
Abstract
To date, song research has focused primarily on the interactions of conspecifics. However, frequent interactions of songbirds with heterospecifics may necessitate adequate communication outside the species boundary. In this study, we focus on heterospecific communication behaviour of two small sympatric congeneric passerines, great and blue tits (Parus major and Parus caeruleus), which breed in overlapping territories and compete for food and nesting cavities. By means of a first playback experiment, we show that (1) heterospecific matching (imitating songs of the other species) is a strategy frequently used by great tits but not by blue tits, (2) both blue tit trilled and untrilled song can be accurately matched by great tits and that (3) almost half of the great tits in our study population match at least one blue tit song across all studied breeding stages, indicating that this heterospecific matching behaviour is a common feature in this population. A second playback experiment showed that these great tit imitations of blue tit songs do not function in intraspecific communication between male great tits. Hence, these heterospecific imitations appear to be designed for interspecific communication with blue tits. These findings suggest a strong heterospecific influence on the vocal learning process, repertoire composition and repertoire use in great tits and provide a possible mechanism that can drive song convergence in songbirds.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
In all animal societies, communication plays an essential role in survival and reproduction. To date, research has focussed primarily on communication strategies between members of the same species. However, members of one’s own species are not the only individuals that have access to signals. Heterospecifics may also be capable of capitalising on signal-associated cues and communication involving only intraspecies effects is apt to be the exception rather than the rule (see Kostan 2002). Vocal communication in birds is well developed and one of the most consistent and striking characteristics of bird vocalisations is its species’ specificity. Interspecific discrimination of sounds minimises unnecessary expenditures of time and energy, interspecific conflicts and hybridisation and in general makes intraspecific vocal communication more efficient (Becker 1982). Apart from obvious intraspecific functions (e.g. mate attraction, territory defence), avian vocalisations can have important interspecific functions too, as in parasite–host relationships or the sharing of alarm calls between different songbirds (e.g. Morton 1976; Baylis 1982).
When resources are limited, competition between species may result in a need for interspecies communication skills, and thus may, like intraspecific competition for territories and mates, affect signal design and vocal differentiation. An increasing number of studies have shown that vocal signals play an important role in interspecific territoriality between species (e.g. Reed 1982; Sorjonen 1986; Martin et al. 1996; Martin and Martin 2001; Secondi et al. 2003). Birds that often occupy territories adjacent to congeners might well learn to recognise and react to vocal signals of invading heterospecific competitors who are regularly heard and encountered in competitive interactions (Catchpole 1978). Social encounters with heterospecifics can even promote signal convergence between species (e.g. Cody 1973; Secondi et al. 2003; Haavie et al. 2004) because it may increase the efficiency of territorial signalling directed towards heterospecifics. Exclusion of competitors can be achieved by direct attack or physical aggression. Obviously, such reactions are time and energy demanding and they can lead to life-threatening injuries. Hence, most songbirds have developed a less costly way to expel competitors, namely through vocal signalling. Two examples of vocal signalling behaviour between members of the same species are singing on top of the songs of a competitor (overlapping) or singing the same song as the intruder (matching; e.g. Krebs et al. 1981a,b; Dabelsteen et al. 1996; 1997).
Between members of the same species, vocal matching occurs when a signaller changes some features of its singing behaviour to resemble more closely a preceding song from another individual (McGregor et al. 1992). Hence, matching is a method of directing an omnidirectional signal to a specific rival singing male (Brémond 1968) and several studies in different bird species provide strong evidence that vocal matching is a more threatening singing behaviour than is vocal non-matching (Todt and Naguib 2000). If this vocal strategy is effective in intraspecific communication, it might be possible that this matching behaviour might prove to be effective in interspecific communication as well. Possible evidence for heterospecific matching comes from studies describing different ways of imitating sounds of other species defined as vocal mimicry, vocal copying, vocal imitation (e.g. Dobkin 1979; Baylis 1982; Ferguson et al. 2002) and mixed singing (Sorjonen 1986; Catchpole and Baptista 1988; Baptista and Catchpole 1989; Osiejuk and Kuczynski 2000, 2003; Owen-Ashley et al. 2002; Haavie et al. 2004). Vocal signals of other species can be partially or wholly imitated with low to high resemblance. It may play an important role in interspecific competition, whilst usually it is regarded only as an aberration deprived of functional significance (Catchpole and Slater 1995). Imitating vocal signals of other species is undoubtedly more probable in closely related species and could increase the efficiency of interspecific territoriality as well as reduce intraspecific song functionality (see Osiejuk and Kuczynski 2000). Hence, singing heterospecific songs, might be a more specialised form of matched singing in some species.
Matching songs of interspecifically territorial and sympatric species might be a way of capturing the attention of intruding heterospecifics to maintain and defend an interspecific territory (see Baylis 1982). Because most birds predominantly pay attention to the vocal signals of their own species (Hailman 1977), phrasing a threat in their own language might be the most effective method of chasing heterospecifics out of one’s territory (Baylis 1982). For heterospecific matching to occur, a number of conditions need to be fulfilled first: both species have to be sympatric and one species has to be interspecifically aggressive towards the other species. Matching should be part of the communication skills of at least one species and there should be a minimal overlap between the sound characteristics of both species (e.g. frequency range, element morphology) that allows matching to occur, as often is the case between close relatives.
