
Abstract
Myocardial energy and lipid homeostasis is crucial for normal cardiac structure and function. Either shortage of energy or excessive lipid accumulation in the heart leads to cardiac disorders. Peroxisome proliferator-activated receptors (PPARα, -β/δ and -γ), members of the nuclear receptor transcription factor superfamily, play important roles in regulating lipid metabolic genes. All three PPAR subtypes are expressed in cardiomyocytes. PPARα has been shown to control transcriptional expression of key enzymes that are involved in fatty acid (FA) uptake and oxidation, triglyceride synthesis, mitochondrial respiration uncoupling, and glucose metabolism. Similarly, PPARβ/δ is a transcriptional regulator of FA uptake and oxidation, mitochondrial respiration uncoupling, and glucose metabolism. On the other hand, the role of PPARγ on transcriptional regulation of FA metabolism in the heart remains obscure. Therefore, both PPARα and PPARβ/δ are important transcriptional regulators of myocardial energy and lipid homeostasis. Moreover, it appears that the heart needs to have two PPAR subtypes with seemingly overlapping functions in maintaining myocardial lipid and energy homeostasis. Further studies on the potential distinctive roles of each PPAR subtype in the heart should provide new therapeutic targets for treating heart disease.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Fatty acid (FA) metabolism provides a large fraction of the amaranthine energy needs to support normal cardiac function. Perturbation in myocardial energy and lipid homeostasis is a common feature for many cardiac disorders. One of the key determinants of myocardial energy and lipid homeostasis is a transcriptional network that governs relative fluxes of energy substrates through affecting expression levels of key proteins in various metabolic pathways. Emerging evidence shows that altering expression of genes directing FA metabolism in the heart is associated with substrate switches in pathological conditions. Peroxisome proliferator-activated receptors (PPARα, -β/δ, and -γ), members of the ligand-activated nuclear receptor superfamily, are the key transcriptional regulators in FA metabolism. Each PPAR subtype functions as an obligate heterodimer with the retinoid X receptor (RXR). The PPAR and RXR heterodimer binds to the PPAR responsive element (PPRE) on its target genes and activates them [1]. FAs and/or lipid metabolites serve as endogenous ligands of PPARs to mediate adaptive metabolic responses to changes in systemic energy supplies [1]. A growing number of natural ligands have been identified, such as leukotriene B4 and 8-(S)-hydroxyeicosatetraenoic acid and 15-deoxy-Δ12, 14-prostaglandin J2 (15d-PGJ2) [3, 4]. At present, no high affinity natural ligand has been identified for any of the PPARs. Therefore, a physiological role of the receptor may be to sense the total flux of Free FA (FFA) in important tissues, such as the heart. The lipid lowering drug fibrates and the antidiabetic Thiazolidinediones (TZDs) have been identified as PPARα and PPARγ ligands, respectively. In addition, synthetic compounds have been developed for the selective activation of each or multiple PPAR subtypes.
This review focuses on discussing the essential roles of PPARs in the heart as the key transcriptional determinants of myocardial energy and lipid homeostasis.
