. 2014), but the combination of GWAs and gene expression was essentially reported in human genetics until now (Cheung et al. 2005; Cookson et al. 2009) and in few studies on A. thaliana (Nicolae et al. 2010; Chan et al. 2011). It is probably because GWA models may be less powerful than bi-parental population to identify QTLs when they have small effects, which is the case for most of the QTLs involved in phenotypic plasticity (Des Marais et al. 2012). This limitation can be bypassed using large populations designed to balance the allele frequencies. Combining QTL mapping, GWAs in large population and gene expression will constitute a complete framework to obtain a fine picture of the genetic control of genotype by watering regime interactions in plant. The advantage of a dual mapping strategy would be to reduce the rate of false positives and detect false negatives suffered by GWA due to structuration of the mapping panel, while taking advantage of the great allelic diversity in diverse populations (Brachi et al. 2010; Sterken et al. 2012).
Conclusion
This work is the first QTL study of response to water deficit in cultivated tomato (S. lycopersicum). At the phenotypic levels, significant genotype by watering regime interactions were reported. Large fruit tomatoes were more sensitive to drought and will require specific breeding considerations for growing under deficit irrigation, to achieve a trade-off between fruit quality improvement and yield. At the genotypic level, we identified interactive QTLs, many exhibiting effects changing direction depending on the watering regime. In the scope of plant breeding program, these QTLs could be used in marker-assisted selection (MAS) to develop tomato genotypes adapted to water limitation with intent to limit overuse of groundwater. In regard to genetic developments, we demonstrated a convenient way of combining QTL data, gene expression analysis and polymorphism data to identify candidate genes for plant adaptation to drought in the high-throughput area. Further studies need to confirm their roles. Besides, organ-specific transcriptome analysis will be of main interest to reveal the regulation network of tomato response to water deficit in a more refined scale.
Author contribution statement
EA conducted experiments in France, analyzed data and wrote the manuscript. JG and NB sampled and collected phenotypic data in France. JB sampled and collected phenotypic data in Morocco. SP, JPT and FB performed microarray experiments and gene expression quantification. MC supervised the project, built the experimental design and revised the manuscript. All authors discussed the results and commented the manuscript. Authors declared no conflict of interest in the authorship and publication of this document.
References
Anderson CM, Kohorn BD (2001) Inactivation of Arabidopsis SIP1 leads to reduced levels of sugars and drought tolerance. J Plant Physiol 158:1215–1219. doi:10.1078/S0176-1617(04)70149-2
Ashraf M (2010) Inducing drought tolerance in plants: recent advances. Biotechnol Adv 28:169–183. doi:10.1016/j.biotechadv.2009.11.005
Asins MJ, Raga V, Roca D et al (2015) Genetic dissection of tomato rootstock effects on scion traits under moderate salinity. Theor Appl Genet 128:667–679. doi:10.1007/s00122-015-2462-8
Babu RC, Nguyen BD, Chamarerk V et al (2003) Genetic analysis of drought resistance in rice by molecular markers. Crop Sci 43:1457–1469. doi:10.2135/cropsci2003.1457
Bai M, Yang G-S, Chen W-T et al (2012) Genome-wide identification of Dicer-like, Argonaute and RNA-dependent RNA polymerase gene families and their expression analyses in response to viral infection and abiotic stresses in Solanum lycopersicum. Gene 501:52–62. doi:10.1016/j.gene.2012.02.009