Two interesting species to study in this regard are great and blue tits (Parus major and Parus caeruleus). They occur sympatrically throughout Europe. They are closely related, compete for food and nesting cavities and respond to the opposite species’ sounds (Dhondt 1977; Perrins 1979; Cramp and Perrins 1993; Schottler 1995; Slikas et al. 1996; Doutrelant et al. 1998, 2000; Törok and Toth 1999). Vocalisations of great tits can be classified as either songs or calls. The basic unit of a great tit song is a phrase, which is a fixed combination of one to more than five notes. A song is usually build up by a sequence of two to 20 stereotypically repeated phrases that constitute a strophe (Lambrechts 1996). Calls usually differ from songs in that they lack the repetition of stereotypically repeated phrases (mostly only one or two phrases), they are shorter than songs (have lower number of notes) and have broader frequency ranges (Gorissen 2005). The vocalisations of male blue tits can be classified into two categories: song types with a trill (trilled song) and song types without a trill (untrilled song). A trill is a series of identical notes in the last subphrase of a strophe, rapidly repeated in a sequence (e.g. Doutrelant et al. 2000; Gorissen and Eens 2004). Untrilled songs can be divided into single-phrased or multiple-phrased song types (Gorissen 2005). A large-scale playback experiment with great tits showed that great tits respond equally aggressive towards blue tit untrilled song and conspecific song, whilst the response to blue tit trilled song is significantly lower (Doutrelant et al. 2000). Moreover, the incidence and/or proportion of trilled song types in the blue tit repertoire appear to be correlated with the presence/abundance of great tits (Doutrelant and Lambrechts 2001). Hence, interspecific interference with the more dominant great tits may promote song divergence in blue tits (Character shift hypothesis; Doutrelant et al. 2000).
While blue tit trilled song and great tit vocalisations are differentiated in most sound features, great tit calls and blue tit untrilled song resemble each other in many sound characteristics (Gorissen 2005, Figs. 1 and 2). In addition, great tits have been reported to use calls resembling other Parus and non-Parus species’ calls (Cramp and Perrins 1993). Hence, the opportunity for heterospecific matching behaviour to evolve is at hand. In both species, matching conspecific song is part of their vocal behaviour and, in great tits, this matching behaviour is associated with approach and aggressive encounters (e.g. Krebs et al. 1981a,b; Falls et al. 1982; McGregor et al. 1992; Poesel et al. 2004). Hence, these species provide a great opportunity to investigate interspecific communication strategies. By means of a first playback experiment, we investigated whether blue and great tits imitate vocal signals of the opposite species when confronted with heterospecific playback stimuli (heterospecific matching). We predict that if heterospecific matching occurs, that it will be most pronounced in the more dominant and superior species, the great tit. By means of a second playback experiment, we investigated whether matching the opposite species’ sounds might function in interspecific communication by studying intra- and interspecies functionality of these heterospecific imitations.

An example of a blue tit trilled song (a) and a great tit song (b) that were used as playback stimuli for “Experiment 1”. All vocal signals produced by the focal bird were compared to the playback stimuli in five different dimensions: (1) Frequency range, (2) Frequency pattern, (3) Syntactical order, (4) Structure and (5) Note morphology

Sonograms of blue tit songs followed by a great tit vocalisation. According to the classification scale used, the accuracy of matching is very high in a with a score of 5, high in b and c with a score of 4, intermediate in d with a score of 3 and is zero in e. The blue tit songs of a, d and e are trilled songs, whilst b and c are untrilled song types. The great tit imitations of a, b and c are examples of the song stimuli used in the second playback experiment
Materials and methods
Study species and study area
Great and blue tits are sympatric congeneric territorial passerines that breed in secondary holes and artificial nest boxes in all types of wooded areas throughout Europe and parts of Asia and North Africa (Cramp and Perrins 1993; Slikas et al. 1996). Coexistence of blue and great tits has important fitness consequences for both species. It has been shown experimentally that the larger and more dominant great tits prevent blue tits from roosting in (large-holed) nest boxes that are accessible for both species (Kempenaers and Dhondt 1991) and earlier studies showed that the presence and abundance of blue tits have a negative impact on great tit nestling weight, suggesting that blue tits reduce great tits’ access to food (Minot 1981; Törok and Toth 1999). The study area is situated in the nature reserve Fort 7 in Antwerp, (Belgium). This study site has a size of about 30 ha and is, since 1999, provided with 36 nest boxes with small entrance hole (28 mm) for blue tits and 27 nest boxes with large entrance hole (32 mm) for great tits. From March 12, 2002 to March 17, 2004, nest boxes were checked frequently to follow the nest building, egg laying and incubation stage of blue and great tits and the majority of birds had been individually marked earlier, with a numbered metal ring and a unique combination of colour rings. The relative great tit density in our study area measured as the number of breeding great tit pairs divided by the number of breeding great and blue tit pairs equalled 0.6 in 2002 and 0.53 in 2004.