Myocardial energy and lipid homeostasis determines cardiac structure and function
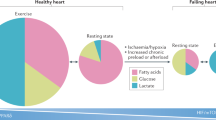
The energy and lipid homeostasis in cardiomyocytes relies on the coordinating regulation of lipid uptake, oxidation, triglyceride (TG) synthesis, and lipolysis, as well as lipoprotein secretion (Fig. 1). Excessive FA uptake, depressed FA oxidation (FAO), or reduced lipid secretion potentially contribute to lipid accumulation in cardiomyocytes. FA uptake and FAO are the key steps that determine the myocardial energetic supplies. In physiological state, FA and carbohydrates such as glucose serve as energy substrates for postnatal mammalian cardiac muscle, with 60–70% from FAO [5–7]. Optimal cardiac function depends on a precise coupling of energy supply and expenditure as well as a delicate balance in energy flux between FAs and glucose. The lipid and energy homeostasis of the heart is tightly controlled by various mechanisms (see review [8]). Abnormality of energy metabolism is one of the most common pathological phenomena in many cardiac disorders. Inherited defects in many key enzymes on the mitochondrial FAO pathway are associated with cardiomyopathy and sudden death in children and young adults [9]. Acquired cardiac disorders such as myocardial ischemia/reperfusion and cardiac hypertrophy are also associated with alteration of energy homeostasis [10, 11]. Myocardial metabolism switches from the utilization of FA to the utilization of glucose during the development of cardiac hypertrophy and heart failure [10, 12–15]. This metabolic switch may initially be adaptive. However, accumulation of intracellular FA in these acquired conditions may contribute to contractile dysfunction [16]. Subsequent myocardial lipid accumulation results in cardiomyocyte apoptosis and congestive heart failure [17–19]. Similarly, animal models of diabetes exhibit myocardial lipid metabolism disorder, contributing to the pathogenesis of diabetic cardiomyopathy [17, 20–22]. It remains unclear whether the cardiomyopathies developed in these pathologic states result from shortage of energy or toxic lipid accumulation. Recent studies have shed light on how transcriptional regulation controls gene expression of mitochondrial and peroxisomal lipid metabolism in the heart. It is evident that PPARs are the key transcriptional determinants of myocardial energy and lipid homeostasis. Ligands that specifically activate PPAR subtype(s) to regulate transcriptional activities of lipid metabolic genes logically become appealing drugs for treating perturbed myocardial energy and lipid homeostasis in various cardiac pathological conditions. Therefore, in-depth studies on the expression patterns and the potentially distinctive functions of each PPAR subtype in the heart become indispensable.

Schematic illustration of major aspects of lipid and energy metabolism in cardiomyocytes. Fatty acids are the main sources of substrates used by cardiomyocytes to generate energy. Fatty acid metabolism in cardiomyocytes involves in multiple aspects, such as free fatty acid (FFA) uptake, β-oxidation, energy uncoupling, triglyceride (TG) synthesis, and lipoprotein secretion. LDL, Low-density lipoproteins; VLDL, very low density lipoprotein; FATP, fatty acid binding protein; LACS, long-chain acyl-CoA synthetase; FAT/CD36, fatty acid translocase; CPT-I, carnitine palmitoyltransferase-I; CPT-II, carnitine palmitoyltransferase-II; CACT, carnitine-acylcarnitine translocase; LCAD, long-chain acyl-CoA dehydrogenase; HAD, 3-hydroxyacyl CoA dehydrogenase; Thiolase, 3-oxoacyl-CoA thiolase; UCP2, uncoupling protein 2; UCP3, uncoupling protein 3
Myocardial expression profile of PPARs
It has been well recognized that PPARα is abundantly expressed in cardiomyocytes [23, 24]. Whereas it is recognized that PPARβ/δ has a ubiquitous expression pattern, its expression is predominant in cardiomyocytes of the rodent heart [24]. It is controversial on the cardiac expression of PPARγ. It has been reported that PPARγ transcript is very low to undetectable in ribonucleic acid (RNA) samples extracted from cultured neonatal and adult cardiomyocytes and heart tissues [25]. Nevertheless, Northern blot [26] and real-time reverse-transcriptase polymerase chain reaction [27] both reveal the transcript expression of PPARγ on RNA samples of human heart tissues. In addition, RNA-protecting assay can also detect the PPARγ transcript in RNA samples from heart tissues of pigs [28]. Immunoblotting experiments demonstrate the expression of PPARγ in cardiomyocytes, which reaches about 30% of the abundance observed in adipocytes [29]. Expression of PPARγ protein in samples from neonatal and adult rat cardiomyocytes and from neonatal and adult heart ventricles can be detected by Western blots [25]. Immunostainings on heart sections from humans also support that PPARγ protein expresses within cardiomyocytes [27]. Recently, PPARγ protein expression was also demonstrated by Western blots on protein samples of mouse heart tissues [30]. More importantly, cardiomyocyte-restricted PPARγ knockout in mice leads to cardiac hypertrophy with increased nuclear factor-κB activities [30]. Therefore, it becomes clear that PPARγ is indeed expressed in cardiomyocytes from various species of animals. However, it is not clear whether each individual subtype of PPARs exerts distinctive biological effects in the heart.