Barnabas B, Jager K, Feher A (2007) The effect of drought and heat stress on reproductive processes in cereals. Plant Cell Environ 31:11–38. doi:10.1111/j.1365-3040.2007.01727.x
Bertin N, Guichard S, Leonard C et al (2000) Seasonal evolution of the quality of fresh glasshouse tomatoes under Mediterranean conditions, as affected by air vapour pressure deficit and plant fruit load. Ann Bot 85:741–750. doi:10.1006/anbo.2000.1123
Blum A (2011) Plant water relations, plant stress and plant production. Plant breeding for water-limited environments. Springer, New York, pp 11–52
Box G, Cox D (1964) An analysis of transformations. J R Stat Soc Ser B 26:211–252
Boyle EI, Weng S, Gollub J et al (2004) GO: TermFinder—open source software for accessing gene ontology information and finding significantly enriched gene ontology terms associated with a list of genes. Bioinformatics 20:3710–3715. doi:10.1093/bioinformatics/bth456
Brachi B, Faure N, Horton M et al (2010) Linkage and association mapping of Arabidopsis thaliana flowering time in nature. PLoS Genet 6:e1000940. doi:10.1371/journal.pgen.1000940
Broman KW, Wu H, Sen S, Churchill GA (2003) R/qtl: QTL mapping in experimental crosses. Bioinformatics 19:889–890. doi:10.1093/bioinformatics/btg112
Bruhn CM, Feldman N, Garlitz C et al (1991) Consumer perceptions of quality: apricots, cantaloupes, peaches, pears, strawberries and tomatoes. J Food Qual 14:187–195. doi:10.1111/j.1745-4557.1991.tb00060.x
Causse M, Saliba-Colombani V, Lesschaeve I, Buret M (2001) Genetic analysis of organoleptic quality in fresh market tomato. 2. Mapping QTLs for sensory attributes. TAG Theor Appl Genet 102:273–283. doi:10.1007/s001220051644
Causse M, Desplat N, Pascual L et al (2013) Whole genome resequencing in tomato reveals variation associated with introgression and breeding events. BMC Genom 14:791. doi:10.1186/1471-2164-14-791
Chakrabarti M, Zhang N, Sauvage C et al (2013) A cytochrome P450 regulates a domestication trait in cultivated tomato. Proc Natl Acad Sci 110:17125–17130. doi:10.1073/pnas.1307313110
Chan EKF, Rowe HC, Corwin JA et al (2011) Combining genome-wide association mapping and transcriptional networks to identify novel genes controlling glucosinolates in Arabidopsis thaliana. PLoS Biol 9:e1001125. doi:10.1371/journal.pbio.1001125
Chaves MM, Oliveira MM (2004) Mechanisms underlying plant resilience to water deficits: prospects for water-saving agriculture. J Exp Bot 55:2365–2384. doi:10.1093/jxb/erh269
Chaves MM, Pereira JS, Maroco J et al (2002) How plants cope with water stress in the field? Photosynthesis and growth. Ann Bot 89:907–916. doi:10.1093/aob/mcf105
Chaves MM, Maroco JP, Pereira JS (2003) Understanding plant responses to drought—from genes to the whole plant. Funct Plant Biol 30:239. doi:10.1071/FP02076
Chaves MM, Flexas J, Pinheiro C (2009) Photosynthesis under drought and salt stress: regulation mechanisms from whole plant to cell. Ann Bot 103:551–560. doi:10.1093/aob/mcn125
Cheung VG, Spielman RS, Ewens KG et al (2005) Mapping determinants of human gene expression by regional and genome-wide association. Nature 437:1365–1369. doi:10.1038/nature04244
Collins NC, Tardieu F, Tuberosa R (2008) Quantitative trait loci and crop performance under abiotic stress: where do we stand? Plant Physiol 147:469–486. doi:10.1104/pp.108.118117
Cookson W, Liang L, Abecasis G et al (2009) Mapping complex disease traits with global gene expression. Nat Rev Genet 10:184–194. doi:10.1038/nrg2537