Playback experiments
Experiment 1
To investigate whether great and blue tits can imitate sounds of the opposite species, we performed a playback experiment in 2002 in which we exposed 19 territorial great tit males and 17 territorial blue tit males to conspecific and heterospecific song stimuli (blue tit trilled song, blue tit untrilled song and great tit song) during three different stages of the breeding cycle (nest building, egg laying and incubation) from March 15 till May 13. Playback trials were conducted in the morning (between 09.00 and 13.00 h) and were postponed until the next day if weather conditions were not favourable (e.g. heavy wind or rain). Each playback experiment lasted for at least 25 min during which the focal male was followed continuously and all the three song stimuli were broadcast in random order to control for environmental influences, factors of time and inter-subject variation.
Each playback experiment consisted of three trials during which a song stimulus was broadcast at the natural species’ specific sound pressure level (which is 94±1 dB (A) for great tits and 80±1 dB (A) for blue tits at 1 m; measured by an Extech SPL meter with 1.5 dB accuracy, 0.1 dB resolution and measuring range between 35–130 dB) during 3 min at 2 m from the nest box and followed by 2 min of silence. Two trials were separated by a break of 5 min or more until the focal bird lost interest in the song stimulus and resumed its normal behaviour. To investigate whether response to one stimulus carried over into the next, we tested for effects of song stimulus order in our statistical design. The mini-vox Anchor PB-25 speaker (maximum SPL at 1 m is 108 dB, frequency response between 100 and 12 kHz) was placed approximately 1.5 m above the ground and connected to a portable Panasonic SL-SX270 cd player, which was placed at a distance of 15 m from the nest box and controlled by one investigator whilst the second investigator recorded all sounds produced by the focal bird during each trial using a Sennheiser ME 67 shotgun microphone, connected to a portable SONY MZ-R700 Minidisc Recorder.
To limit problems of pseudoreplication (see Kroodsma et al. 2001), between 9–14 different trilled and untrilled blue tit songs and great tit songs, all recorded during the dawn chorus of as many different males in Antwerp, were used as song stimuli. A given playback stimulus was played as a minimum to one and as a maximum to three focal birds, in the latter case all of which were spatially widely separated. Details about recording procedure and equipment and spectral analysis are described in Gorissen and Eens (2004). The strophe length of the playback stimuli varied within the species specific natural range (between 0.94 and 2.30 for blue tit trilled songs, 0.53 and 2.10 for blue tit untrilled songs and between 1.44 and 3.80 for great tit songs; Gorissen 2005), which differs from the playback experiment performed by Doutrelant et al. (2000) in other blue tit populations where strophe lengths of the playback stimuli of both species were of equal length (see also Gorissen 2005). Ten successive strophes originating from spontaneous song recordings were separated by pauses approximating the species specific natural song rate (0.33±0.05) of this population, which were looped during the playback trial (unpublished data). During the nest building stage, we often attracted blue tits of an adjacent territory to a great tit territory when broadcasting blue tit song and vice versa. We were not able to exclude these trials because both species have been reported to live in overlapping territories (Dhondt and Eyckerman 1980; Doutrelant et al. 1998) and taking into account only those trials where territories do not overlap, is not an accurate reflection of the natural situation in our population. Because a live intruder may influence the focal male’s behaviour, we compared the focal bird’s singing behaviour in these instances to the vocalisations produced by the intruder as well as to the playback stimulus.
Because blue and great tits differ in body and bill size (Cramp and Perrins 1993), and birds of a given size are physically unable to produce vocalisations beyond particular frequency ranges, durations and repetition rates (Ryan and Brenowitz 1985; Nowicki et al. 1992), the scoring of song matching between species is far more difficult than within species. Hence, a perfect match as defined within species, might not be possible when comparing sounds between species. We already know that blue tit untrilled songs show a close resemblance with great tit calls in parameters of the time, frequency and syntactical domain whilst blue tit trilled songs are very dissimilar from great tit sounds (Gorissen 2005). Hence, both species are able to match some acoustic characteristics of the other species’ sounds. But are they able to produce vocal imitations of the opposite species? Because vocal signals of other species can be partially or wholly imitated with low to high resemblance, we quantified the accuracy of matching of conspecific or heterospecific song by comparing a printed sonogram of the focal bird’s vocal signals to a printed sonogram of the playback stimulus or in the case a heterospecific bird was attracted to the song stimulus and was singing in the territory, also to a printed sonogram of its song (see Falls et al. 1982; Beecher et al. 2000; Molles and Vehrencamp 2001). The following sound features were chosen for this comparison because of their possible importance as species-specific parameters (unpublished data; Becker 1982); (1) structure of song, (2) frequency range of song, (3) frequency pattern of song, (4) similarity in note morphology and (5) syntactical order (see Figs. 1 and 2).
A note is the smallest element shown on a spectogram, always followed by an interval of silence. Frequency pattern is the pattern of the frequency shifts along the notes, covering the whole frequency range. Syntactical order is the sequence of notes of different morphology and song structure represents the unique combination of all notes (Fig. 1). The difference between syntactical order and structure can best be explained as follows: if we would give all notes with different morphology, a different letter, the syntactical order would be abc whilst the structure would be abbcccccccccc for the blue tit song and ababababab and aabaabaabaabaab, respectively, for the great tit song presented in Fig. 1. Hence, structure gives information about phrase composition (see “Introduction”). The sounds produced by the focal bird were scored 0 in case of no overlap or 1 in case of overlap in the above described five different sound features. Overlap means that there is a close resemblance between the playback stimulus and the sound feature of the focal bird whereas no overlap means that the quantified sound feature is very dissimilar from that of the playback stimulus.