Roles of PPARα in myocardial energy and lipid homeostasis
Transcriptional regulation of FAO genes by PPARs in the heart has been the main topic of studies as FAO supplies a majority of the needed energy for the heart to work as a pump. Studies on the in vitro effect of PPARα ligands such as Wy14643 or on the conventional PPARα knockout mice have unraveled how PPARα regulates FA metabolic genes. More recently, studies on heart-specific PPARα overexpression transgenic lines confirmed many of the previous findings on potential target genes of PPARα in the heart [31]. Studies using PPARα-selective synthetic ligands such as Wy14643 on cultured cardiomyocytes provide additional information on potential target genes of PPARα in cardiomyocytes [24, 25, 32].
PPARα regulates FA uptake genes
As shown in Table 1, PPARα regulates the transcript expression of key enzymes that are important components of FA uptake. These FA uptake genes include FA translocase (FAT/CD36) [23, 31] and FA transport protein 1 (FATP1) [23, 31]. FAT/CD36 has been proposed to be the predominant mechanism to facilitate FA uptake by myocytes [33]. Heart-specific transgenic overexpression of FATP1 leads to cardiac dysfunction with the increased FFA uptake by the heart [34]. Wy14643 administration significantly induces the cardiac expression of genes encoding proteins involved in cellular FA import and thioesterification, such as FATP1, CD36, and long-chain fatty acyl-CoA synthetase (LACS), in PPARα-overexpressed hearts, but not in nontransgenic controls [31]. Consistent findings have also been reported in PPARα null hearts [23]. PPARα-selective ligand, such as Wy14643, also induces LACS expression in cultured cardiomyocytes [24, 25]. Therefore, it is clear that PPARα in cardiomyocytes plays a crucial role in governing the transcript expression of FA uptake genes.
PPARα regulates FAO genes in cardiomyocytes
PPARα is abundantly expressed in tissues that require high rates of FAO and mediates lipid-induced activation of FAO genes based on studies of a conventional PPARα knockout mice [35]. The metabolic phenotype of PPARα null mice is associated with failure of the liver and the heart to induce β-oxidative pathways in response to physiological or pharmacological perturbations in lipid metabolism [36–38]. Promoter studies showed that PPARα regulates transcriptional expression of medium-chain acyl-CoA dehydrogenase (MCAD) and carnitine palmitoyltransferase I (CPT-I) [39]. Further studies on PPARα knockout hearts and on cultured cardiomyocytes treated with Wy14643 demonstrated that cardiac transcriptional expression of both mitochondria and peroxisome FAO genes are regulated by PPARα (Table 1). These mitochondrial FAO enzymes include muscle CPT-I [24, 25, 31, 32, 36], liver CPT-I [24, 37], long-chain acyl-CoA dehydrogenase (LCAD) [24, 25, 36], MCAD [24, 36], 3-hydroxyacyl CoA dehydrogenase [2], 3-oxoacyl-CoA (thiolase) [24, 32], and mitochondrial thioesterase 1 [40]. The peroxisomal-specific FAO enzymes include acyl-CoA oxidase (ACO) [31, 32] and the very long chain acyl-CoA dehydrogenase (VLCAD) [23, 32] and are also regulated by PPARα. Pyruvate dehydrogenase kinase 4 (PDK4), an enzyme suppressing glucose oxidation via inhibiting pyruvate dehydrogenase complex activity, is shown to be upregulated by PPARα in the heart [24, 31, 32] (Table 1). Mitochondrial respiration uncoupling proteins such as uncoupling protein 2 and 3 [24, 25, 31] are upregulated by PPARα (Table 1). As a result, PPARα null hearts consequently show reduced FAO rates and increased glucose oxidation rates [23]. Reduced expression of malonyl-CoA decarboxylase (MCD) in the PPARα null heart also contributes to the higher concentrations of malonyl-CoA, thus lowering FAO rates [41].
