
Abstract
Normal maize germplasm could be used to improve the grain yield of popcorn inbreds. Our first objective was to locate genetic factors associated with trait variation and make first assessment on the efficiency of advanced backcross quantitative trait locus (AB-QTL) analysis for the identification and transfer of favorable QTL alleles for grain yield components from the dent corn inbred. A second objective was to compare the detection of QTL in the BC2F2 population with results using F2:3 lines of the same parents. Two hundred and twenty selected BC2F2 families developed from a cross between Dan232 and an elite popcorn inbred N04 were evaluated for six grain yield components under two environments, and genotyped by means of 170 SSR markers. Using composite interval mapping (CIM), a total of 19 significant QTL were detected. Eighteen QTL had favorable alleles contributed by the dent corn parent Dan232. Sixteen of these favorable QTL alleles were not in the same or near marker intervals with QTL for popping characteristics. Six QTL were also detected in the F2:3 population. Improved N04 could be developed from 210 and 208 families with higher grain weight per plant and/or 100-grain weight, respectively, and 35 families with the same or higher popping expansion volume than N04. In addition, near isogenic lines containing detected QTL (QTL-NILs) for grain weight per plant and/or 100-grain weight could be obtained from 12 families. Our study demonstrated that the AB-QTL method can be applied to identify and manipulate favorable QTL alleles from normal corn inbreds and combine QTL detection and popcorn breeding efficiently.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Grain yield is subject to complex physiological events and is determined by simultaneous action of different yield components. Popcorn germplasm is generally inferior to normal maize in yield and other agronomic traits. It is common for breeders to introduce normal maize germplasm into popcorn to improve grain yield and yield components, as well as plant, flowering, and resistance traits (Crumbaker et al. 1949; Dofing et al. 1991; Ziegler and Ashman 1994; Li and Lu 2000; Li et al. 2002). Popping characteristics of derived lines from crosses between popcorn and normal corn germplasm play a determinant role in marketability. Elite popping characteristics of the popcorn germplasm must be recovered through 1–2 backcrosses with popcorn germplasm as the recurrent parents (Dofing et al. 1991; Ziegler and Ashman 1994; Li et al. 2002). Negative correlations between popping characteristics and grain yield and most grain yield components have kept backcross efficiencies low, with regard to combining traits through phenotypic selection (Dofing et al. 1990, 1991; Li et al. 1999). Progeny or lines from popcorn × dent/flint corn crosses showing improved grain yield components and plant traits combined with elite popping characteristics have been obtained through 1–2 backcrosses and successive self and selection for important target traits in each population (Johnson and Eldredge 1953; Robbins and Ashman 1984; Ashman 1991; Li and Lu 2000; Li et al. 2005; Niu 2006).
Advanced backcross quantitative trait locus (AB-QTL) analysis was first introduced to simultaneously identify and transfer favorable QTL alleles from donor lines (wild or unadapted germplasm) into the genomes of elite cultivars (Tanksley and Nelson 1996). From the advanced backcross population where the favorable QTL alleles were identified, improved lines with the genome almost the same as the elite recurrent parent, and near isogenic lines (NILs) containing the QTL of interest could be developed rapidly (Bernacchi et al. 1998a; Xie et al. 2007) and put to use in breeding or production, and used to further genetic research. This strategy has been successfully applied in tomato (Tanksley et al. 1996; Bernacchi et al. 1998a, b; Fulton et al.1997, 2000), rice (Xiao et al. 1998; Moncada et al. 2001; Septiningsih et al. 2003a, b), normal maize (Ho et al. 2002), wheat (Huang et al. 2003), and Barley (Pillen et al. 2003). To date, the QTL strategy has not been applied in an effort to improve popcorn.
In popcorn, QTL for popping characteristics have been detected with BC1 (Lu et al. 2003) and F2:3 (Babu et al. 2006; Li et al. 2006a, b) early generations derived from dent/flint corn × popcorn crosses. However, simulation results showed that the efficiency of marker-assisted selection (MAS) declined in later generations partly because of recombination between markers and QTL (Gimelfarb and Lande 1994; Edwards and Page 1994). The result of comparative QTL mapping in F2:3 and F6:7 generations in normal corn by Austin and Lee (1996) further suggested some of the QTL detected during early generations of maximum linkage disequilibria may be due to multiple, linked genes that are separated via recombination. This may result in dissipation of effects and reduce the potential gain from selection. Using a population of 220 BC2F2 families derived from a cross between a dent corn inbred Dan232 and an elite popcorn inbred N04, QTL for popping expansion volume (PEV) have been detected (Niu 2006). In consideration of the main objective to incorporate normal corn germplasm into popcorn, six grain yield components were investigated in this BC2F2 population in the present study. Our study was designed to locate and characterize genetic factors associated with trait variation and assess the efficiency of AB-QTL analysis for the identification and transfer of favorable QTL alleles for grain yield components from the dent corn inbred. Second, we compared the detection of QTL in the BC2F2 population with results from a previous study using F2:3 lines of the same parents grown at the same location and evaluated in the same year with the same cultivation practice (Li et al. 2006a, b). Previous studies have evaluated the consistency in QTL detection in different populations, environments, and generations, but have not evaluated the same parents at early unselected (F2:3) and later selected (BC2F2) populations under the same environment.
Materials and methods
Population development
The dent corn inbred Dan232 was used as the female parent to make a cross with N04, a popcorn inbred. Dan232 was derived from Lu 9 kuan × Dan340, which are classified as members of the Ludahonggu heterotic group according to pedigree. N04 was derived from a Chinese popcorn variety BL03. The F1 plants were backcrossed to N04 as the recurrent parent to develop 235 BC1F1 plants. All BC1F1 plants were backcrossed a second time to N04 as the recurrent parent to develop BC2F1 seeds and were selfed simultaneously. The 72 best BC2F1 families were selected based on grain weight per plant (>N04), 100-grain weight (>N04), and PEV (≥N04) of selfed ears of BC1F1 plants, and grown in the field to produce BC2F2 seeds. Subsequently, 220 of 812 BC2F2 families were selected according to the same criterion used to select the best BC2F1. During the selection, the grain weight per plant, 100-grain weight and PEV for N04 were 25.6 ± 0.4 g, 8.6 ± 0.1 g, and 20.2 ± 0.2 ml/g, respectively, which was used as the threshold for BC2F1 and BC2F2 families. Grain yield and grain yield component and other traits were measured in these 220 families.
Field trials and trait evaluation
The 220 BC2F2 families, F1 and both parents were evaluated in a completely random design of one-row plots with two replications in two environments, spring and summer, at the Scientific Research and Education Center of Henan Agricultural University near Zhengzhou, Henan, China, in 2004. The rows were 4 m long with 0.67 m spacing between rows. Plots were planted by hand at a density of 60,000 plants per ha. Standard cultivation management practices were used at each environment.
Ten plants from the middle of each plot were chosen for evaluation of the six yield components: ear length (EL, cm), ear diameter (ED, cm), kernel number per row (RKN), row number per ear (ERN), grain weight per plant (GWP, g), and 100-grain weight (100GW, g). Trait measurements averaged over the two replications were used as the preliminary data in the analyses.
Phenotypic data analysis
The correlation coefficients among the six grain yield components and PEV were calculated using PROC CORR of SAS Software (SAS Institute Inc. 1989). Coefficients of genotypic correlations between two traits were estimated by dividing the genotypic covariance between them (Covg12) by the square root of the product of their genotypic variances (σ 2 g1 and σ 2 g1). Broad sense heritabilities for the BC2F2 families on an entry mean basis were calculated by dividing the genotypic variance by the phenotypic variance (Hallauer and Mirando 1981). Confidence intervals on heritability estimates were calculated according to Knapp et al. (1985).
SSR analysis and map construction
Twenty seeds per BC2F2 family, F1 and the two parents were cultivated in a climatic chamber, and the leaf tissues (<2 weeks old) were collected and bulked for each entry. All leaf samples were stored at −80°C, and DNA was extracted using a CTAB procedure (Saghai-Maroof et al. 1984). SSR analysis was conducted as reported in Senior and Heun (1993).
A total of 613 SSR primer pairs, chosen from Maize GDB (http://www.maizegdb.org) for their uniform distribution throughout all ten maize chromosomes, were initially screened for their polymorphism between the two parents. Ultimately, 193 markers that clearly showed co-dominant segregation, were used to genotype the 220 BC2F2 families for which phenotypic data were available. Twenty-three SSR markers that showed serious segregation distortion were excluded from the analysis. Finally, 170 SSR markers were used to construct the linkage map with Joinmap 3.0b (van Ooijen and Voorrips 2001), according to the genetic linkage map constructed using 259 F2 populations derived from the same cross. This linkage map covered ten maize chromosomes with a total length of 1,575.1 cM and an average interval of 9.3 cM.
QTL analysis
Composite interval mapping (CIM) was used to map QTL and estimate their effects for each trait (Zeng 1993, 1994). Model 6 of the Zmapqtl procedure in QTL Cartographer Version 2.5 (Wang et al. 2006) was employed, specifying the five markers identified by stepwise regression that explained most of the variation for a given trait as genetic background parameters and a window size of 10 cM on either side of the markers flanking the test site. To identify an accurate significance threshold for each trait, an empirical threshold was determined for CIM using 1,000 permutations (Churchill and Doerge 1994).
Results
Marker segregation and genome composition of BC2F2 families
In an unselected BC2 population, the expected genotypic ratio would be 75% homozygotes (N04/N04) to 25% heterozygotes (N04/Dan232), or an allele frequency of 87.5% N04 to 12.5% Dan232 alleles. The ratio in our study population was 68% N04/N04 to 30% N04/Dan232 and 83% N04 to 15% Dan232, respectively, which clearly showed skewness to N04/Dan232 genotype and Dan232 alleles. This was due to skewed allele frequencies at 82 of 170 (48%) of marker loci. Seventy-seven of these loci (94%) were skewed toward N04/Dan232, while five (6%) were skewed toward N04 (χ 2 > 6.63 and P < 0.01). The distorted markers were distributed along all ten of the chromosomes, with 9, 6, 12, 6, 5, 3, 12, 10, 6, and 5 on each chromosome, respectively. Skewing toward the heterozygotes N04/Dan232 could be explained by the superiority of heterosis in selection carried out in the BC1 and BC2 generations during population development.
We compared the results of this study with our previous study of the F2:3 population, which included the same parents and the same 613 SSR markers, and found that more markers showed distortion in the BC2F2 population. Only 16 markers showed distortion in the F2:3 population, with 7, 6, and 3 (44, 38, and 19%) toward N04, Dan232, and N04/Dan232, respectively. Among them, only five markers showed distortion in both studies. However, the direction of skewness was different for each marker, with all toward N04/Dan232 in this study, while four were toward Dan232 and one was toward N04 in the previous F2:3 population study.
Performance of BC2F2 families for grain yield components and comparison with the F2:3 population
All traits differed greatly between the two parents and among BC2F2 families, and showed a pattern of continuous distribution around the mean (Table 1 and Fig. 1). Compared with the F2:3 population, the performance of all traits was lower in the BC2F2 population, especially for GWP and 100GW. Heritability estimates for all traits were from low to high (Table 2). About 100GW, EL, ERN, and ED all showed high positive phenotypic and genotypic correlations with GWP, indicating they all played important roles in determining GWP. The directions of correlation between RKN and GWP for phenotype and genotype were negatively and positively opposed, perhaps due to low heritability. Besides the phenotypic and genotypic correlations between EL and RKN and between ERN and RKN, and the genotypic correlation between 100GW and ERN and RKN, significant positive correlations were detected among all other traits, suggesting the potential for simultaneous improvement. For the correlations of PEV with grain yield components, both significant negative phenotypic and genotypic correlations between PEV and 100KW, and significant negative phenotypic correlations between PEV and RKN and significant negative genotypic correlations between PEV and GWP were detected. Significant positive phenotypic or genotypic correlations between PEV and ERN, and ED were found. The main reason for the positive correlations might be the very low GWP for BC2F2 families, ranging from 19.2 to 50.5 g (average 32.3 g). Interestingly, similar results were reported in previous studies using popcorn lines (Li et al. 2005). In the F2:3 population, both significant negative phenotypic and genotypic correlations between 100GW and RKN, and significant negative phenotypic and/or genotypic correlations between PEV and GWP, 100GW, ED, RKN, and RKN were detected.

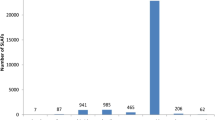
Frequency histograms for grain yield component traits of 220 BC2F2 families based on combined data across two environments
Compared with the popcorn inbred N04, all grain yield components were highly improved for BC2F2 families. For GWP, 100GW, and ED, 92–95% BC2F2 families had traits that were higher than N04; 82–89% for EL and RKN, and 38% for ERN.
QTL detected for each trait and comparison with the F2:3 population
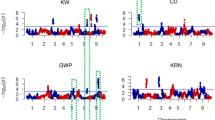
A total of 19 QTL significantly associated with six grain yield components were detected in BC2F2 families in this study. The QTL were located at chromosomes 1, 2, 3, 4, 5, 8, and 10, respectively, each with 1–5 per chromosome (Fig. 2 and Table 3).

SSR linkage map for Dan232 × N04 F2:3 and BC2F2 families. QTL one-LOD support intervals were indicated by vertical bars, and the maximum LOD peak positions were indicated by open/solid diamonds. QTL detected in the BC2F2 and F2:3 populations were indicated in solid and open vertical bars, respectively
GWP
Four QTL for GWP were detected and were located on chromosomes 1, 4, 8, and 10. The contributions to phenotypic variations for a single QTL varied between 5 and 19%, with qBEW-8-1 recording the highest contribution. The total contributions were 29%. All of the positive alleles were contributed by the dent corn parent Dan232 (the parent with much greater GWP in this and other environments). In the F2:3 population, only one QTL on chromosome 5 was detected. No identical QTL were detected between the two populations.
100GW
Three QTL for 100GW were detected on chromosomes 1, 5, and 8. The contributions to phenotypic variations for a single QTL varied between 7 and 15%, with qB100GW8-1 the highest. The total contributions were 32%. All of the positive alleles were contributed by the dent corn parent Dan232 (the parent with much greater 100GW in this and other environments). QTL on chromosomes 1 and 5 were also detected in the near chromosome interval and with similar contributions in the F2:3 population. However, the QTL on chromosome 8 of the BC2F2 was not detected in the F2:3 population, while a QTL on chromosome 7 was detected in the F2:3 population.
EL
Three QTL for EL were detected on chromosomes 3 and 4, with one QTL on chromosome 3 and two QTL on chromosome 4. The contributions to phenotypic variations for a single QTL varied between 7 and 11%, with qBEL4-1 contributing the highest percentage. The total contributions were 29%. The positive allele on chromosome 3 was contributed by the popcorn parent N04, while all of the positive alleles of the other two on chromosome 4 were contributed by the dent corn parent Dan232, although the EL of N04 was a little longer than Dan232. In the F2:3 population, three QTL on chromosomes 1, 3, and 7 were detected. The QTL on chromosome 3 was located in the near chromosome intervals umc1773 ∼ bnlg1035 and bnlg1452 ∼ umc1012, with similar contributions (7 and 6%) between the two populations.
ED
Six QTL for ED were detected on chromosomes 2, 4, 8, and 10, with one QTL on chromosomes 2 and 4, and two QTL on chromosomes 8 and 10. The contributions to phenotypic variations for a single QTL varied between 4% and a maximum of 16% for qBED8-1. The total contributions were 55%. All the positive alleles were contributed by the dent corn parent Dan232 (the parent with much greater ED in this and other environments). In the F2:3 population, four QTL on chromosomes 4, 5, 8, and 10 were detected. Although the QTL on chromosome 4 were in the same chromosome interval, phi079 ∼ bnlg1621, their contributions were 8 and 15% for the BC2F2 and F2:3 populations, respectively. qBED8-1 (phi115 ∼ bnlg1863) and qBED10-2 (umc1938 ∼ umc1053) were in the near chromosome interval with the QTL detected in the F2:3 population (bnlg2046 ∼ umc1562 and umc1677 ∼ umc2122), also varied, contributing 16 and 7% and 7 and 9%, respectively.
ERN
Two QTL for ERN were detected on chromosomes 5 and 10, with contributions to phenotypic variations of 8 and 9% for a total contribution of 15%. The positive alleles were all contributed by the dent corn parent Dan232. This was surprising because Dan232 has lower values for ERN. In the F2:3 population, four QTL on chromosomes 4, 7, and 10 were detected, with two QTL located on chromosome 4. Although QTL with similar contributions (9 and 9%) were detected on chromosome 10 in both populations, they were located on different chromosome intervals, umc2017 ∼ umc1877 and umc1677 ∼ umc2122 for F2:3 and BC2F2, respectively.
RKN
Only one QTL for RKN was detected on chromosome 8, with a contribution to phenotypic variation of 8%. Surprisingly, the positive allele was again contributed by the dent corn parent Dan232, even though Dan232 has lower values for RKN. Also, only one QTL were detected in the F2:3 population, and it was located on chromosome 7, with a contribution to phenotypic variation of 8%.
Discussion
Comparison of QTL detected in the BC2F2 and F2:3 populations
In all, nineteen and sixteen QTL were detected in the BC2F2 and F2:3 populations, respectively, for the same six grain yield components. Only six QTL (accounting for 32%) detected in the BC2F2 population were located in the same/near chromosome intervals as in the F2:3 population, and only three resulted in similar contributions. Thirteen QTL were specific to the BC2F2 population and ten QTL were only detected in the F2:3 population. This discrepancy may indicate a need for caution while carrying out MAS in backcross breeding using the mapping result with the F2:3 population.
In our study, both the F2:3 and BC2F2 populations were planted at the same location and in the same seasons in the same year and cultivated using the same methods. In addition, the same markers with polymorphism were used to analyze the genotypes of both populations. The QTL consistency in our comparison of F2:3 and BC2F2 populations was much lower than in previous studies using F2:3 and F6:7 generations, both for grain yield and yield components and for flowering and plant height (Veldboom et al. 1994; Veldboom and Lee 1994; Austin and Lee 1996, 1998). The previous experiments were conducted at the same locations but in different years. Also, in the F2:3 and F6:7 generation research, no selection was carried out during the development of F6:7 lines, and 150 F2:3 and 186 F6:7 lines, and 103 and 101 RFLP loci and one morphological marker were used, respectively, of which 84 loci were found in common between the two lines. Similar QTL consistency across generations was observed for grain yield and yield components, and for six flowering and plant height traits, with 22 of 35 (63%) of the regions associated with F2:3 QTL containing F6:7 QTL with corresponding parental effects (Austin and Lee 1996). Additionally, 16 (70%) of 23 regions associated with F6:7 QTL contained F2:3 QTL with the same parental alleles, and all were associated with increasing trait value.
Because the environmental effects were similar between the two studies in our comparison, the inconsistency in QTL detection might be attributable to two things. First, selection and genetic background may be important. Through simulation study, Moreno-Gonzalez (1993) showed that different generations had different efficiencies for estimating marker-associated QTL effects by multiple regression. Beavis et al. (1994) suggested that genetic background was one of the reasons for differences in QTL detection among two F3 backcrossed lines (Stuber et al. 1992), F4 lines and topcrossed populations from the same B73 × Mo17 cross. Genetic mapping with hybrid progeny across three testers and two generations (F2:3 and F6:7) for grain yield and grain moisture conducted by Austin et al. (2000) showed individual tester QTL effects were not consistent in rank or detection across generations. During the development of advanced backcross population, some non-recurrent introgressions might have been selected against (Septiningsih et al. 2003a, b; Yan and Gu 2000). By Mendelian expectation, only 25% of the plants in each BC2F2 family contained the donor parent allele for a given introgression. This “dilution effect” by non-carrier individuals could mask small effects of non-recurrent parent QTL alleles occurring at low frequencies (∼12.5%) (Ho et al. 2002). In our study, rigorous selection in BC1 and BC2 generations during BC2F2 population development might have caused abandonment of some elite genes, especially those coming from the non-recurrent parent, Also, large changes occurred in population structure, resulting from backcross and selection, with an allele frequency of 83% N04 to 15% Dan232 and a genotypic ratio of 68% N04/N04 homozygotes to 30% N04/Dan232 heterozygotes in this BC2F2 population. Meanwhile, an allele frequency of 50% N04 to 50% Dan232 and genotypic ratio of 25% N04/N04 homozygotes to 51% N04/Dan232 heterozygotes to 24% Dan232/Dan232 homozygotes occurred in the F2:3 population (Li et al. 2006b). Therefore, backcross and rigorous selection, causing changes in genetic background or population structure could cause inconsistencies in QTL detection.
The second reason for inconsistent QTL detection may be the mode of gene action. In an F2 population, recombination can dismantle the assembly of favorable epistatic gene combination accumulated by traditional breeding (Allard 1996; Ho et al. 2002). If high-order gene interactions play a significant role in the expression of key agronomic traits in maize, disruption of favorable epistatic interactions in an elite hybrid is minimized by initiating selection in an advanced backcross population. Because an advanced backcross population is skewed toward the recurrent parent genome, favorable epistatic interactions among recurrent parents are less likely to be disrupted. Conversely, there is a higher probability of detecting donor parent alleles with useful additive and dominant effects that will be expressed in a nearly isogenic background (Tanksley and Nelson 1996). This could be the major reason we observed slightly more QTL in the BC2F2 population than in the F2:3 population, suggesting a higher power for detecting QTL with additive effects in this BC2F2 population even through severe selection.
Austin and Lee (1996) considered that gene action accounts for some instances in which QTL were detected in the F2:3 but not in the F6:7 generation. QTL in the F2:3 with true overdominant gene action would not be expected to be detected if the additive effects were small. In their study for flowering and plant height traits, of the six QTL in the F2:3 with additive, partial dominant, or dominant gene action, five (83%) were detected in the same regions with the same parental effects as the F6:7 generation. Of the 17 QTL in the F2:3 with overdominant gene action, 11 (65%) were detected in the same regions with the same parental effects as the F6:7 generation. This trend, more consistent detection for F2:3 QTL with additive to dominant gene action versus those with overdominant gene action, was also observed for grain yield components in the same population (Austin and Lee 1998). In fact, QTL with dominant effects, might have their effects changed according to the homozygosity through backcross or inbreeding. Of the 16 QTL detected in our F2:3 population, the numbers of QTL expressing additive, partially dominant, dominant, and over-dominant effects were 2, 10, 2, and 2, respectively. Fourteen (88%) QTL with different degrees of dominant effects may play the most important role in their inconsistency across populations. Only the two QTL (100%) with additive effects and the four with partially dominant gene action (40%), were detected in the same regions with the same parental effects as the BC2F2 population. No QTL with dominant and overdominant gene action were detected in the same regions with the same or different parental effects as the BC2F2 population. Therefore, using QTL detected in early generations in separation, and maintaining good results in MAS or fine mapping or near-gene line development, requires caution. Careful attention should be given to QTL with additive gene actions. Generations developed through severe selection should have QTL redetected, and marker selection should be done in each population.
Comparison of QTL detected in the BC2F2 and other studies in normal maize
Because of differences in mapping populations (parents and progeny type), as well as a paucity of common loci and environments, direct comparisons of QTL mapping results across studies are difficult. One important consideration for QTL detection is the degree to which QTL location and effects from one population are observed in other populations (Lee 1995) or subsamples of the same population (Beavis 1994). Inconsistent detection of QTL may be the result of sampling variation (Beavis 1994), genetic heterogeneity of the phenotype (Beavis et al. 1991), and other factors (Lee 1995). Although studies in maize have indicated few QTL common among populations for most traits (Abler et al. 1991; Koester et al. 1993; Phillips et al. 1992), detection of some QTL in the same genomic regions across populations for some traits have been reported (Bubeck et al. 1993; Schön et al. 1994; Austin and Lee 1996, 1998; Yang et al. 2005; Tang et al. 2005). Comparing QTL data across different studies in maize can provide preliminary/suggestive information upon which some QTL alleles expressed in similar ways across multiple genetic backgrounds could be distinguished from those interacted with their genetic background and/or environments (Septiningsih et al. 2003a, b). QTL detection for grain yield components using popcorn inbreds has not been reported prior to this report. Comparison of the results in this study with previous results in normal maize may provide insight into the effect of popcorn background.
Several regions associated with QTL for GWP, 100GW, EL, RKN, ED, and ERN in our study have been reported previously, and compared with studies by previous researchers on yield and yield components in normal maize. Region 8L (8.03, phi115-bnlg1863) on chromosome 8 is associated with the QTL with the largest effects for GWP and 100KW (bnlg1863–umc2147), and QTL for ED, RKN herein. At the SSR marker loci of bnlg1834 (near bnlg1863)/phi115/phi119 (near phi115), Xiang et al. (2001), Lan et al. (2005), and Xiao et al. (2005) have detected QTL for GWP, RKN, grain weight per ear, or 100GW. QTL for grain yield, kernel weight/300-grain weight, ED, EL have also been found associated with umc48 or its near RFLP marker loci at bin 8.03 by Austin and Lee (1998), Austin et al. (2000), Melchinger et al. (1998), Stuber et al. (1992), and Veldboom and Lee (1994). At region 4L (4.05–4.06), QTL with the largest effects for EL (phi079–bnlg1621) and QTL for GWP (nc005–phi079) and ED (phi079–bnlg1621) have been detected in present study. Xiang et al. (2001), Yang et al. (2005), and Lan et al. (2005) have detected QTL for 1000KW/300KW/100KW on region 4L (4.05/4.08) at bnlg667/bnlg2162/php20608 (near bnlg1621). QTL for 300KW, EL, ED or ERN have been associated with region 4L (4.08) at npi410 or its near RFLP marker loci, umc15, npi292, and pio100025 (Austin and Lee 1996, 1998; Beavis et al. 1994; Veldboom and Lee 1994). Therefore, several regions detected in this study seem to contribute to trait variation in a diverse array of normal maize populations and environments.
QTL associated with multiple traits
Previous maize studies showed that correlated traits shared regions associated with QTL (Abler et al. 1991; Veldboom et al. 1994; Austin and Lee 1996, 1998; Tang et al. 2005; Yang et al.2005; Lan et al.2005). In this study, a total of 19 QTL were detected for the six grain yield components, and these QTL were located in 15 marker intervals comparing 26 loci. Of these loci, nine (33%) were associated with QTL for two to four traits. 100GW, EL, ED, and RKN, all highly positively correlated with GWP in phenotype and genotype, shared at least one QTL for RKN and both QTL for 100GW, EL and ED. Meanwhile, EL and ED, 100GW, ED and RKN, and ED and ERN shared one QTL each, and the QTL were located on chromosomes 4, 8, and 10, respectively.
Genetic explanations for multiple associations include QTL with pleiotropic effects or the presence of linked QTL controlling different traits. According to Austin and Lee (1996), the linkage pattern and parental effects of QTL for multiple traits in a region may indicate which of the two possibilities is more likely, although these two possibilities cannot be resolved in this type of study. In the marker interval nc005 ∼ bnlg1621 on chromosome 4, four QTL for three traits were detected in four marker intervals, with all favorable alleles contributed by the dent corn parent Dan232. One QTL for GWP was located in the marker interval nc005 ∼ phi079 (6.74 cM), one QTL for EL and one QTL for ED was in the marker interval phi079 ∼ bnlg1621 (13 cM). Because one QTL for EL was detected in the marker interval located with one QTL for ED, linked QTL controlling GWP, EL and ED could be present in this region (nc005 ∼ bnlg1621), which might be a cluster for QTL controlling different grain yield components. One QTL for 100GW and one for ED were detected in the near regions, bnlg1863 ∼ umc2147 and umc2147 ∼ umc1741, on chromosome 8, and one QTL for ED and one for ERN were in near regions phi059 ∼ umc2017 and umc2017 ∼ umc1877 on chromosome 10, respectively. It may be that two-linked QTL control two different traits in these two regions. In the marker interval umc2025 ∼ bnlg1811 on chromosome 1 and phi115 ∼ bnlg1863 on chromosome 8, QTL for GWP and 100GW, and QTL for GWP, ED and RKN were all detected, respectively, with all favorable alleles contributed by the dent corn parent Dan232. A single QTL could be present in these two regions with a pleiotropic effect on two or three traits, respectively. In our previous study with the F2:3 population for the same six traits, 10 of 22 marker loci or 5 of 11 marker intervals (45%) were associated with QTL for two traits. These results were higher than results in this study. QTL for the pairs ED and ERN, GWP and ED, and ED and ERN were all detected on the same marker interval—phi079 ∼ bnlg1621, phi109188 ∼ umc1221 and umc1677 ∼ umc2122 on chromosome 4, 5, and 10, respectively, with all favorable alleles contributed by Dan232. QTL for both EL and RKN, and both 100GW and ERN were located in the marker interval umc2332 ∼ phi069 and umc2057 ∼ umc1567 on chromosome 7, with their favorable alleles contributed by different parents. In the first case, a single QTL could be present in these three regions with a pleiotropic effect on two traits, while the two-linked QTL might be present to control two different traits in the later case.
Value of favorable QTL alleles for grain yield components in popcorn breeding
The breeding value of trait-improving QTL alleles depends on their possible association with negative effects on other traits, as well as their gene action and germplasm backgrounds. In popcorn breeding, normal maize inbreds can be used to improve popcorn germplasm in grain yield and yield component, plant, flowering and resistant traits (Crumbaker et al. 1949; Dofing et al. 1991; Ziegler and Ashman 1994; Li and Lu 2000; Li et al. 2002). However, elite popping characteristics of the popcorn germplasm must be recovered through 1 to 2 backcrosses with popcorn germplasm as recurrent parents (Dofing et al. 1991; Ziegler and Ashman 1994; Li et al. 2002). Because grain yield and most yield components are negatively correlated with popping characteristics (Dofing et al. 1990, 1991; Li et al. 1999), favorable QTL alleles for grain yield and yield components attributed by the normal corn parent should not have negative effects on popping characteristics. Comparing QTL detected for six grain yield components in this study with QTL for three popping characteristics using the same population (Niu 2006), two QTL for grain weight per ear and 100-grain weight, qBGWP1-1and qB100GW1-1 (umc2025 ∼ bnlg1811) and two QTL for PEV and popping rate (bnlg1811 ∼ umc2227), and two QTL for ear diameter and grain row number per ear, qBED10-1and qBERN10-1 (phi059 ∼ umc2017 and umc2017 ∼ umc1877) and one QTL for PEV (phi059 ∼ umc2017) were in the near or same marker intervals, with favorable alleles of three QTL for popping characteristics all contributed by the popcorn parent N04, while four QTL for grain yield components were contributed by the dent corn parent Dan232. In these two regions, a single QTL or linked multiple QTL could be present, with a pleiotropic effect on all three or four traits, or each controlling one or more traits. So, MAS for qBGWP1-1 or/and qB100GW1-1, and qBED10-1 or/and qBERN10-1 could accompany a decrease in popping characteristics. However, QTL for grain yield components and for popping volume detected on chromosomes 3, 5, and 8, with favorable alleles all contributed by Dan232, were very useful for large flake popcorn breeding. Although QTL for three grain yield components and for PEV were all detected on chromosome 4 with favorable alleles contributed by different parents, they could be interrupted through recombination and selection because their marker loci is 34.2 cM apart. One QTL for ED and one for popping volume detected on chromosomes 2 and 7, respectively, could be used in MAS.
Utility of BC2F2 families in further research and popcorn breeding
In this study, 220 BC2F2 families were maintained by selection in BC2 and BC2F2 according to GWP > N04, 100GW > N04, and PF ≥ N04. Of the 220 BC2F2 families, GWP of 210 families (95%) and 100GW of 208 families (95%) were higher than N04, and among these, the PEV of 35 families (16%) was nearly the same or higher than N04 (Niu 2006). After 1–2 generations of self-pollination, these 35 families could be used to replace N04 in popcorn breeding. This is the most significant advantage of advanced backcross generation over BC1 and self-developed generations (e.g., F2, F2:3, and RILs) in QTL mapping, especially in detection and transference of valuable QTL from wild or unadapted germplasm into elite breeding lines or varieties (Tanksley et al. 1996; Fulton et al. 1997, 2000; Bernacchi et al. 1998a, b; Xiao et al. 1998; Moncada et al. 2001; Septiningsih et al. 2003a, b; Huang et al. 2003; Xie et al. 2007). In QTL detection with F2:3 families derived from crosses between normal corn and popcorn inbreds, almost all of the F2:3 families could not be used directly in popcorn breeding, even after 2–3 generations of self-pollination and selection, because their popping characteristics were very poor (Li et al. 2005; Babu et al. 2006). Therefore, another cycle of backcross with popcorn germplasm must be carried out to use these lines to obtain elite popcorn inbreds.
Practice in normal maize breeding has revealed that increases in inbred yield have played a very important role in the increase in yield for hybrids, although heterosis is critical in maize breeding (Duvick 1992). Therefore, improvement in yield for popcorn inbreds could be helpful in increasing the yield for popcorn hybrids. The combining ability of self-selected lines from these BC2F2 families is currently being tested. In fact, the increase in 100GW for the popcorn parent N04 has special value in popcorn breeding, because hybrids with middle to large grain were much more favorable in the popcorn market in China. However, the grain of N04 is a little small, and the resulting hybrids belong to the small grain type, for example, 100GW of 9–12 g for Yubao2 (N04 × N10). Improving the grain weight of N04 would lead to increased utility in popcorn breeding.
In this study, the genotypic ratios of Dan232 of 18 families in all 220 BC2F2 families were lower than 10%. Among them, 12 families possessed QTL for GWP and/or 100GW, and seven families had QTL for popping characteristics, and their PEV was higher than N04. These families could be used to develop QTL-NILs for GWP and/or 100GW through selfing or another 1–2 backcross with N04, conducting MAS for target QTL and N04 germplasm background. Work is currently being done in this area. Expected QTL-NILs could be used to study their consistency of effects in environments, their interactions with environments, the interactions among them, fine mapping, and even their functions (Tanksley et al. 1996; Fulton et al. 2000; Huang et al. 2003; Brouwer and Clair 2004; Xie et al. 2007; Fan et al. 2006). In fact, successive backcross with the recurrent parent is the most efficient way to develop QTL-NILs for further research.
References
Abler SB, Edwards MD, Stuber CW (1991) Isoenzymatic identification of quantitative trait loci in crosses of elite maize inbreds. Crop Sci 31:267–274
Allard RW (1996) Genetic basis of the evolution of adaptedness in plants. Euphytica 92:1–11
Ashman RB (1991) Registration of three popcorn (Maize) parental lines, HP62-02, HP72-11, and HP68-07. Crop Sci 31:1402
Austin DF, Lee M (1996) Comparative mapping in F2:3 and F6:7 generations of quantitative trait loci for grain yield and yield components in maize. Theor Appl Genet 92:817–826
Austin DF, Lee M (1998) Detection of quantitative trait loci for grain yield and yield components in maize across generations in stress and non-stress environments. Crop Sci 38:1296–1308
Austin DF, Lee M, Veldboom LR, Hallauer AR (2000) Genetic mapping in maize with hybrid progeny across testers and generations: grain yield and grain moisture. Crop Sci 40:30–39
Babu R, Nair SK, Kumar A, Rao HS, Verma P, Gahala A, Singh IS, Gupta HS (2006) Mapping QTLs for popping ability in a popcorn × flint corn cross. Theor Appl Genet 112:1392–1399
Beavis WD (1994) The power and deceit of QTL experiments. In: Proceedings of the 49th annual corn and Sorghum research conference, Washington, DC, pp 250–266
Beavis WD, Grant D, Albertsen M, Fincher R (1991) Quantitative trait loci for plant height in four maize populations and their associations with quantitative genetic loci. Theor Appl Genet 83:141–145
Beavis WD, Smith OS, Grant D, Fincher R (1994) Identification of quantitative trait loci using a small sample of topcrossed and F4 progeny from maize. Crop Sci 34:882–892
Bernacchi D, Beck BT, Eshed Y, Lopez Y, Petiard V, Uhlig J, Zamir D, Tanksley SD (1998a) Advanced backcross QTL analysis in tomato. I. Identification of QTLs for traits of agronomic importance from Lycopersicon hirsutum. Theor Appl Genet 97:381–397
Bernacchi D, Beck-Bunn T, Emmatty D, Eshed Y, Inai S, Lopez Y, Petiard V, Sayama H, Uhlig J, Zamir D, Tanksley SD (1998b) Advanced backcross QTL analysis of tomato. II. Evaluation of near-isogenic lines carrying single-donor introgressions for desirable wild QTL-alleles derived from Lycopersicon hirsutum and L. pimpinellifolium. Theor Appl Genet 97:170–180, 1191–1196
Brouwer DJ, St Clair DA (2004) Fine mapping of three quantitative trait loci for late blight resistance in tomato using nearisogenic lines (NILs) and sub-NILs. Theor Appl Genet 104:628–638
Bubeck DM, Goodman MM, Beavis WD, Grant D (1993) Quantitative trait loci controlling resistance to gray leaf spot in maize. Crop Sci 33:838–847
Churchill GA, Doerge RW (1994) Empirical threshold values for quantitative trait mapping. Genetics 138:963–971
Crumbaker DE, Johnson IJ, Eldredge JC (1949) Inheritance of popping volume and associated characters in crosses between popcorn and dent corn. Agron J 41:207–212
Dofing SM, Thomas-Compton MA, Buck JS (1990) Genotype × popping method interaction for expansion volume in popcorn. Crop Sci 30:62–65
Dofing SM, D’Croz-Mason N, Thomas-Compton MA (1991) Inheritance of expansion volume and yield in two popcorn × dent corn crosses. Crop Sci 31:715–718
Duvick DN (1992) Genetic contributions to advances in yield of US maize. Maydica 37:69–79
Edwards MD, Page NJ (1994) Evaluation of marker-assisted selection through computer simulation. Theor Appl Genet 88:376–382
Fan CC, Xing YZ, Mao HL, Lu TT, Han B, Xu CG, Li XH, Zhang QF (2006) GS3, a major QTL for grain length and weight and minor QTL for grain width and thickness in rice, encodes a putative transmembrane protein. Theor Appl Genet 112:1164–1171
Fulton TM, Beck-Bunn T, Emmatty D, Eshed Y, Lopez J, Petiard V, Uhlig D, Zamir D, Tanksley SD (1997) QTL analysis of an advanced backcross of Lycopersion peruvinum to the cultivated tomato and comparisons with QTLs found in other wild species. Theor Appl Genet 95:881–894
Fulton TM, Grandillo S, Beck-Bunn T, Fridman E, Frampton A, Lopez J, Petiard V, Uhlig D, Zamir D, Tanksley SD (2000) Advanced backcross QTL analysis of a Lycopersion peruvinum × Lycopersicon parviflorum cross. Theor Appl Genet 100:1025–1042
Gimelfarb A, Lande R (1994) Simulation of marker assisted selection in hybrid populations. Genet Res Camb 63:39–47
Hallauer AR, Mirando FJB (1981) Quantitative genetics in maize breeding. Iowa State University Press, Ames
Ho JC, McCouch SR, Smith ME (2002) Improvement of hybrid yield by advanced backcross yield analysis in elite maize. Theor Appl Genet 105:440–448
Huang XQ, Cöster H, Ganal MW, Röder MS (2003) Advanced backcross QTL analysis for the identification of quantitative trait loci alleles from wild relatives of wheat (Triticum aestivum L.). Theor Appl Genet 106:1379–1389
Johnson IJ, Eldredge JC (1953) Performance of recovered popcorn inbred lines derived from outcrosses to dent corn. Agron J 45:105–110
Knapp SJ, Stroup WW, Ross WM (1985) Exact confidence intervals for heritability on progeny mean basis. Crop Sci 25:192–194
Koester RP, Sisco PH, Stuber CW (1993) Identification of quantitative trait loci controlling days to flowering and plant height in two near-isogenic lines of maize. Crop Sci 33:1209–1216
Lan JH, Li XH, Gao SR, Zhang BS, Zahng SH (2005) QTL analysis of yield components in maize under different environments. Acta Agron Sin 31:1253–1259
Lee M (1995) DNA markers and plant breeding programs. Adv Agron 55:265–344
Li YL, Xue XM, Jin YS, Hu XA (1999) Preliminary results of the single-cross breeding in popcorn. Chin Agric Sci Bull 15:11–12
Li YL, Lu ZJ (2000) Study on the expansion characteristics of the derived lines from popcorn × normal corn crosses. J Henan Agric Univ 34:210–212
Li YL, Lu FY, Du ZW, Wu SW, Han CP (2002) Preliminary report on improvement result to popcorn using normal corn germplasm and backcross. Acta Agric Boreali Sin 17:37–43
Li YL, Lu DB, Wu XJ, Dong YB (2005) The effect of successive selection on the self and backcrossing progenies from normal corn × popcorn crosses. Acta Agric Boreali Sin 20:12–16
Li YL, Dong YB, Cui DQ, Niu SZ, Wang YZ, Yu YL (2006a) QTL Mapping of popping characteristics in popcorn using the model of trisomic inheritance in the endosperm. Sci Agric Sin 39:448–455
Li YL, Dong YB, Niu SZ (2006b) QTL analysis of popping fold and the consistency of QTLs under two environments in popcorn. Acta Genet Sin 33:724–732
Lu HJ, Bernardo R, Ohm HW (2003) Mapping QTL for popping expansion volume in popcorn with simple sequence repeat markers. Theor Appl Genet 106:423–427
Melchinger AE, Utz HF, Schön CC (1998) Quantitative trait locus (QTL) mapping using different testers and independent population samples in maize reveals low power of QTL detection and large bias in estimates of QTL effects. Genetics 149:383–403
Moncada P, Martinez CP, Borrero J, Chatel M, Gauch H, Guimaraes E, Tohme J, McCouch SR (2001) Quantitative trait loci for yield and yield components in an Oryza sativa × Oryza rufipogon BC2F2 population evaluated in an upland environment. Theor Appl Genet 102:41–52
Moreno-Gonzalez J (1993) Efficiency on generations for estimating marker-associated QTL effects by multiple regression. Genetics 135:223–231
Niu SZ (2006) Advanced backcross QTL analysis and its application in genetics and breeding in popcorn. MS D Thesis. Henan Agricultural University, Zhengzhou, Henan, China
Phillips RL, Kim TS, Kaeppler SM, Parentoni SN, Shaver DL, Stucker RE, Openshaw SJ (1992) Genetic dissection of maturity using RFLPs. In: Proceedings of the 47th annual corn and Sorghum research conference, ASTA, Washington, pp 136–150
Pillen K, Zacharias A, Leon J (2003) Advanced backcross QTL analysis in barley (Hordeum vulgare L.). Theor Appl Genet 107:340–352
Robbins WA, Ashman RB (1984) Parent-offspring popping expansion correlations in progeny of dent corn × popcorn and flint corn × popcorn crosses. Crop Sci 24:119–121
Saghai Maroof MA, Soliman KM, Jorgensen RA, Allard RW (1984) Ribosomal DNA spacer length polymorphisms in barley: mendelian inheritance, chromosomal location, and population, and population dynamics. PNAS 81:8014–8018
SAS Institute Inc. (1989) SAS language guide. Release 6.03 edition. SAS Institute, Cary, NC, USA
Schön CC, Melchinger AE, Boppenmaier J, Brunklaus-Jung E, Herrmann RG, Seitzer JF (1994) RFLP mapping in maize: quantitative trait loci affecting testcross performance of elite European flint lines. Crop Sci 34:378–389
Senior ML, Heun M (1993) Mapping maize microsatellites and polymerase chain reaction confirmation of the targeted repeates using a CT primer. Genome 36:884–889
Septiningsih EM, Prasetiyono J, Lubis E, Tai TH, Tjubaryat T, Moeljopawiro S, McCouch SR (2003a) Identification of quantitative trait loci for yield and yield components in an advanced backcross population derived from the Oryza sativa variety IR64 and the wild relative O. rufipogon. Theor Appl Genet 107:1419–1432
Septiningsih EM, Trijatmiko KR, Moeljopawiro S, McCouch S (2003b) Identification of quantitative trait loci for grain quality in an advanced backcross population derived from the Oryza sativa variety IR64 and the wild relative O. rufipogon. Theor Appl Genet 107:1433–1441
Stuber CW, Lincoln SE, Wolff DW, Helentjaris T, Lander ESP (1992) Identification of genetic factors contributing to heterosis in a hybrid from two elite maize inbred lines using molecular markers. Genetics 132:823–839
Tang H, Yan JB, Huang YQ, Zheng YL, Li JS (2005) QTL mapping of five agronomic traits in maize. Acta Genet Sin 32:203–209
Tanksley SD, Nelson JC (1996) Advanced backcross QTL analysis: a method for the simultaneous discovery and transfer of valuable QTLs from unadapted germplasm into elite breeding lines. Theor Appl Genet 92:191–203
Tanksley SD, Grandillo S, Fulton TM, Zamir D, Eshed Y, Petiard V, Lopez J, Beck-Bunn T (1996) Advanced backcross QTL analysis in a cross between an elite processing line of tomato and its wild relative L. pimpinellifolium. Theor Appl Genet 92:213–224
Van Ooijen JW, Voorrips RE (2001) JoinMap 3.0, software for the calculation of genetic linkage maps. Plant Research International, Wageningen, The Netherlands
Veldboom LR, Lee M (1994) Molecular-marker facilitated studies of morphological traits in maize. II. Determination of QTLs for grain yield and yield components. Theor Appl Genet 88:451–458
Veldboom LR, Lee M, Woodman WL (1994) Molecular-marker facilitated studies in an elite maize population. I. Linkage analysis and determination of QTL for morphological traits. Theor Appl Genet 88:7–16
Wang S, Basten CJ, Zeng ZB (2006) Windows QTL cartographer 2.5. Department of statistics, North Carolina State University, Raleigh, NC (http://statgen.ncsu.edu/qtlcart/WQTLCart. htm)
Xiang DQ, Cao HH, Cao YG, Yang JP, Huang LJ, Wang SC, Dai JR (2001) Construction of a genetic map and location of quantitative yrait loci for yield component traits in maize by SSR markers. Acta Genet Sin 28:778–784
Xiao JH, Li J, Grandillo S, Ahn SN, Yuan L, Tanksley SD, McCouch SR (1998) Identification of trait-improving quantitative trait loci alleles from a wild rice relative, Oryze rufipogon. Genetics 150:899–909
Xiao YN, Li XH, George ML, Li MS, Zhang SH, Zheng YL (2005) Quantitative trait locus analysis of drought tolerance and yield in maize in China. Plant Mol Biol Rep 23:155–165
Xie XB, Song MH, Jin FX, Ahn SN, Suh JP, Hwang HG, McCouch SR (2007) Fine mapping of a grain weight quantitative trait locus on rice chromosome 8 using near-isogenic lines derived from a cross between Oryza sativa and Oryza rufipogon. Theor Appl Genet 113:885–894
Yan CJ, Gu MH (2000) Advanced backcross QTL analysis and its potential in rice high yield breeding. Hereditas 22:419–422
Yang JP, Rong TZ, Xiang DQ, Tang HT, Huang LJ, Dai JR (2005) QTL mapping of quantitative traits in maize. Acta Agron Sin 314:188–196
Zeng ZB (1993) Theoretical basis of separation of multiple linked gene effects on mapping quantitative trait loci. PNAS 90:10972–10976
Zeng ZB (1994) Precision mapping of quantitative trait loci. Genetics 136:1457–1468
Ziegler KE, Ashman B (1994) Popcorn. In: Hallauer AR (ed) Specialty corns. CRC, New York, pp 189–223
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by C. Hackett.
Rights and permissions
About this article
Cite this article
Li, Y.L., Niu, S.Z., Dong, Y.B. et al. Identification of trait-improving quantitative trait loci for grain yield components from a dent corn inbred line in an advanced backcross BC2F2 population and comparison with its F2:3 population in popcorn. Theor Appl Genet 115, 129–140 (2007). https://doi.org/10.1007/s00122-007-0549-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00122-007-0549-6