Thus, each sound produced by the focal bird received a score between 0 and 5. All vocalisations were scored by one experienced and one inexperienced observer. As a measure of agreement, we calculated the effective percentage of agreement of vocal match (a score of 4 or 5) or non-match (a score of 3 or less) as the number of agreements between both observers divided by the sum of the number of agreements and disagreements (see Jones et al. 2001). Of the 228 comparisons made, 30 were classified as disagreement leading to a percentage of agreement of 88.3%. Still, care should be taken when interpreting interobserver reliability, for classification by eye is limited by subjectivity and high scores of reliability do not always indicate high validity (see Jones et al. 2001). All vocal signals produced during a 15-min period after the start of the playback stimulus were taken into account. When the focal bird produced more than one vocal signal (apart from scolding), we compared all vocal signals to the playback stimuli. Because we are not interested in the average score of matching, only the sound with the highest score was used in our statistical model. This method takes into account an unknown but wide range of sound features, but does not consider how the birds themselves classify avian sounds. Furthermore, it is possible that this set of sound features leaves out some defining aspect of the signals such as song amplitude or strophe length for instance. Hence, although the method used may not be sensitive enough to demonstrate more subtle ways of matching, it is a first step in studying heterospecific song matching.
Experiment 2
To investigate whether copying other species’ sounds might function in interspecies communication, we examined whether imitations and non-imitations evoked different behavioural responses by the species that was imitated as well as whether these imitations reduced intraspecific song functionality in the imitating species. To achieve this, a second playback experiment was conducted between April 1 and April 14 during the dawn chorus (between 05.30 and 07.30 h) in the egg-laying stage in 2004, in which we quantified the behavioural response of 14 blue and great tit males to three different vocal stimuli. We choose the dawn chorus because in this period, male great and blue tits sing vigorously in the vicinity of their female’s nest box and are not involved in other activities (such as foraging or resting). This allowed us to control for the effect of activity when quantifying the response of the focal bird. The protocol of this experiment was very similar to that described above. Three stimuli were broadcast in random order and the speaker was placed in a tree at about 1.5–2 m height at approximately 5 m from the nest box in a blue tit territory that bordered on a great tits’ territory (approximately 20–30 m from the great tit’s nest box). Each trial lasted for 3 min during which vocal signals were broadcast and trials were separated by 5 min of silence or more. Approximately 20 min before the start of the dawn chorus, a researcher concealed him or herself at a distance of 10 m from the nest box from where the portable cd player was operated.
The sound stimuli used were (1) normal great tit calls, (2) great tit imitations of blue tit songs and (3) wren songs, which were used as a control. The normal great tit calls and great tit imitations of blue tit songs with a score of 4 or 5 as agreed by both observers had been recorded during the playback experiment in 2002 and only high quality sounds, recorded from less than 10 m were chosen (see Fig. 3). The wren songs were recorded during the dawn chorus in the same population in 2002. Within each stimulus, we used six different songs/calls to minimise problems of pseudoreplication. All vocal signals were separated by 2 s intervals and the length of all vocal signals varied within the species’ specific range (Gorissen 2005, Fig. 3). Because great tit imitations of blue tit songs are very different from great tit songs whilst the structure and length of these imitations are more comparable to great tit calls, we used normal great tit calls to compare great tit response to great tit matching blue tit songs (unpublished data). We believe that using normal great tit calls in this experiment has no major impact on the results because of the following reasons: in an earlier experiment we played back great tit song in blue tit territories at dawn and the approach response of neighbouring great tits was similar to the approach response in this experiment (Gorissen 2002 and unpublished data). Secondly, in our study population, great tits call and sing in quite the same way (repetitions of calls alternate with repetitions of song) throughout their dawn chorus (see also Gorissen and Eens 2004).

Stimuli used in the second playback experiment. a Two examples of wren songs. b Six great tit imitations of blue tit songs. c Six great tit calls
During these playback experiments, it became evident that our playback design was not suitable to objectively quantify blue tit response towards these three playback stimuli for the following reasons: (1) in contrast to great tit males, several blue tit males did not sing their dawn chorus in close proximity of their nest box (and hence the playback speaker) which did not allow us to quantify their behavioural response and (2) in 10 of 14 playback trials, we attracted a neighbouring great tit to the speaker when broadcasting great tit calls, which may have confounded blue tit’s response to the playback stimulus because in normal conditions, blue and great tits do not approach each other during the dawn chorus (unpublished data). For these reasons, data interpretation on the functionality of heterospecific matching is not possible, and therefore, we refrained from integrating blue tit response in the results.
To investigate intraspecific functionality of these imitations in great tits, we quantified the approach response of great tit neighbours to these three different vocal playback stimuli. When the great tit approached the speaker and the distance between both was less than 10 m during the 3-min playback, a score of 1 was assigned. When the great tit did not approach the speaker and when the distance was more than 10 m, a score of 0 was assigned. In total, the approach response of 14 great tits was quantified. Only two of these great tit males were present in this study site in 2002; thus, no song or call of these two males were used as playback stimuli (they did not produce imitations of blue tit sounds).
Statistical analyses
To investigate the importance of playback stimulus, species, period and order of the song stimuli for the matching behaviour of great and blue tits (Experiment 1), data were analysed using generalised linear mixed models (GLMM) using the SAS for Windows software (version 8.1, 1999–2000). Because matching behaviour is a variable with a Poisson response, a GLIMMIX macro model allowed the use of a GLMM with Poisson errors and log link function. Interactions and non-significant effects were excluded from the model using a backward deletion procedure. To investigate whether the great tit’s approach response towards wren song, great tit calls and great tit imitations of blue tit songs that were broadcast in an adjacent blue tit territory during the dawn chorus differed between these three playback stimuli, a Fisher’s Exact Test was performed using the software StatXact-3 (Experiment 2).
Results
Experiment 1
During the playback experiment in 2002, we recorded in total 12 great tit imitations of blue tit song from nine different individuals; seven during nest building, two during egg-laying and three during incubation. Great tits matched blue tit trilled song as well as blue tit untrilled song (Fig. 2). On the other hand, no blue tit imitation of great tit song was recorded. Hence, heterospecific matching appears to be a vocal strategy used by great tits but not by blue tits (Fisher Exact Test, p=0.029). Statistical analysis of the matching behaviour of blue and great tits using GLMM models, showed no significant effect of order and period on the matching behaviour of great and blue tits towards the three different song stimuli but the species–stimulus interaction was significant (GLMM; F 2,202; F=18.80; p<0.001) indicating that the accuracy of matching of a particular song is species-dependent. Tukey post hoc tests (see Table 1) show that great tits are significantly more accurate in matching great tit song than blue tits (t=5.38; df=202; p<0.0001), whereas blue tits are significantly more accurate in matching blue tit trilled song than great tits (t=−2.55; df=202; p=0.012) as expected. The accuracy of matching blue tit untrilled song is not significantly different between great and blue tits (t=−1.49; df=202; p=0.14) or, in other words, great tits are, according to the classification scale used, as accurate in matching blue tit untrilled song as the blue tits themselves. Great tits are better in matching blue tit untrilled song than in matching trilled song, although this difference is not significant (t=−0.61; df=202; p=0.54; Table 1). Also, our results show that great tits are better in matching blue tit songs than blue tits in matching great tit songs (t=2.85; df=202; p=0.0048).
Experiment 2
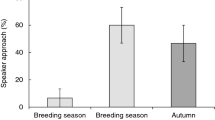
Three out of 14 great tits did not respond to either of the three song stimuli broadcast during the dawn chorus. Only one great tit approached the speaker when broadcasting wren song or great tit imitations of blue tit songs whilst 10 out of 14 great tits approached the speaker when broadcasting normal great tit calls, thereby leaving the proximity of their females’ nest box or even leaving their own territory (Fisher’s Exact Test, p=0.0002). The average approach response (quantified as the distance between focal bird and speaker) to the playback speaker was 3.87±0.55 m (range 2–6 m).
Discussion
The first playback experiment clearly demonstrates the ability of blue and great tits to distinguish between and respond with some specificity, to conspecific and heterospecific song. The non-significant effects of period and order of the song stimuli show that the stage of breeding and the temporal position of each playback stimulus relative to the other stimuli have no effect on the matching behaviour of great and blue tits. Our results show that heterospecific matching is a strategy frequently used by great tits but not by blue tits and because about 47% of the great tits studied accurately match at least one blue tit song across all periods according to our classification scale, this heterospecific matching behaviour appears to be quite common in this population. The fact that some great tits matched blue tit trilled songs, which are clearly of different syntax than great tit vocalisations (Gorissen 2005), has never been reported before and clearly indicates that great tits do not merely use those sounds in their repertoire that resemble blue tit sounds, but that they actively imitate blue tit songs that are very different from conspecific sounds.
Blue and great tits compete for resources such as food and nesting cavities and their coexistence has important fitness consequences for both species as mentioned before. These fitness costs of coexistence might promote interspecific aggression because resources need to be defended against heterospecifics as well. So the use of efficient vocal strategies such as matching might prove to be effective in interspecific communication. Hence, heterospecific matching could be a way of phrasing a threat in the language of the heterospecfic intruder. But why is this heterospecific matching behaviour common in great tits and uncommon in blue tits? Perhaps learning to produce great tit song is impossible for blue tits because of possible constraints in morphology and/or anatomy because they are notably smaller than great tits. Otherwise, matching heterospecific song might be effective only in the more dominant species because they are physically able to deter and overcome the smaller heterospecifics whilst the opposite is not the case (see Robinson and Terborgh 1995).
It could equally be well argued that these imitations of blue tit sounds have no function at all and are merely the result of learning mistakes in the sensitive period of great tits because blue and great tits form mixed foraging flocks together with other, mainly Parus species from late summer to spring (Cramp and Perrins 1993). If this would be the case, we would expect to find more illustrations of great tit imitations of blue tit song in literature because great tit singing behaviour has been the focus of numerous studies in the past. On the other hand, heterospecific learning could have evolved not because of learning mistakes but because of the frequent interactions of great and blue tits during the first year of life (Cramp and Perrins 1993). After fledging, most juveniles aggregate in multiple species flocks (Cramp and Perrins 1993) and the social interactions between blue and great tits may cause recognition and memorisation of blue tit sounds by great tits. Hence, this might be an example of perceptual learning or learning for discrimination, which is different from learning for performance (McGregor and Avery 1986), which may facilitate neighbour recognition between individuals of competing species. Because it has been shown that learning for discrimination is not restricted to early life (McGregor and Avery 1986) and there is some evidence that learning songs for performance can occur throughout life as well in the great tit (McGregor and Krebs 1989), intense competition or social stimulation might trigger a mechanism that activates performance of songs learned for discrimination. Support for this prediction comes from a study of McGregor and Krebs (1989) that showed that the composition of the great tits’ repertoire can change throughout life and that this is a common feature in great tits. Recent studies in nightingales (Luscinia megarhynchos) and white-crowned sparrows (Zonotrichia leucophrys) provide evidence of a silent storage of songs during the sensitive phase that were not used in overt song behaviour but could be activated through vocal interactions later in life (Hough II et al. 2000; Nelson 2000; Geberzahn et al. 2002; Geberzahn and Hultsch 2003).
If heterospecific matching is adaptive and not the result of learning mistakes, it should be beneficial to great tits who engage in this kind of behaviour. A possible benefit could be that no time and energy is lost in aggressive encounters with heterospecifics, which is very important during the fertile period of great tit females because of the possible threat of loss of paternity (Blakey 1994; Krokene et al. 1998; Strohbach et al. 1998; Lubjuhn et al. 1999). Of course, this implies that blue tits know that an aggressive attack will follow when they ignore matching great tits. Under this scenario, naïve and experienced blue tits will probably respond differently when matched by a great tit. Hence, instead of chasing an intruding blue tit, a great tit can match its song to scare it off without leaving his female out of sight. Although these ideas are still speculative, our results suggest a strong heterospecific influence on the acquisition and composition of the vocal signals of great tits and they provide a possible mechanism that can drive song convergence in songbirds.
If the function of heterospecific vocal matching is to threaten and scare off heterospecifics, then these vocal signals should not function in intraspecific communication and heterospecific matching should evoke different behavioural responses in naïve and non-naïve blue tits. While the first prediction is simple and straightforward to test in natural conditions (quantifying great tit response to imitations and non-imitations), the latter is far more difficult to assess in the wild. Firstly, naïve and experienced blue tits will probably respond differently to imitations and secondly, because matching behaviour occurs on a short time scale, the investigator should be able to match just that song that the blue tit is singing at the very moment. Hence, to infer the consequences of heterospecific matching on blue tit behaviour requires a large archive of great tit imitations of blue tit songs and interactive playback experiments where the investigator can match the immediate song of the blue tit. Because our playback experiment was not designed to assess blue tit behaviour accurately and was confounded by the reasons mentioned in the “Materials and methods” section (e.g. attraction of neighbouring great tits), we are at this moment not able to make inferences about the effectiveness of heterospecific matching in interspecific communication. What we can say from our playback experiment is that blue tit response varied greatly among individuals: some became quiet, others changed song rate and some apparently did not change any aspect of their singing behaviour when confronted with great tit imitations.
On the other hand, the second playback experiment revealed a clear, straightforward differential response of neighbouring great tits to imitations and non-imitations: great tits did not respond to imitations but responded vigorously to normal great tit calls broadcast from an adjacent blue tit’s territory. Ten males even left the centre of their territory and the proximity of their female’s nest box to approach the sound source with an average approach response of less than 4 m. By doing this, they took a great risk of missing the emergence of their female from the nest box which may reduce the efficiency in male mate guarding (preventing the female to copulate with another male). The fact that there was virtually no response towards wren song suggests that it was a good control. The lack of response towards the great tit’s imitations of blue tit song is consistent with the finding of Doutrelant et al. (2000) who found that artificially adding a trill to great tit song resulted in a decrease in responsiveness compared to unmodified great tit songs. Obviously, these imitations either conceal species identity or they do not represent a threat to great tits. If the first is true, then these sounds deceive the mimicking species, but because the species identity is still obvious for a trained human observer, deception seems rather unlikely. Maybe one advantage of singing blue tit song in a heterospecific conflict is that the great tit will not attract another great tit, which would complicate the situation. Nevertheless, this experiment clearly shows that great tits are able to discriminate between conspecific non-imitative and imitative sounds and the weaker response to great tit imitations of blue tit songs indicates that these imitations are not part of the great tit’s repertoire that is used for intraspecific communication. These findings suggest that imitations are purely sung for interspecific purposes, which is in support of the hypothesis of heterospecific matching. This also demonstrates that heterospecific interference/interactions may influence the communication signals and vocal behaviour of a small passerine. This study is only a first step in the survey of heterospecific communication in sympatric congeneric songbirds. More work is now needed to establish the efficiency in scaring off heterospecific intruders by matching their songs and in unravelling the possible benefits this behaviour might have for the mimicking species.
References
Baptista LF, Catchpole CK (1989) Vocal mimicry and interspecific aggression in songbirds: experiments using white-crowned sparrow imitation of song sparrow song. Behaviour 109:247–257
Baylis JR (1982) Avian vocal mimicry: its function and evolution. In: Kroodsma DE, Miller EH (eds) Acoustic communication in birds. Production, perception and design features of sounds. Academic, New York pp 51–83
Becker PH (1982) The coding of species-specific characteristics in bird sounds. In: Kroodsma DE, Miller EH (eds) Acoustic communication in birds. Production, perception and design features of sounds. Academic, New York pp 213–252
Beecher MD, Campbell SE, Burt JM, Hill CE, Nordby JC (2000) Song-type matching between neighbouring song sparrows. Anim Behav 59:21–27
Blakey JK (1994) Genetic evidence for extra-pair fertilizations in a monogamous passerine, the great Parus major. Ibis 136:457–462
Brémond JC (1968) Recherches sur la sémantique et les elements vecteurs d’information dans les signaux acoustique du rouge-gorge (Erithacus rubecula L.). Terre Vie 2:109–220
Catchpole CK (1978) Interspecific territorialism and competition Acrocephalus warblers as revealed by playback experiments in area of sympatry and allopatry. Anim Behav 26:1072–1082
Catchpole CK, Baptista LF (1988) A test of the competition hypothesis of vocal mimicry, using song sparrow imitation of white-crowned sparrow song. Behaviour 106:119–128
Catchpole CK, Slater PJB (1995) Bird song: biological themes and variations. Cambridge Univ Press, Cambridge
Cody ML (1973) Character convergence. Annu Rev Syst Ecol 4:189–211
Cramp S, Perrins CM (1993) The birds of the Western Paleartic, vol. VII. Oxford Univ Press, Oxford
Dabelsteen T, McGregor PK, Shepherd M, Whittaker X, Pedersen SB (1996) Is the signal value of overlapping different from that of alternating during matched singing in Great Tits? J Avian Biol 27:189–194
Dabelsteen T, McGregor PK, Holland J, Tobias JA, Pedersen SB (1997) The signal function of overlapping singing in male robins. Anim Behav 53:249–256
Dobkin DS (1979) Functional and evolutionary relationships of vocal copying phenomena in birds. Z Tierpsychol 50: 348–363
Dhondt AA (1977) Interspecific competition between great and blue tit. Nature 268:521–523
Dhondt AA, Eyckerman R (1980) Competition and the regulation of numbers in great and blue tit. Ardea 68:121–132
Doutrelant C, Lambrechts MM (2001) Macrogeographic variation in song—a test of competition and habitat effects in blue tits. Ethology 107:533–544
Doutrelant C, Aubin T, Hitier S, Lambrechts MM (1998) Two distinct song populations of blue tits, Parus caeruleus, in the French Mediterranean. Bioacoustics 9:1–16
Doutrelant C, Leitao A, Otter K, Lambrechts MM (2000) Effect of blue tit song syntax on great tit territorial responsiveness—an experimental test of the character shift hypothesis. Behav Ecol Sociobiol 48:119–124
Falls JB, Krebs JR, McGregor PK (1982) Song matching in the great tit (Parus major): the effect of similarity and familiarity. Anim Behav 30:997–1009
Ferguson JWH, van Zyl A, Delport K (2002) Vocal mimicry in African ossypha robin chats. J Ornithol 139:319–330
Geberzahn N, Hultsch H (2003) Long-time storage of song types in birds: evidence from interactive playbacks. Proc R Soc Lond B 270:1085–1090
Geberzahn N, Hultsch H, Todt D (2002) Latent song type memories are accessible through auditory stimulation in a hand-reared songbird. Anim Behav 64:783–790
Gorissen M (2002) Onderzoek naar de akoestische interacties tussen pimpelmezen en koolmezen. Master’s Thesis, University of Antwerp, Antwerp
Gorissen L (2005) The vocal communication strategies of blue and great tits, studied from an intra- and interspecific perspective. Doctoral Dissertation, University of Antwerp, Antwerp
Gorissen L, Eens M (2004) Interactive communication between male and female great tits (Parus major) during the dawn chorus. Auk 121:184–191
Haavie J, Borge T, Bures S, Garamszegi LZ, Lampe HM, Moreno J, Qvarnström A, Török J, Saetre G-P (2004) Flycatcher song in allopatry and sympatry—convergence, divergence and reinforcement. J Evol Biol 17:227–237
Hailman JP (1977) Optical signals. Indiana Univ Press, Bloomington
Hough GE II, Nelson DA, Volman SF (2000) Re-expression of songs deleted during vocal development in white-crowned sparrows, Zonotrichia leucophrys. Anim Behav 60:279–287
Jones AE, Ten Cate C, Bijleveld CCJH (2001) The interobserver reliability of scoring sonograms by eye: a study on methods, illustrated on zebra finch songs. Anim Behav 62:791–801
Kempenaers B, Dhondt AA (1991) Competition between blue and great tit for roosting sites in winter: an aviary experiment. Ornis Scand 22:73–75
Kostan KM (2002) The evolution of mutualistic interspecific communication: assessment and management across species. J Comp Psychol 116:206–209
Krebs JR, Avery M, Cowie R (1981a) Effect of mate removal on singing behaviour of great tits. Anim Behav 29:635–637
Krebs JR, Ashcroft R, van Orsdol K (1981b) Song matching in the great tit (Parus major L.). Anim Behav 29:918–923
Krokene C, Rigstad K, Dale M, Lifjeld JT (1998) The function of extrapair paternity in blue tits and great tits: good genes or fertility assurance? Behav Ecol 9:649–656
Kroodsma DE, Byers, BE, Goodale E, Johnson S, Liu W-C (2001) Pseudoreplication in playback experiments, revisited a decade later. Anim Behav 61:1029–1033
Lambrechts MM (1996) Organisation of birdsong and constrains on performance. In: Kroodsma DE, Miller EH (eds) Ecology and evolution of acoustic communication in birds. Cornell University Press, Ithaca pp 305-320
Lubjuhn T, Strohbach S, Brün J, Gerken T, Epplen JT (1999) Extra-pair paternity in great tits (Parus major)—a long term study. Behaviour 136:1157–1172
Martin PR, Martin TE (2001) Behavioral interactions between coexisting species: song playback experiments with wood warblers. Ecology 82:207–218
Martin PR, Fotheringham JR, Ratcliffe L, Robertson RJ (1996) Response of American redstarts (Suborder Passeri) and least flycatcher (Suborder Tyrannii) to heterospecific playback: the role of song in aggressive interactions and interference competition. Behav Ecol Sociobiol 39:227–235
McGregor PK, Avery MI (1986) The unsung songs of great tits (Parus major): learning neighbours’ songs for discrimination. Behav Ecol Sociobiol 18:311–316
McGregor PK, Krebs JR (1989) Song learning in adult great tits tits (Parus major): effects of neighbours. Behaviour 108:139–159
McGregor PK, Dabelsteen T, Shepherd M, Pedersen S (1992) The signal value of matched singing in great tits: evidence from interactive playback experiments. Anim Behav 43:987–998
Minot EO (1981) Effects of interspecific competition for food in breeding blue and great tits. J Anim Ecol 50:375–385
Molles LE, Vehrencamp SV (2001) Neighbour recognition by resident males in the banded wren, Thryothorus pleurostictus, a tropical songbird with high song type sharing. Anim Behav 61:119–127
Morton ES (1976) Vocal mimicry in the thick-billed Euphonia. Willson Bull 88:485–487
Nelson DA (2000) Song overproduction, selective attrition and song dialects in the white-crowned sparrow. Anim Behav 60:887–898
Nowicki S, Westneat MW, Hoese WJ (1992) Birdsong: motor function and the evolution of communication. Semin neurosci 4:385–390
Osiejuk TS, Kuczynski L (2000) Mixed and atypical singers among treecreepers Certhia brachydactyla and C. familiaris: a review and preliminary data from western Poland. Biol Bull Pozn 37:83–94
Osiejuk TS, Kuczynski L (2003) Response to typical, mixed and shortened song versions in Eurasian treecreepers, Certhia familiaris. Biologia Bratislava 58:985–989
Owen-Ashley NT, Schoech SJ, Mumme RL (2002) Context-specific response of Florida scrub-jay pairs to northern mockingbird vocal mimicry. Condor 104:858–865
Perrins CM (1979) British Tits. Collins, London
Poesel A, Dabelsteen T, Pedersen SB (2004) Dawn song of male blue tits as a predictor of competitiveness in midmorning singing interactions. Acta Ethol 6:65–71
Reed TM (1982) Interspecific territoriality in the chaffinch and great tit on islands and the mainland of Scotland: playback and removal experiments. Anim Behav 30:171–181
Ryan MJ, Brenowitz EA (1985) The role of body size, phylogeny, and ambient noise in the evolution of bird song. Am Nat 126:87–100
Robinson SK, Terborgh J (1995) Interspecific aggression and habitat selection by Amazonian birds. J Anim Ecol 64:1–11
Schottler B (1995) Songs of blue tits Parus caeruleus palmensis from La Palma (Canary Islands)—a test of hypotheses. Bioacoustics 6:135–152
Secondi J, Bretagnolle V, Compagnon C, Faivre B (2003) Species-specific song convergence in a moving hybrid zone between two passerines. Biol J Linn Soc 80:507–517
Slikas B, Sheldon FH, Gill FB (1996) Phylogeny of titmice (Paridae): estimate of relationships among subgenera based on DNA–DNA hybridization. J Avian Biol 27:70–82
Sorjonen J (1986) Mixed singing and interspecific territoriality—consequences of secondary contact of two ecologically and morphologically similar nightingale species in Europe. Ornis Scand 17:53–67
Strohbach S, Curio E, Bathen A, Epplen JT, Lubjuhn T (1998) Extra-pair paternity in the great tit (Parus major): a test of the good genes hypothesis. Behav Ecol 9:388–396
Todt D, Naguib M (2000) Vocal interactions in birds: the use of song as a model in communication. In: Slater PJB, Rosenblatt JS, Snowdon CT, Roper TJ (eds) Advances in the study of behavior, vol 29. Academic, San Diego
Törok J, Toth L (1999) Asymmetric competition between two tit species: a reciprocal removal experiment. J Anim Ecol 68:338–345
Acknowledgements
We are grateful to R. Oblonsek for the help during fieldwork and the time consuming scoring of sonograms, P. Scheys for help during fieldwork and T. Snoeijs for statistical advice. L. Gorissen was funded by a Ph.D. grant of the Institute for the Promotion of Innovation through Science and Technology in Flanders (IWT Vlaanderen). All experiments performed comply with the current laws of the Belgian Government.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by T. Czeschlik
Rights and permissions
About this article
Cite this article
Gorissen, L., Gorissen, M. & Eens, M. Heterospecific song matching in two closely related songbirds (Parus major and P. caeruleus): great tits match blue tits but not vice versa. Behav Ecol Sociobiol 60, 260–269 (2006). https://doi.org/10.1007/s00265-006-0164-6
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00265-006-0164-6