PPARα regulates TG synthetic enzyme genes in the heart
Little is known about TG synthesis in cardiomyocytes. Many important enzymes that are involved in TG synthesis are expressed in cardiomyocytes. TG synthesis represents an active aspect of lipid metabolism in the heart. Heart-specific PPARα overexpression upregulates glycerol-3-phosphate acyltransferase (GPAT) and diacylglycerolacyltransferase, two key enzymes involved in the esterification of FAs to TG at base line and further with fasting [31] (Table 1). In addition, 1-acyl-sn-glycerol 3-phosphate acyltransferase (AGPAT)-3 (or lysophosphatidic acid acyltransferase; Table 1) is also regulated by PPARα activation [42]. AGPAT catalyses the acylation of lysophosphatidic acid to form phosphatidic acid [43], the precursor of all glycerolipids. The excessive TG in cardiomyocytes forms lipid droplets that are usually surrounded by phospholipids monolayer [44]. Cardiomyocytes contain lipid droplets in various sizes depending on the disease or dietary conditions. These lipid droplets contain a class of proteins in their surface layers that share a homologous sequence. A member of this class of protein named MLDP (myocardial lipid droplet protein) was recently identified. MLDP expression is upregulated in wild-type hearts by Wy14643, but is blocked in PPARα null hearts [45]. Therefore, the TG synthesis pathway within the cardiomyocytes is likely regulated by PPARα. The activation of this pathway in response to increased intracellular FFA should help prevent the deteriorating effects of certain FAs. However, more detailed studies are needed to uncover the biological significance of this regulation.
PPARα regulates FA secretion in cardiomyocytes?
Because lipid accumulation is toxic to the heart, the heart may have a capacity to increase its export of TG in states with reduced FAO and increased FA uptake. The heart can synthesize and secrete lipoproteins [46]. Apo B-100 and its edited product apoB-48 are the major structural apolipoprotein of liver-derived very low density lipoproteins and low-density lipoproteins (LDL) [46–48]. Because the apoB mRNA is not edited in the heart, the full-length apoB100 protein secretes lipoproteins in the heart [46, 49]. The heart also expresses microsomal TG transfer protein (MTP), which is a protein adding lipid to apoB. MTP transfers TG onto the apoB protein structure before transferring into the endoplasmic reticulum. When intracellular FFA or TG is increased, cardiomyocytes secrete apoB-containing lipoproteins to prevent lipid accumulation. Cardiac apoB expression leads to reduced myocardial TG content in mice with type II diabetes [50], LCAD deficiency [51], and heart-specific lipoprotein lipase overexpression [52]. Recently, it has been shown that the activation of PPARα increases the expression and activity of MTP in the liver through the PPRE on the MTP promoter [53]. Although it is feasible to predict that MTP expression should also be regulated by PPARα, it remains unclear if the PPARα-activated expression of MTP is subtype specific or tissue dependent.
Cardiac PPARα expression in pathological cardiac hypertrophy and in aging
Genes encoding FAO enzymes are downregulated in concomitant with the switch from FA to glucose utilization in cardiac hypertrophy and heart failure [15, 54–56]. Interestingly, PPARα is downregulated during the development of cardiac hypertrophy in animal models of ventricular pressure overload [57–59], Thus, it was proposed that the downregulation of PPARα is a key determinant of the energy switch in cardiac hypertrophy and heart failure [60]. Similarly, the capacity of myocardial energy production from FA metabolism is also depressed in the aging heart accompanied by the downregulation of PPARα expression [2]. More importantly, exercise training corrects the downward trend of PPARα expression and activity in concomitant with enhanced FAO in aged rat hearts [2]. However, while reversing the downregulation of PPARα target genes in the hypertrophied heart and preventing substrate switching, reactivation of PPARα exerts detrimental effects on cardiac performance [59]. Therefore, PPARα downregulation may be essential to maintain contractile function of the hypertrophied heart. Furthermore, variation in the PPARα gene influences left ventricular growth in response to exercise and hypertension in humans, implicating that maladaptive cardiac substrate utilization can play a causative role in the pathogenesis of left ventricular hypertrophy. PPARα may serve as a regulator of left ventricle growth in response to an intense short-term physiological stimulus [61]. It has been shown that ligands of PPARα attenuate necrosis in acute myocardial infarction [62] and blunt the development of endothelin-induced cardiac hypertrophy [63, 64]. However, the expression and activity of PPARα are not necessarily always concomitant with a reduced rate of FAO in disease states. In the left ventricular tissues from pacing-induced failing dog hearts, the activity of CPT-I and MCAD decreased, but the expression of PPARα is unchanged [65]. Therefore, differential interaction of PPARα with other factors may also play roles in the reduction in FAO genes in the failing hearts.
In summary, PPARα is a key determinant of myocardial lipid and energy homeostasis by regulating transcriptional expression of key components of FA metabolism of the cardiomyocytes. These key components include almost all aspects of lipid metabolism in a cardiomyocyte, such as lipid uptake, FAO, TG synthesis mitochondrial respiration uncoupling, and glucose metabolism (Fig. 2). More studies are warranted to confirm the functional aspects of PPARα-directed transcriptional activities in determining myocardial energy and lipid homeostasis.

PPARα determines myocardial lipid and energy homeostasis via transcriptional regulation of lipid and energy metabolisms in cardiomyocytes. PPARα governs transcriptional expression of key enzymes that are involved in fatty acid uptake and oxidation, TG synthesis, mitochondrial respiration uncoupling, and glucose metabolism
Roles of PPARβ/δ in myocardial energy and lipid homeostasis
In contrast to the well-characterized roles of PPARα in regulating lipid metabolism in the heart, less is known about the roles of PPARβ/δ in the heart. It has been suspected that it exerts similar action on the heart as those of PPARα. PPARβ/δ is predominantly expressed in cardiomyocytes, but not in other cell types in the myocardium [24]. As synthetic PPARβ/δ-selective ligands have become available, evidence is emerging that PPARβ/δ regulates similar FAO transcripts in cardiomyocytes as PPARα [24, 25].
PPARβ/δ regulates FA uptake genes in cardiomyocytes
LACS has been shown to be upregulated by PPARβ/δ ligand treatment in cultured cardiomyocytes [24, 25]. As LACS plays essential roles in coordinating FA uptake, PPARβ/δ should be a key determinant of FA uptake. Additionally, it would not be a surprise to see that PPARβ/δ also regulates the transcriptional expression of other lipid uptake genes in cardiomyocytes. This is especially true for FATP1 and FAT/CD36, which have been shown to be regulated by PPARα (Table 1).
PPARβ/δ regulates FAO genes in cardiomyocytes
PPARβ/δ-selective ligand treatments and PPARβ/δ overexpression in cultured cardiomyocytes result in elevation of FAO genes and FAO rates in a classic ligand binding dependent mechanism [24]. Both mitochondria-specific (M-CPT-I, L-CPT-I, UCP2, UCP3, PDK4, thiolase, MCAD, LCAD, and MCD) and peroxisome-specific (ACO, VLCAD, and thiolase) FAO genes are regulated by PPARβ/δ in the heart (Table 1). More importantly, a definite and essential role of PPARβ/δ in maintaining constitutive myocardial FAO has been revealed recently in studies performed in a cardiomyocyte-restricted PPARβ/δ knockout mouse model [32]. In addition to depressed FAO, these mice develop severe phenotypic changes, such as cardiac dysfunction, myocardial lipid accumulation, and progressive heart failure. The dominant expression of PPARβ/δ in cardiomyocytes of the heart explains, at least in part, why mice with cardiomyocyte-restricted PPARβ/δ knockout exhibit remarkable phenotypic changes [24, 32]. Thus, PPARβ/δ may play a key role as a “sensor” of intracellular FA content and a constitutive determinant of high-level FA metabolism observed in normal adult hearts. Although PPARα and PPARβ/δ regulate similar set of FAO genes in cardiomyocytes, they are not interdependent on each other. Deletion of one from cardiomyocytes does not affect the effects of another on activating FAO gene expression [24, 32]. Although PPARβ/δ plays overlapping roles as PPARα does on activating myocardial FAO, whether PPARβ/δ also involves transcriptional regulation of other lipid metabolic pathways in the heart remains relatively unclear. Interestingly, cardiomyocyte-restricted PPARβ/δ knockout but not the conventional PPARα knockout mice exhibit myocardial neutral lipid accumulation at baseline condition [23, 32]. Therefore, a mismatch of PPARβ/δ-activated FAO and lipid uptake must be existed. For example, it is likely that depressed myocardial FAO in PPARβ/δ deficient heart could not oxidize the remaining FAs. Nevertheless, it is also possible that PPARβ/δ and PPARα deficient hearts employ, respectively, different regulating mechanisms in response to the different cell contexts: PPARβ/δ is only missing from the cardiomyocytes in the cardiomyocyte-restricted PPARβ/δ knockout mice, whereas PPARα is eliminated from all cell types and all tissues in the conventional PPARα null mice.
In summary, PPARβ/δ is emerging as an essential transcription factor in regulating myocardial lipid and energy homeostasis. It is especially clear that PPARβ/δ regulates major myocardial metabolic genes to activate FAO, increase mitochondrial respiration uncoupling, and suppress glucose oxidation (Fig. 3). It remains unknown whether PPARβ/δ also governs transcription of genes encoding other aspects of myocardial lipid metabolism. The regulation of PPARβ/δ itself in response to dietary and other pathological stimuli is also obscure. It has been recently reported that PPARβ/δ-selective ligand or overexpression of PPARβ/δ in cultured cardiomyocytes suppresses lipopolysaccharide-induced inflammatory responses [66, 67]. Furthermore, activation of PPARβ/δ can inhibit hypertrophic responses in cardiomyocytes [66]. However, there is no direct evidence that the inhibitory effects of PPARβ/δ to inflammation and hypertrophy are associated with its action on FAO gene regulation. Further studies are needed to unravel the important roles of PPARβ/δ on lipid metabolic regulation.

PPARβ/δ determines myocardial lipid and energy homeostasis via transcriptional regulation of lipid and energy metabolisms in cardiomyocytes. PPARβ/δ governs transcriptional expression of key enzymes that are involved in fatty acid uptake and oxidation, mitochondrial respiration uncoupling and glucose metabolism
The mystery role of PPARγ in myocardial energy homeostasis
A primary role of PPARγ in the heart remains elusive. Given the crucial roles of PPARγ on lipogenesis and glucose metabolism in many tissues [68], including the liver [69] and skeletal muscle [70, 71], which express very low levels of PPARγ [72], it is conceivable that even relatively low levels of cardiac PPARγ may play important roles in cardiomyocytes.
Accumulating evidence indicates that PPARγ activators, such as rosiglitazone, troglitazone, and 15d-PGJ2 can suppress hypertrophic response in cultured cardiomyocytes [73–75] and in animal models [76]. In addition, it has been shown by many recent reports that activation of PPARγ with TZD drugs protects the myocardium from ischemic/reperfusion injury [62, 77, 78]. Most recently, Duan et al. showed that cardiomyocyte-specific PPARγ knockout induces cardiac hypertrophy with elevated nuclear factor-κB activities [30]. Treatments with selective PPARγ ligand rosiglitazone in mice also lead to cardiac hypertrophy [30], probably by increased water retention [79, 80]. Interestingly, studies on the above cardiomyocyte-restricted PPARγ knockout mice did not reveal any change on the transcript abundances of key lipid metabolic enzymes [30], thus ruling out PPARγ as a primary effector on suppressing cardiac hypertrophy via regulating FAO. These seemingly contradictory results may be derived from many factors. For example, the beneficial effect of TZD drugs on the heart may be a combination of direct and indirect effects on insulin sensitivity and on inflammatory responses via systemic and/or local PPARγ activation. As a result, TZD drugs may only be effective in pathological conditions of the heart, which usually exhibits exacerbated inflammation and disturbing lipid metabolism [81, 82]. On the other hand, the activation of TZD could lead to increased water retention, resulting in volume-overload, and hence, cardiac hypertrophy. The beneficial and harmful effects of TZD drugs to the heart may depend on various drug actions on different tissues. More in-depth studies will be needed to address the potential tissue selective effects of PPARγ activation. Although these data suggest the involvement of PPARγ in a pathway for negative regulation of cardiac hypertrophy, the roles of cardiac PPARγ on myocardial lipid homeostasis remain unclear.
Future perspectives and conclusion
Although considerable progress has been made in the understanding of the roles of PPARs in myocardial lipid homeostasis, there are many open questions regarding the roles of these important nuclear receptors in the heart. It appears that PPARα and PPARβ/δ in cardiomyocytes share many overlapping functional roles in regulating lipid homeostasis; however, their differential roles remain obscure. Further studies on their potential differential regulation in the heart in response to various conditions, such as dietary stresses, are warranted. Moreover, whether PPARγ plays any roles in regulating myocardial lipid homeostasis remains an open question. Studies on mice with temporally inducible heart-specific PPARs inactivation and/or inducible cardiomyocyte-restricted transgenic overexpression of each PPAR subtypes should provide important clues to answer the above questions. Furthermore, studies on mice with double or triple knockout of two or three of the PPAR subtypes should help to identify potential intersecting of each PPAR in cardiomyocytes. Only by understanding how these multiple PPARs intersect will we be able to exploit the therapeutic potential of PPAR ligands for treating lipid metabolic disorders that underlie in cardiomyopathy. An understanding of the molecular regulatory mechanisms involved in maintaining cardiac lipid and energy homeostasis under various cardiac pathological conditions should provide novel insights into the therapeutic developments of inherited and acquired diseases of the cardiovascular system in humans.
References
Kliewer SA, Xu HE, Lambert MH, Willson TM (2001) Peroxisome proliferator-activated receptors: from genes to physiology. Recent Prog Horm Res 56:239–263
Iemitsu M, Miyauchi T, Maeda S, Tanabe T, Takanashi M, Irukayama-Tomobe Y, Sakai S, Ohmori H, Matsuda M, Yamaguchi I (2002) Aging-induced decrease in the PPAR-alpha level in hearts is improved by exercise training. Am J Physiol Heart Circ Physiol 283:H1750–H1760
Devchand PR, Keller H, Peters JM, Vazquez M, Gonzalez FJ, Wahli W (1996) The PPARalpha-leukotriene B4 pathway to inflammation control. Nature 384:39–43
Kliewer SA, Lenhard JM, Willson TM, Patel I, Morris DC, Lehmann JM (1995) A prostaglandin J2 metabolite binds peroxisome proliferator-activated receptor gamma and promotes adipocyte differentiation. Cell 83:813–819
van der Vusse GJ, Glatz JF, Stam HC, Reneman RS (1992) Fatty acid homeostasis in the normoxic and ischemic heart. Physiol Rev 72:881–940
Vary TC, Reibel DK, Neely JR (1981) Control of energy metabolism of heart muscle. Annu Rev Physiol 43:419–430
Neely JR, Rovetto MJ, Oram JF (1972) Myocardial utilization of carbohydrate and lipids. Prog Cardiovasc Dis 15:289–329
Yang Q, Cheng LH (2005) Molecular regulation of lipotoxicity in the heart. Drug Discov Today 2:101–107
Kelly DP, Strauss AW (1994) Inherited cardiomyopathies. N Engl J Med 330:913–919
Wittels B, Spann JF Jr (1968) Defective lipid metabolism in the failing heart. J Clin Invest 47:1787–1794
Massie BM, Schaefer S, Garcia J, McKirnan MD, Schwartz GG, Wisneski JA, Weiner MW, White FC (1995) Myocardial high-energy phosphate and substrate metabolism in swine with moderate left ventricular hypertrophy. Circulation 91:1814–1823
Bishop SP, Altschuld RA (1970) Increased glycolytic metabolism in cardiac hypertrophy and congestive failure. Am J Physiol 218:153–159
Christe ME, Rodgers RL (1994) Altered glucose and fatty acid oxidation in hearts of the spontaneously hypertensive rat. J Mol Cell Cardiol 26:1371–1375
Takeyama D, Kagaya Y, Yamane Y, Shiba N, Chida M, Takahashi T, Ido T, Ishide N, Takishima T (1995) Effects of chronic right ventricular pressure overload on myocardial glucose and free fatty acid metabolism in the conscious rat. Cardiovasc Res 29:763–767
Taegtmeyer H, Overturf ML (1988) Effects of moderate hypertension on cardiac function and metabolism in the rabbit. Hypertension 11:416–426
Corr PB, Creer MH, Yamada KA, Saffitz JE, Sobel BE (1989) Prophylaxis of early ventricular fibrillation by inhibition of acylcarnitine accumulation. J Clin Invest 83:927–936
Zhou YT, Grayburn P, Karim A, Shimabukuro M, Higa M, Baetens D, Orci L, Unger RH (2000) Lipotoxic heart disease in obese rats: implications for human obesity. Proc Natl Acad Sci USA 97:1784–1789
Regan TJ, Weisse AB (1992) Diabetic cardiomyopathy. J Am Coll Cardiol 19:1165–1166
Zoneraich S, Mollura JL (1993) Diabetes and the heart: state of the art in the 1990s. Can J Cardiol 9:293–299
Kenno KA, Severson DL (1985) Lipolysis in isolated myocardial cells from diabetic rat hearts. Am J Physiol 249:H1024–H1030
Denton RM, Randle PJ (1967) Measurement of flow of carbon atoms from glucose and glycogen glucose to glyceride glycerol and glycerol in rat heart and epididymal adipose tissue. Effects of insulin, adrenaline and alloxan-diabetes. Biochem J 104:423–434
Paulson DJ, Crass MF 3rd (1982) Endogenous triacylglycerol metabolism in diabetic heart. Am J Physiol 242:H1084–H1094
Watanabe K, Fujii H, Takahashi T, Kodama M, Aizawa Y, Ohta Y, Ono T, Hasegawa G, Naito M, Nakajima T, Kamijo Y, Gonzalez FJ, Aoyama T (2000) Constitutive regulation of cardiac fatty acid metabolism through peroxisome proliferator-activated receptor alpha associated with age-dependent cardiac toxicity. J Biol Chem 275:22293–22299
Cheng L, Ding G, Qin Q, Xiao Y, Woods D, Chen YE, Yang Q (2004) Peroxisome proliferator-activated receptor [delta] activates fatty acid oxidation in cultured neonatal and adult cardiomyocytes. Biochem Biophys Res Commun 313:277–286
Gilde AJ, van der Lee KA, Willemsen PH, Chinetti G, van der Leij FR, van der Vusse GJ, Staels B, van Bilsen M (2003) Peroxisome proliferator-activated receptor (PPAR) alpha and PPARbeta/delta, but not PPARgamma, modulate the expression of genes involved in cardiac lipid metabolism. Circ Res 92:518–524
Mukherjee R, Jow L, Croston GE, Paterniti JR Jr (1997) Identification, characterization, and tissue distribution of human peroxisome proliferator-activated receptor (PPAR) isoforms PPARgamma2 versus PPARgamma1 and activation with retinoid X receptor agonists and antagonists. J Biol Chem 272:8071–8076
Mehrabi MR, Haslmayer P, Humpeler S, Strauss-Blasche G, Marktl W, Tamaddon F, Serbecic N, Wieselthaler G, Thalhammer T, Glogar HD, Ekmekcioglu C (2003) Quantitative analysis of peroxisome proliferator-activated receptor gamma (PPARgamma) expression in arteries and hearts of patients with ischaemic or dilated cardiomyopathy. Eur J Heart Fail 5:733–739
Xu Y, Gen M, Lu L, Fox J, Weiss SO, Brown RD, Perlov D, Ahmad H, Zhu P, Greyson C, Long CS, Schwartz GG (2005) PPAR-gamma activation fails to provide myocardial protection in ischemia and reperfusion in pigs. Am J Physiol Heart Circ Physiol 288:H1314–H1323
Liu LS, Tanaka H, Ishii S, Eckel J (1998) The new antidiabetic drug MCC-555 acutely sensitizes insulin signaling in isolated cardiomyocytes. Endocrinology 139:4531–4539
Duan SZ, Ivashchenko CY, Russell MW, Milstone DS, Mortensen RM (2005) Cardiomyocyte-specific knockout and agonist of peroxisome proliferator-activated receptor-gamma both induce cardiac hypertrophy in mice. Circ Res 97:372–379
Finck BN, Lehman JJ, Leone TC, Welch MJ, Bennett MJ, Kovacs A, Han X, Gross RW, Kozak R, Lopaschuk GD, Kelly DP (2002) The cardiac phenotype induced by PPARalpha overexpression mimics that caused by diabetes mellitus. J Clin Invest 109:121–130
Cheng L, Ding G, Qin Q, Huang Y, Lewis W, He N, Evans RM, Schneider MD, Brako FA, Xiao Y, Chen YE, Yang Q (2004) Cardiomyocyte-restricted peroxisome proliferator-activated receptor-delta deletion perturbs myocardial fatty acid oxidation and leads to cardiomyopathy. Nat Med 10:1245–1250
Brinkmann JF, Abumrad NA, Ibrahimi A, van der Vusse GJ, Glatz JF (2002) New insights into long-chain fatty acid uptake by heart muscle: a crucial role for fatty acid translocase/CD36. Biochem J 367:561–570
Chiu HC, Kovacs A, Blanton RM, Han X, Courtois M, Weinheimer CJ, Yamada KA, Brunet S, Xu H, Nerbonne JM, Welch MJ, Fettig NM, