
Abstract
Although vocal mimicry in songbirds is well documented, little is known about the function of such mimicry. One possibility is that the mimic produces the vocalisations of predatory or aggressive species to deter potential predators or competitors. Alternatively, these sounds may be learned in error as a result of their acoustic properties such as structural simplicity. We determined the mimetic repertoires of a population of male spotted bowerbirds Ptilonorhynchus maculatus, a species that mimics predatory and aggressive species. Although male mimetic repertoires contained an overabundance of vocalisations produced by species that were generally aggressive, there was also a marked prevalence of mimicry of sounds that are associated with alarm such as predator calls, alarm calls and mobbing calls, irrespective of whether the species being mimicked was aggressive or not. We propose that it may be the alarming context in which these sounds are first heard that may lead both to their acquisition and to their later reproduction. We suggest that enhanced learning capability during acute stress may explain vocal mimicry in many species that mimic sounds associated with alarm.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Vocal mimicry occurs when a species learns a vocalisation from a heterospecific or an environmental source (Baylis 1982). Despite a variety of proposed explanations for this behaviour, the function of vocal mimicry in many bird species is poorly understood (reviewed in Kelley et al. 2008). Mimicry may be used to increase vocal repertoire size and thus increase the likelihood of attracting a mate (Coleman et al. 2007), to identify individuals within a population (Harcus 1977) or to increase foraging success (Flower 2010). Another hypothesis is that mimicry is used as a defence mechanism during predatory events, either by eliciting aid from a third party or by directly deterring/confusing the attacker (Dobkin 1979; Chu 2001). For mimicry to deter a potential attack it should cause the predator to mistake the identity of its prey for the presence of another predator (acoustic Batesian mimicry: Bates 1862; Dobkin 1979). Classically, Batesian vocal mimicry is used to describe the production of vocal mimicry of predators so as to deter predators but such mimicry may also be used to deter competitors (either con- or heterospecifics). An extension of this hypothesis is that mimicry of aggressive or competitor species (rather than predators) could be used to deter competitors (Rechten 1978; Gorissen et al. 2006). Here we refer to the mimicry of predators to deter predators as Batesian mimicry and the combination of this type of mimicry together with related uses of mimicry we refer to as “deterrent mimicry”.
The function of mimicry is often inferred through identification of the heterospecifics that are mimicked (models). For Batesian mimicry, the species mimicked are expected to be predators such as the production of rattlesnake Crotalus viridis rattles by burrowing owls Athene cunicularia (Rowe et al. 1986). There are also numerous anecdotal reports of female birds mimicking when protecting nestlings or eggs, which has been interpreted as deterrent mimicry (Chisholm 1932; Warham 1962). In yet other cases non-predatory species are mimicked such as the observation of nesting thick-billed euphonias (Euphonia violacea) mimicking the alarm calls of other species nesting nearby when disturbed (Morton 1976). In this case, deterrence may be achieved through apparent alarm calling rather than through mimicry of intimidating species. Therefore, the type of sounds (e.g., alarm calls, mobbing calls) that are mimicked as well as what model is being mimicked is important: alarm mimicry of non-threatening species may also act as a deterrent.
In all of these examples, however, it may be that it is the structure of the sounds that leads to their acquisition by the mimic. An alternative explanation to the functional hypotheses explaining mimicry is that it is the mechanism by which the sound is acquired that results in the mimetic repertoire. In particular, it has been proposed that the song learning system is especially susceptible to the mistaken acquisition of sounds that are simple, loud and common (the learning mistakes hypothesis: Hindmarsh 1986; Hausberger et al. 1991; Ferguson et al. 2002). It may be, therefore, that the calls of predators are mimicked as a result of the mimic’s learning mechanism rather than because of the potential use to which the mimic could apply that learning.
Although it is difficult to distinguish experimentally between these two kinds of explanations it may be possible to use the entirety of a mimic’s repertoire to do so, using a correlative approach to compare the origin and structure of mimicked and non-mimicked sounds. We recorded the mimetic repertoires of a population of individually identifiable male spotted bowerbirds Ptilonorhynchus maculatus during two sequential breeding seasons as these birds typically include predatory species in their mimetic repertoire (Warham 1962; Frith and Frith 2004; Coe 2005). If mimicry is used solely to deter predators or competitors then we would expect that only the vocalisations of predatory, aggressive or competitor species would be mimicked. If sounds are learned due to their acoustic properties then sounds with particular properties (for example short notes) that occur in the sound environment should be overrepresented in the mimetic repertoire (Hindmarsh 1986).
Materials and methods
We recorded the vocalisations of male bower-owning spotted bowerbirds in Taunton National Park (23.3° S, 149.1° E), central Queensland, Australia, between July and November 2007 and 2008. Individuals were identified by a unique series of colour bands on both legs. We recorded the vocalisations of 19 males in 2007 and 14 of these males were recorded again in 2008. Males were recorded vocalising at their bowers using a Sennheiser ME66/K6 microphone and a Sony TCD-D8 DAT recorder (sampling rate of 44.1 kHz and 16 bits resolution). All recordings were carried out by an observer sitting approximately 10 m from the bower and took place between 5:00 am and 2:00 pm. Recording sessions lasted between 3 and 7 h and the average time spent recording at each bower was 16 ± 1 h (mean ± SE) per bower in 2007 and 17 ± 2 h in 2008.
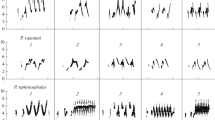
All recordings were converted into spectrograms using Raven Pro v1.3 (Charif et al. 2004) using a Hanning window and a 512-point fast Fourier transform. Suspected mimetic sounds were easily isolated due to their structural and acoustic dissimilarity to species-specific sounds, which are typically hisses, other harsh notes and mechanical noises (Frith and Frith 2004). Mimicry was then identified by listening to recordings and by visually inspecting spectrograms, and comparing them to recordings of their putative models using the Simpson and Day (1999) CD-ROM of bird sounds and sounds recorded opportunistically around bowers (as described in Kelley and Healy 2010; Fig. 1). Identification of mimetic sounds was confirmed by Dr J. A. Nicholls, an ornithologist with over 10 years experience of Australian bird vocalisations.

Spectrograms of model vocalisation (left) and bowerbird mimicry (right). a Yellow-throated miner Manorina flavigula mobbing calls; b willie wagtail Rhipidura leucophrys aggressive “rattle” call; c pied butcherbird Cracticus nigrogularis alarm call
We constructed repertoire profiles for all male bower owners in 2007 and 2008. Saturation curves were used to confirm that the entire mimetic repertoire had been captured. To determine the nature of mimicked sounds, we investigated what models bower owners mimicked from the suite of sounds available in their acoustic environment. We compiled a list of species present at Taunton using the Atlas of Australian Birds (using www.birdata.com.au, which is an online resource that lists all birds observed within a defined area) and checked that the listing matched up with species seen or heard during observations. Given that we cannot be sure of the exact acoustic habitat that each bird has been exposed to for the duration of their life we have used a bird atlas as an approximate measure of acoustic exposure. Of the 91 bird species present at Taunton, 15 species were mimicked by bowerbirds (excluding one human model) and 76 were not (see Appendix 1). All 91 species were then classified according to whether the intended receiver was a bowerbird or a heterospecific so as to determine whether mimicry may be acting as a deterrent towards conspecifics or heterospecifics.
In the first category (i.e., the intended receiver is a bowerbird), species were classified as predatory if they prey upon bowerbirds, aggressive if they behave aggressively towards bowerbirds (for example by displacing them from a perch) and as food competitors based on their diet. Spotted bowerbirds are primarily frugivores so any species whose main source of food was fruit and berries was classified as a competitor. Birds were classified using personal observations and using size, habitat and diet information from Simpson and Day (1999), and could be assigned to more than one group. In the second category, species were classified as to whether they were generally predatory or aggressive to non-bowerbird species (for example, the whistling kite Haliastur sphenurus is not a predator of bowerbirds but will prey on smaller birds). J. A. Nicholls also classified all species into these five groups without prior knowledge of whether they were mimicked or not. The only discrepancies in the categorisation of model species occurred in the competitor/non-competitor category, so the analysis was run twice using each set of classifications.
To address whether sounds with certain structural properties were preferentially mimicked, we measured the maximum frequency, minimum frequency, peak frequency and average duration of one note and total duration of the 91 possible models from recordings taken at Taunton and from professional recordings for species where no field recordings were obtained (Simpson and Day 1999). We also noted whether the model sounds had harmonics or trills and the size of the model bird. All terms were then entered into a binomial logistic regression. The data were manipulated and analysed using Excel and SPSS (v. 13).
The sounds mimicked were also classified according to their function in the model species. For example, a sound was classified as an alarm call if the model species produced it when alarmed. Given the abundance of the model species at the field site and the duration of observations, we observed the model species in a range of contexts and became familiar with their vocalisations. Our classifications were confirmed by sending samples of mimicry to researchers that worked on each species. Alarm mimicry was defined as the mimicry of a predator or the alarm, mobbing or aggressive calls of other species (Goodale and Kotagama 2006). Non-alarm mimicry was defined as sounds that are not associated with alarm, for example contact calls and song.
Results
Individual male bowerbirds varied in the size and composition of their mimetic repertoires in both 2007 and 2008. The range of models mimicked was zero to eight, and the most commonly mimicked species was the brown falcon Falco berigora. Although males varied in the number of models they mimicked each year, repertoire size did not increase or decrease consistently between years (paired t test t = −1.092, p = 0.29, n = 14).
Male bowerbirds preferentially mimicked species that were generally aggressive (Wald χ 2 = 4.83, p = 0.027; n = 18, Fig. 2; overall Wald χ 2 = 12.62, p < 0.001, Nagelkerke r 2 = 0.55). They also tended to mimic sounds with a high peak frequency (Wald χ 2 = 2.99, p = 0.08). No other category of model species was preferentially mimicked and sounds that were mimicked were not otherwise structurally different from sounds that were not mimicked (Table 1).

Size and composition of the mimetic repertoires of bower owners, each bar represents one bower owner. Data shown here are mimicked models that were classified as generally aggressive (black bars), both aggressive and predatory (grey bars) and other (white bars). All species that were predators, aggressors or competitors of bowerbirds are included in these data but are not identified for clarity
Of the species that were mimicked, sounds that were associated with alarm were common in mimetic repertoires, with alarm mimicry comprising 80.2% of the average male’s repertoire (Fig. 3). Only four non-alarm sounds were mimicked: the ‘babble’ contact call of the grey-crowned babbler Pomatostomus temporalis (mimicked by nine of the 19 males), the quiet notes at the start of the laughing kookaburra Dacelo novaeguineae “laugh” (mimicked by seven of 19 males) and two parrot contact calls (each mimicked by one male). Alarm-associated mimicry included mimicry of five predators, four alarm calls, two aggressive calls and one mobbing call.

Mimetic repertoires of males categorised by call type. Black bars represent alarm mimicry (mimicry of predators, alarm calls, mobbing calls and aggressive calls) and white bars represent non-alarm mimicry (contact calls and song)
Discussion
Male spotted bowerbirds preferentially mimicked species that were aggressive towards other birds. This is consistent with the prediction from the deterrent hypothesis that mimicry in this species could be used to deter predators or competitors. Given that sounds produced by predators or species that were aggressive to bowerbirds were not preferentially mimicked, it seems that the models mimicked might be more likely to be used to deter heterospecifics rather than directed towards conspecifics. As sounds that were mimicked were not more similar structurally than were sounds that were not mimicked, it does not appear that mimetic sounds are learned because of their acoustic structure (although mimicked sounds did tend to have higher peak frequencies than non-mimicked sounds).
Within and across males, the combination of sounds in each male’s mimetic repertoire seems best described as alarm-associated sounds. Mimicry of alarm calls and predators has been noted in several species (Robinson 1974; Snow 1974; Greenlaw et al. 1998) and these sounds are often produced in alarm contexts (Vernon 1973; Morton 1976; Goodale and Kotagama 2006; Ratnayake et al. 2010). Although it was originally proposed that these sounds were mimicked in an attempt to recruit assistance (Stefanski and Falls 1972), there is little evidence for or against this suggestion (Morton 1976; Chu 2001). Likewise, the suggestion that mimicry of alarm sounds may be used to startle a predator or to attract a second predator has little support as yet (Conover 1994).
The lack of structural similarity across mimicked sounds does not lend support to the learning mistakes hypothesis. However, given the propensity for these birds to acquire alarming sounds, we suggest that the context in which the sounds are learned may play a significant role in that acquisition. Specifically, we suggest that the calls of predators and heterospecific alarm calls are produced in contexts that may be acutely stressful. As learning and memory are typically enhanced during stressful events, alarming sounds may be especially likely to be acquired and remembered (Joëls et al. 2006). This could also explain the production of such sounds in future alarming contexts, i.e. they are reproduced when conditions are similar to those in which the sounds were learned (de Kloet et al. 1999; Goodale and Kotagama 2006; Ratnayake et al. 2010). Although there is considerable evidence that acute stress can enhance learning and memory during novel stressful events (e.g. Shors et al. 1992; Thaker et al. 2010), the role that stress may play in the enhancement or impairment of vocal learning has been less well studied. The little we do know concerns the negative impact of chronic stress rather than the striking effects of acute stress on learning: birds subjected to chronic stress during early development sing poorer songs as adults than do conspecifics that do not experience such developmental stress (e.g. Nowicki et al. 2002; Buchanan et al. 2003; Spencer et al. 2003). If acute stress does have this kind of effect on the acquisition of mimicry, it may be difficult to disentangle the cause of its production. To do this we will need to understand a lot more about the mechanistic basis of both the acquisition and the production of mimicry. Either natural selection has acted on a mimic’s cognitive abilities such that it “understands” a context and then produces the appropriate sound or natural selection has acted on a mimic’s sensory and hormonal apparatus such that the mimic produces alarming sounds when both are appropriately stimulated, i.e. conditioning (and, indeed, both may have occurred). One possible first step would be to determine whether it is the context or the sound itself that determines the learning of mimicry. For example, does an acutely stressful context enhance the learning of any sound? Alternatively, irrespective of context are alarm sounds learned more readily than non-alarm sounds? Given that several recent reviews have found little evidence for a function of mimicry in many species (Garamszegi et al. 2007; Kelley et al. 2008), we suggest that mechanistic factors such as those we describe here may explain the occurrence of mimicry in more species than previously assumed. It may also be the case that both learning mistakes and acute stress are responsible for the repertoires observed in many species.
When considering some of the alternative explanations for vocal mimicry, identification of the entire mimetic repertoire may be of limited use, but quantifying individual mimetic repertoires within a population and any changes over time can be helpful. With regard to inter-sexual selection, for example, while mimicry is incorporated into the display of the closely related satin bowerbird Ptilonorynchus violaceus, it is difficult to predict what sounds should be mimicked (Coleman et al. 2007). It is not immediately obvious why alarming sounds would be overrepresented in a male’s repertoire as we found in our spotted bowerbirds. However, changes in repertoire size do not support the mate attraction hypothesis: male spotted bowerbird mimetic repertoire size did not increase between years, as we would expect if females preferred males with large repertoires (Coleman et al. 2007; Zann and Dunstan 2008). We also never observed mimicry during sexual displays, so it seems unlikely that males use mimicry to attract mates. With regard to individual recognition, we would expect that bowerbirds within a population would have dissimilar repertoires, which was not the case (Thorpe 1961; Hindmarsh 1986). We would also predict that neighbouring individuals should have less similar mimetic repertoires, which they did not (Kelley and Healy 2010).
In conclusion, as vocal mimicry in wild birds is not readily amenable to experimental manipulation, we suggest observational techniques such as the recording and examining of a complete vocal repertoire, which is much used to investigate non-mimetic song (Beecher et al. 2000; Nowicki et al. 2000; Franco and Slabbekoorn 2009). This may prove to be of more use than such methods are often accorded nowadays. In the case of the spotted bowerbirds, at least, the examination of the composition, and of the structure of the sounds therein, of the mimetic repertoire does not contradict one of several hypotheses (the deterrence hypothesis) while concomitantly not supporting at least two other explanations (the sexual selection and individual recognition hypotheses). Furthermore, it has led to a possible mechanistic explanation as to why some sounds are preferentially acquired: that acute stress enhances vocal learning. Confirmation of this explanation requires learning experiments to demonstrate that birds are more amenable to learning sounds during or immediately following an acutely stressful stimulus.
References
Bates HW (1862) Contributions to an insect fauna of the Amazon Valley (Lepidoptera, Heliconidae). Trans Linn Soc Lond 23:495–566
Baylis JR (1982) Avian vocal mimicry: its function and evolution. Acoustic communication in birds. Academic Press, Inc, New York, pp 51–80
Beecher MD, Campbell SE et al (2000) Song-type matching between neighbouring song sparrows. Anim Behav 59:21–27
Buchanan KL, Spencer KA et al (2003) Song as an honest signal of past developmental stress in the European starling (Sturnus vulgaris). Proc R Soc B Biol Sci 270:1149–1156
Charif RA, Clark CW et al (2004) Raven 1.2 user’s manual. Cornell Laboratory of Ornithology, Ithaca
Chisholm AH (1932) Vocal mimicry among Australian birds. Ibis 13:605–625
Chu M (2001) Heterospecific responses to scream calls and vocal mimicry by phainopeplas (Phainopepla nitens) in distress. Behaviour 138:775–787
Coe RL (2005) Vocal mimicry in the spotted bowerbird Chlamydera maculata. University of Edinburgh, Edinburgh
Coleman SW, Patricelli GL et al (2007) Female preferences drive the evolution of mimetic accuracy in male sexual displays. Biol Lett 3(5):463–466
Conover MR (1994) Stimuli eliciting distress calls in adult passerines and response of predators and birds to their broadcast. Behaviour 131:19–37
De Kloet ER, Oitzl MS et al (1999) Stress and cognition: are corticosteroids good or bad guys? Trends Neurosci 22:422–426
Dobkin DS (1979) Functional and evolutionary relationships of vocal copying phenomena in birds. Z Fur Tierpsychol J Comp Ethol 50(4):348–363
Ferguson JWH, Van Zyl A et al (2002) Vocal mimicry in African Cossypha robin chats. J Fur Ornithologie 143(3):319–330
Flower T (2010) Fork-tailed drongos use deceptive mimicked alarm calls to steal food. Proc R Soc B Biol Sci. doi:10.1098/rspb.2010.1932
Franco P, Slabbekoorn H (2009) Repertoire size and composition in great tits: a flexibility test using playbacks. Anim Behav 77:261–269
Frith CB, Frith DW (2004) The bowerbirds. Oxford University Press, Oxford
Garamszegi LZ, Eens M et al (2007) A comparative study of the function of heterospecific vocal mimicry in European passerines. Behav Ecol 18:1001–1009
Goodale E, Kotagama SW (2006) Context-dependent vocal mimicry in a passerine bird. Proc R Soc B Biol Sci 273(1588):875–880
Gorissen L, Gorissen M et al (2006) Heterospecific song matching in two closely related songbirds (Parus major and P. caeruleus): great tits match blue tits but not vice versa. Behav Ecol Sociobiol 60:260–269
Greenlaw JS, Shackelford CE et al (1998) Call mimicry by eastern towhees and its significance in relation to auditory learning. Wilson Bull 110(3):431–434
Harcus JL (1977) Functions of mimicry in vocal behavior of chorister robin. Z Fur Tierpsychol J Comp Ethol 44(2):178–193
Hausberger M, Jenkins PF et al (1991) Species-specificity and mimicry in bird song—are they paradoxes—a re-evaluation of song mimicry in the European starling. Behaviour 117:53–81
Hindmarsh AM (1986) The functional significance of vocal mimicry in song. Behaviour 99:87–100
Joëls M, Pu Z et al (2006) Learning under stress: how does it work? Trends Cogn Sci 10(4):152–158
Kelley LA, Healy SD (2010) Vocal mimicry in male bowerbirds: who learns from whom? Biol Lett 6(5):626–629
Kelley LA, Coe RL et al (2008) Vocal mimicry in songbirds. Anim Behav 76:521–528
Morton ES (1976) Observations of vocal mimicry in the thick-billed euphonia. Wilson Bull 88(3):485–487
Nowicki S, Hasselquist D et al (2000) Nestling growth and song repertoire size in great reed warblers: evidence for song learning as an indicator mechanism in mate choice. Proc R Soc B Biol Sci 267:2419–2424
Nowicki S, Searcy WA et al (2002) Brain development, song learning and mate choice in birds: a review and experimental test of the "nutritional stress hypothesis". J Comp Physiol A 188:1003–1014
Ratnayake CP, Goodale E et al (2010) Two sympatric species of passerine birds imitate the same raptor calls in alarm contexts. Naturwissenschaften 97:103–108
Rechten C (1978) Interspecific mimicry in birdsong—does Beau Geste hypothesis apply? Anim Behav 26:305–306
Robinson FN (1974) The function of vocal mimicry in some avian displays. Emu 74:9–10
Rowe MP, Coss RG et al (1986) Rattlesnake rattles and burrowing owl hisses: a case of acoustic Batesian mimicry. Ethology 72:53–71
Shors TJ, Weiss C et al (1992) Stress-induced facilitation of classical conditioning. Science 257:537–539
Simpson K and Day N (1999) Field guide to the birds of Australia. Penguin Group
Snow BK (1974) Vocal mimicry in the violaceous euphonia, Euphonia violacea. Wilson Bull 86(2):179–180
Spencer KA, Buchanan KL et al (2003) Song as an honest indicator or developmental history in the zebra finch (Taeniopygia guttata). Horm Behav 44:132–139
Stefanski RA, Falls JB (1972) A study of distress calls of song, swamp and white-throated sparrows (Aves: Fringillidae) I. Intraspecific responses and functions. Can J Zool 50(12):1501–1502
Thaker M, Vanak AT et al (2010) Stress and aversive learning in a wild vertebrate: the role of corticosterone in mediating escape from a novel stressor. Am Nat 175(1):50–60
Thorpe WH (1961) Bird song. Cambridge University Press, Cambridge
Vernon CJ (1973) Vocal imitation by southern African birds. Ostrich 44:23–30
Warham J (1962) Field notes on Australian bowerbirds and catbirds. Emu 62:1–30
Zann R, Dunstan E (2008) Mimetic song in superb lyrebirds: species mimicked and mimetic accuracy in different populations and age classes. Anim Behav 76:1043–1054
Acknowledgements
We would like to thank James Nicholls for his assistance both in and out of the field. We are grateful to QPWS for field site access and our field assistants for all their help. We thank Anne Goldizen for assistance with ethics and Tom Little for statistical advice. We thank two anonymous reviewers for their useful comments on this manuscript. LAK would like to thank NERC for funding. This research was approved by the University of Queensland Animal Ethics Committee (SIB/272/07/NERC & SIB/326/08/NERC) and the Queensland Environmental Protection Agency (WISP04545307 and WITK04545407).
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by: Sven Thatje
Electronic supplementary material
Below is the link to the electronic supplementary material.
ESM 1
(PDF 93 kb)
Rights and permissions
About this article
Cite this article
Kelley, L.A., Healy, S.D. The mimetic repertoire of the spotted bowerbird Ptilonorhynchus maculatus . Naturwissenschaften 98, 501–507 (2011). https://doi.org/10.1007/s00114-011-0794-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00114-011-0794-z