Coupel-Ledru A, Lebon E, Christophe A et al (2014) Genetic variation in a grapevine progeny (Vitis vinifera L. cvs Grenache × Syrah) reveals inconsistencies between maintenance of daytime leaf water potential and response of transpiration rate under drought. J Exp Bot 65:6205–6218. doi:10.1093/jxb/eru228
Cui N, Du T, Kang S et al (2008) Regulated deficit irrigation improved fruit quality and water use efficiency of pear-jujube trees. Agric Water Manag 95:489–497. doi:10.1016/j.agwat.2007.11.007
Dai A (2011) Drought under global warming: a review. Wiley Interdiscip Rev Clim Chang 2:45–65. doi:10.1002/wcc.81
Des Marais DL, McKay JK, Richards JH et al (2012) Physiological genomics of response to soil drying in diverse Arabidopsis accessions. Plant Cell 24:893–914. doi:10.1105/tpc.112.096180
Des Marais DL, Hernandez KM, Juenger TE (2013) Genotype-by-environment interaction and plasticity: exploring genomic responses of plants to the abiotic environment. Annu Rev Ecol Evol Syst 44:5–29. doi:10.1146/annurev-ecolsys-110512-135806
Dhanda SS, Sethi GS, Behl RK (2004) Indices of drought tolerance in wheat genotypes at early stages of plant growth. J Agron Crop Sci 190:6–12. doi:10.1111/j.1439-037X.2004.00592.x
Dorais M, Ehret DL, Papadopoulos AP (2008) Tomato (Solanum lycopersicum) health components: from the seed to the consumer. Phytochem Rev 7:231–250. doi:10.1007/s11101-007-9085-x
Durán Zuazo VH, Pleguezuelo CRR, Tarifa DF (2011) Impact of sustained-deficit irrigation on tree growth, mineral nutrition, fruit yield and quality of mango in Spain. Fruits 66:257–268. doi:10.1051/fruits/2011038
Easlon HM, St. Clair DA, Bloom AJ (2014) An introgression from wild tomato (Solanum habrochaites) affects tomato photosynthesis and water relations. Crop Sci 54:779–784. doi:10.2135/cropsci2013.06.0401
El-Soda M, Boer MP, Bagheri H et al (2014a) Genotype-environment interactions affecting preflowering physiological and morphological traits of Brassica rapa grown in two watering regimes. J Exp Bot 65:697–708. doi:10.1093/jxb/ert434
El-Soda M, Malosetti M, Zwaan BJ et al (2014b) Genotype × environment interaction QTL mapping in plants: lessons from Arabidopsis. Trends Plant Sci 19:390–398. doi:10.1016/j.tplants.2014.01.001
Farooq M, Hussain M, Wahid A, Siddique KHM (2012) Plant responses to drought stress. Springer, Berlin, Heidelberg
Favati F, Lovelli S, Galgano F et al (2009) Processing tomato quality as affected by irrigation scheduling. Sci Hortic (Amsterdam) 122:562–571. doi:10.1016/j.scienta.2009.06.026
Foolad MR (2004) Recent advances in genetics of salt tolerance in tomato. Plant Cell Tissue Organ Cult 76:101–119. doi:10.1023/B:TICU.0000007308.47608.88
Foolad MR, Zhang LP, Subbiah P (2003) Genetics of drought tolerance during seed germination in tomato: inheritance and QTL mapping. Genome 46:536–545. doi:10.1139/g03-035
Frary A (2000) fw2.2: a quantitative trait locus key to the evolution of tomato fruit size. Science 289(80):85–88. doi:10.1126/science.289.5476.85
Gagnot S, Tamby J-P, Martin-Magniette M-L et al (2008) CATdb: a public access to Arabidopsis transcriptome data from the URGV-CATMA platform. Nucleic Acids Res 36:D986–D990. doi:10.1093/nar/gkm757
Gao X, Giorgi F (2008) Increased aridity in the Mediterranean region under greenhouse gas forcing estimated from high resolution simulations with a regional climate model. Glob Planet Change 62:195–209. doi:10.1016/j.gloplacha.2008.02.002
Gershenzon J (1984) Changes in the levels of plant secondary metabolites under water and nutrient stress. In: Timmermann BN, Steelink C, Loewus FA (eds) Phytochemical adaptations to stress. Springer, Boston, pp 273–320
Giovannucci E (1999) Tomatoes, tomato-based products, lycopene and cancer: review of the epidemiologic literature. JNCI J Natl Cancer Inst 91:317–331. doi:10.1093/jnci/91.4.317
Giovannucci E (2002) A prospective study of tomato products, lycopene and prostate cancer risk. Cancer Spectr Knowl Environ 94:391–398. doi:10.1093/jnci/94.5.391
Gong M, Chen S-N, Song Y-Q, Li Z-G (1997) Effect of calcium and calmodulin on intrinsic heat tolerance in relation to antioxidant systems in maize seedlings. Aust J Plant Physiol 24:371. doi:10.1071/PP96118
Guedj M, Robin S, Celisse A, Nuel G (2009) Kerfdr: a semi-parametric kernel-based approach to local false discovery rate estimation. BMC Bioinformat 10:84. doi:10.1186/1471-2105-10-84
Guichard S, Gary C, Leonardi C, Bertin N (2005) Analysis of growth and water relations of tomato fruits in relation to air vapor pressure deficit and plant fruit load. J Plant Growth Regul 24:201–213. doi:10.1007/s00344-005-0040-z
Gur A, Semel Y, Osorio S et al (2011) Yield quantitative trait loci from wild tomato are predominately expressed by the shoot. Theor Appl Genet 122:405–420. doi:10.1007/s00122-010-1456-9
Harb A, Krishnan A, Ambavaram MMR, Pereira A (2010) Molecular and physiological analysis of drought stress in Arabidopsis reveals early responses leading to acclimation in plant growth. Plant Physiol 154:1254–1271. doi:10.1104/pp.110.161752
Hausmann NJ, Juenger TE, Sen S et al (2005) Quantitative trait loci affecting δ13 C and response to differential water availability in Arabidopsis thaliana. Evolution (N Y) 59:81–96. doi:10.1111/j.0014-3820.2005.tb00896.x
Hirayama T, Shinozaki K (2010) Research on plant abiotic stress responses in the post-genome era: past, present and future. Plant J 61:1041–1052. doi:10.1111/j.1365-313X.2010.04124.x
Hsieh T, Lee J, Charng Y, Chan M (2002) Tomato plants ectopically expressing Arabidopsis CBF1 show enhanced resistance to water deficit stress. Plant Physiol 130:618–626. doi:10.1104/pp.006783.KIN1
Huang Z, van der Knaap E (2011) Tomato fruit weight 11.3 maps close to fasciated on the bottom of chromosome 11. Theor Appl Genet 123:465–474. doi:10.1007/s00122-011-1599-3
Illa-Berenguer E, Van Houten J, Huang Z, van der Knaap E (2015) Rapid and reliable identification of tomato fruit weight and locule number loci by QTL-seq. Theor Appl Genet. doi:10.1007/s00122-015-2509-x
Jansen RC, Van Ooijen JW, Stam P et al (1995) Genotype-by-environment interaction in genetic mapping of multiple quantitative trait loci. Theor Appl Genet 91:33–37. doi:10.1007/BF00220855
Jiang M, Zhang J (2002) Water stress-induced abscisic acid accumulation triggers the increased generation of reactive oxygen species and up-regulates the activities of antioxidant enzymes in maize leaves. J Exp Bot 53:2401–2410. doi:10.1093/jxb/erf090
Jiang S-Y, Ma Z, Ramachandran S (2010) Evolutionary history and stress regulation of the lectin superfamily in higher plants. BMC Evol Biol 10:79. doi:10.1186/1471-2148-10-79
Jureková Z, Németh-molnár K, Paganová V (2011) Physiological responses of six tomato (Lycopersicon esculentum Mill.) cultivars to water stress. J Hortic For 3:294–300
Kader AA, Stevens MA, Albright-Holton M, et al (1977) Effect of fruit ripeness when picked on flavor and composition in fresh market tomatoes. J Am Soc Hortic Sci 102:724–731
Katerji N, Mastrorilli M, Rana G (2008) Water use efficiency of crops cultivated in the Mediterranean region: review and analysis. Eur J Agron 28:493–507. doi:10.1016/j.eja.2007.12.003
Khachik F, Carvalho L, Bernstein PS et al (2002) Chemistry, distribution, and metabolism of tomato carotenoids and their impact on human health. Exp Biol Med 227:845–851
Kirda C, Cetin M, Dasgan Y et al (2004) Yield response of greenhouse grown tomato to partial root drying and conventional deficit irrigation. Agric Water Manag 69:191–201. doi:10.1016/j.agwat.2004.04.008
Kissoudis C, Chowdhury R, van Heusden S et al (2015) Combined biotic and abiotic stress resistance in tomato. Euphytica 202:317–332. doi:10.1007/s10681-015-1363-x
Korte A, Vilhjálmsson BJ, Segura V et al (2012) A mixed-model approach for genome-wide association studies of correlated traits in structured populations. Nat Genet 44:1066–1071. doi:10.1038/ng.2376
Lander ES, Botstein D (1989) Mapping mendelian factors underlying quantitative traits using RFLP linkage maps. Genetics 121:185–199
Langridge P (2006) Functional genomics of abiotic stress tolerance in cereals. Brief Funct Genom Proteom 4:343–354. doi:10.1093/bfgp/eli005
Lecomte L, Saliba-Colombani V, Gautier A et al (2004) Fine mapping of QTLs of chromosome 2 affecting the fruit architecture and composition of tomato. Mol Breed 13:1–14. doi:10.1023/B:MOLB.0000012325.77844.0c
Lee H, Yoo SJ, Lee JH et al (2010) Genetic framework for flowering-time regulation by ambient temperature-responsive miRNAs in Arabidopsis. Nucleic Acids Res 38:3081–3093. doi:10.1093/nar/gkp1240
Lester GE (2006) Environmental regulation of human health nutrients (ascorbic acid, carotene and folic acid) in fruits and vegetables. HortScience 41:1
Li S, Wang J, Zhang L (2015) Inclusive composite interval mapping of QTL by environment interactions in biparental populations. PLoS One 10:e0132414. doi:10.1371/journal.pone.0132414
Liu H, Wang X, Zhang H et al (2008) A rice serine carboxypeptidase-like gene OsBISCPL1 is involved in regulation of defense responses against biotic and oxidative stress. Gene 420:57–65. doi:10.1016/j.gene.2008.05.006
Liu G, Zhu H, Zhang G et al (2012) Dynamic analysis of QTLs on tiller number in rice (Oryza sativa L.) with single segment substitution lines. Theor Appl Genet 125:143–153. doi:10.1007/s00122-012-1822-x
Lovell JT, Juenger TE, Michaels SD et al (2013) Pleiotropy of FRIGIDA enhances the potential for multivariate adaptation. Proc R Soc B Biol Sci 280:20131043. doi:10.1098/rspb.2013.1043
Lovell JT, Mullen JL, Lowry DB et al (2015) Exploiting differential gene expression and epistasis to discover candidate genes for drought-associated QTLs in Arabidopsis thaliana. Plant Cell 27:969–983. doi:10.1105/tpc.15.00122
Lu C, Shen L, He P et al (1997) Comparative mapping of QTLs for agronomic traits of rice across environments by using a doubled-haploid population. Theor Appl Genet 94:145–150. doi:10.1007/s001220050393
Malosetti M, Ribaut JM, Vargas M et al (2007) A multi-trait multi-environment QTL mixed model with an application to drought and nitrogen stress trials in maize (Zea mays L.). Euphytica 161:241–257. doi:10.1007/s10681-007-9594-0
Mirás-Avalos JM, Alcobendas R, Alarcón JJ et al (2013) Assessment of the water stress effects on peach fruit quality and size using a fruit tree model, QualiTree. Agric Water Manag 128:1–12. doi:10.1016/j.agwat.2013.06.008
Mitchell JP, Shennann C, Grattan SR, May DM (1991) Tomato fruit yields and quality under water deficit and salinity. J Am Soc Hortic 116:215–221
Mitra J (2001) Genetics and genetic improvement of drought resistance in crop plants. Curr Sci 80:758–763
Moura DS, Bergey DR, Ryan CA (2001) Characterization and localization of a wound-inducible type I serine-carboxypeptidase from leaves of tomato plants (Lycopersicon esculentum Mill.). Planta 212:222–230. doi:10.1007/s004250000380
Muir W, Nyquist WE, Xu S (1992) Alternative partitioning of the genotype-by-environment interaction. Theor Appl Genet 84:193–200. doi:10.1007/BF00224000
Murshed R, Lopez-Lauri F, Sallanon H (2013) Effect of water stress on antioxidant systems and oxidative parameters in fruits of tomato (Solanum lycopersicon L, cv. Micro-tom). Physiol Mol Biol Plants 19:363–378. doi:10.1007/s12298-013-0173-7
Nicolae DL, Gamazon E, Zhang W et al (2010) Trait-associated SNPs are more likely to be eQTLs: annotation to enhance discovery from GWAS. PLoS Genet 6:e1000888. doi:10.1371/journal.pgen.1000888
Nora L, Dalmazo GO, Nora F, Rombaldi CV (2012) Controlled water stress to improve fruit and vegetable postharvest quality. In: Ismal M, Mofizur R, Hiroshi H (eds) water stress. Tech Open Science, Rijeka, pp 59–72
Pascual L, Albert E, Sauvage C et al (2015) Dissecting quantitative trait variation in the resequencing era: complementarity of bi-parental, multi-parental and association panels. Plant Sci. doi:10.1016/j.plantsci.2015.06.017
Patanè C, Cosentino SL (2010) Effects of soil water deficit on yield and quality of processing tomato under a Mediterranean climate. Agric Water Manag 97:131–138. doi:10.1016/j.agwat.2009.08.021
Paterson AH, Damon S, Hewitt JD et al (1991) Mendelian factors underlyiing quantitative traits in tomato: comparison across species, generation and environments. Genetics 127:181–197
Perruc E, Charpenteau M, Ramirez BC et al (2004) A novel calmodulin-binding protein functions as a negative regulator of osmotic stress tolerance in Arabidopsis thaliana seedlings. Plant J 38:410–420. doi:10.1111/j.1365-313X.2004.02062.x
Rai AC, Singh M, Shah K (2013) Engineering drought tolerant tomato plants over-expressing BcZAT12 gene encoding a C2H2 zinc finger transcription factor. Phytochemistry 85:44–50. doi:10.1016/j.phytochem.2012.09.007
Ratanachinakorn B, Klieber A, Simons DH (1997) Effect of short-term controlled atmospheres and maturity on ripening and eating quality of tomatoes. Postharvest Biol Technol 11:149–154. doi:10.1016/S0925-5214(97)00021-5
R Development Core Team (2012) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria
Reddy ASN, Ali GS, Celesnik H, Day IS (2011) Coping with stresses: roles of calcium- and calcium/calmodulin-regulated gene expression. Plant Cell 23:2010–2032. doi:10.1105/tpc.111.084988
Ripoll J, Urban L, Staudt M et al (2014) Water shortage and quality of fleshy fruits, making the most of the unavoidable. J Exp Bot 65:4097–4117. doi:10.1093/jxb/eru197
Sabadin PK, Malosetti M, Boer MP et al (2012) Studying the genetic basis of drought tolerance in sorghum by managed stress trials and adjustments for phenological and plant height differences. Theor Appl Genet 124:1389–1402. doi:10.1007/s00122-012-1795-9
Saïdou A-A, Thuillet A-C, Couderc M et al (2014) Association studies including genotype by environment interactions: prospects and limits. BMC Genet 15:3. doi:10.1186/1471-2156-15-3
Saliba-Colombani V, Causse M, Gervais L, Philouze J (2000) Efficiency of RFLP, RAPD, and AFLP markers for the construction of an intraspecific map of the tomato genome. Genome 43:29–40. doi:10.1139/gen-43-1-29
Saliba-Colombani V, Causse M, Langlois D et al (2001) Genetic analysis of organoleptic quality in fresh market tomato. 1. Mapping QTLs for physical and chemical traits. TAG Theor Appl Genet 102:259–272. doi:10.1007/s001220051643
Santesteban LG, Royo JB (2006) Water status, leaf area and fruit load influence on berry weight and sugar accumulation of cv. “Tempranillo” under semiarid conditions. Sci Hortic (Amsterdam) 109:60–65. doi:10.1016/j.scienta.2006.03.003
Sauvage C, Segura V, Bauchet G et al (2014) Genome-Wide Association in tomato reveals 44 candidate loci for fruit metabolic traits. Plant Physiol 165:1120–1132. doi:10.1104/pp.114.241521
Schlichting CD (1986) The evolution of phenotypic plasticity in plants. Annu Rev Ecol Syst 17:667–693
Seki M, Narusaka M, Ishida J et al (2002) Monitoring the expression profiles of 7000 Arabidopsis genes under drought, cold and high-salinity stresses using a full-length cDNA microarray. Plant J 31:279–292. doi:10.1046/j.1365-313X.2002.01359.x
Semel Y, Schauer N, Roessner U et al (2007) Metabolite analysis for the comparison of irrigated and non-irrigated field grown tomato of varying genotype. Metabolomics 3:289–295. doi:10.1007/s11306-007-0055-5
Shaar-Moshe L, Hübner S, Peleg Z (2015) Identification of conserved drought-adaptive genes using a cross-species meta-analysis approach. BMC Plant Biol 15:111. doi:10.1186/s12870-015-0493-6
Shinozaki K, Yamaguchi-Shinozaki K (2007) Gene networks involved in drought stress response and tolerance. J Exp Bot 58:221–227. doi:10.1093/jxb/erl164
Silva D, Albuquerque D, de Azevedo (2013) Drought and its consequences to plants—from individual to ecosystem. In: Akinci S (ed) Responses of organisms to water stress. InTech, Turkey, p 17
Sim S-C, Durstewitz G, Plieske J et al (2012) Development of a large SNP genotyping array and generation of high-density genetic maps in tomato. PLoS One 7:e40563. doi:10.1371/journal.pone.0040563
Skirycz A, Vandenbroucke K, Clauw P et al (2011) Survival and growth of Arabidopsis plants given limited water are not equal. Nat Biotechnol 29:212–214. doi:10.1038/nbt.1800
Smirnoff N (1996) The function and metabolism of ascorbic acid in plant. Ann Bot 78:661–669
Smyth GK (2005) Limma: linear models for microarray data. Bioinformatics and computational biology solutions using R and bioconductor. Springer, New York, pp 397–420
Sterken R, Kiekens R, Boruc J et al (2012) Combined linkage and association mapping reveals CYCD5;1 as a quantitative trait gene for endoreduplication in Arabidopsis. Proc Natl Acad Sci 109:4678–4683. doi:10.1073/pnas.1120811109
Stevens MA (1972) Relationships between components contributing to quality variation among tomato lines. J Am Soc Hortic Sci 97:70–76
Stevens R, Buret M, Garchery C et al (2006) Technique for rapid, small-scale analysis of vitamin C levels in fruit and application to a tomato mutant collection. J Agric Food Chem 54:6159–6165. doi:10.1021/jf061241e
Stevens R, Page D, Gouble B et al (2008) Tomato fruit ascorbic acid content is linked with monodehydroascorbate reductase activity and tolerance to chilling stress. Plant Cell Environ 31:1086–1096. doi:10.1111/j.1365-3040.2008.01824.x
Storey JD (2007) The optimal discovery procedure: a new approach to simultaneous significance testing. J R Stat Soc Ser B Stat Methodol 69:347–368. doi:10.1111/j.1467-9868.2007.005592.x
Tardieu F, Tuberosa R (2010) Dissection and modelling of abiotic stress tolerance in plants. Curr Opin Plant Biol 13:206–212. doi:10.1016/j.pbi.2009.12.012
Tardieu F, Granier C, Muller B (2011) Water deficit and growth. Co-ordinating processes without an orchestrator? Curr Opin Plant Biol 14:283–289. doi:10.1016/j.pbi.2011.02.002
Tétard-Jones C, Kertesz MA, Preziosi RF (2011) Quantitative trait loci mapping of phenotypic plasticity and genotype-environment interactions in plant and insect performance. Philos Trans R Soc Lond B Biol Sci 366:1368–1379. doi:10.1098/rstb.2010.0356
Tomato Genome Consortium’ (2012) The tomato genome sequence provides insights into fleshy fruit evolution. Nature 485:635–641. doi:10.1038/nature11119
Uozumi A, Ikeda H, Hiraga M et al (2012) Tolerance to salt stress and blossom-end rot in an introgression line, IL8-3, of tomato. Sci Hortic (Amsterdam) 138:1–6. doi:10.1016/j.scienta.2012.01.036
Van Damme EJM, Barre A, Rougé P, Peumans WJ (2004) Cytoplasmic/nuclear plant lectins: a new story. Trends Plant Sci 9:484–489. doi:10.1016/j.tplants.2004.08.003
Van Eeuwijk FA, Bink MC, Chenu K, Chapman SC (2010) Detection and use of QTL for complex traits in multiple environments. Curr Opin Plant Biol 13:193–205. doi:10.1016/j.pbi.2010.01.001
Vaucheret H (2004) The action of ARGONAUTE1 in the miRNA pathway and its regulation by the miRNA pathway are crucial for plant development. Genes Dev 18:1187–1197. doi:10.1101/gad.1201404
Vaucheret H (2008) Plant ARGONAUTES. Trends Plant Sci 13:350–358. doi:10.1016/j.tplants.2008.04.007
Via S, Lande R (1985) Genotype-environment interaction and the evolution of phenotypic plasticity. Evolution (N Y) 39:505–522
Westwood JH, Mccann L, Naish M et al (2013) A viral RNA silencing suppressor interferes with abscisic acid-mediated signalling and induces drought tolerance in Arabidopsis thaliana. Mol Plant Pathol 14:158–170. doi:10.1111/j.1364-3703.2012.00840.x
Wricke G (1964) The calculation of ecovalence in summer wheat and oat. Z Pflanzenzuecht 52:127–138
Yang YH, Dudoit S, Lin DM et al (2002) Normalization for cDNA microarray data: a robust composite method addressing single and multiple slide systematic variation. Nucleic Acids Res 30:15e–15. doi:10.1093/nar/30.4.e15
Yang J, Hu C, Hu H et al (2008) QTLNetwork: mapping and visualizing genetic architecture of complex traits in experimental populations. Bioinformatics 24:721–723. doi:10.1093/bioinformatics/btm494
Yilmaz E (2001) The chemistry of fresh tomato flavor. Turk J Agric 25:149–155
Zairi A, El Amami H, Slatni A et al (2003) Coping with drought: deficit irrigation strategies for cereals and field horticultural crops in central Tunisia. In: Rossi G, Cancelliere A, Pereira LS, Oweis T, Shatanawi M, Zairi A (eds) Tools for drought mitigation in Mediterranean regions. Springer, Netherlands, pp 181–201
Zhang N, Brewer MT, van der Knaap E (2012) Fine mapping of fw3.2 controlling fruit weight in tomato. Theor Appl Genet 125:273–284. doi:10.1007/s00122-012-1832-8
Zheng J, Huang G, Jia D et al (2013) Responses of drip irrigated tomato (Solanum lycopersicum L.) yield, quality and water productivity to various soil matric potential thresholds in an arid region of Northwest China. Agric Water Manag 129:181–193. doi:10.1016/j.agwat.2013.08.001
Zhu M, Chen G, Zhang J et al (2014) The abiotic stress-responsive NAC-type transcription factor SlNAC4 regulates salt and drought tolerance and stress-related genes in tomato (Solanum lycopersicum). Plant Cell Rep 33:1851–1863. doi:10.1007/s00299-014-1662-z
Acknowledgments
We acknowledge the experimental teams of UR GAFL and Gautier SEMENCES for their help in experimentation. We thank especially Yolande Carretero, Alain Goujon, Esther Pelpoir, Renaud Duboscq, Claire Duffes and the employees of “Domaine Margau” (Agadir) for their help in phenotyping plants. Thanks to Christopher Sauvage for his proofreading. The CTPS project TOMSEC supported this work. EA was supported by an INRA PhD fellowship.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by H.-Q. Ling.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Albert, E., Gricourt, J., Bertin, N. et al. Genotype by watering regime interaction in cultivated tomato: lessons from linkage mapping and gene expression. Theor Appl Genet 129, 395–418 (2016). https://doi.org/10.1007/s00122-015-2635-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00122-015-2635-5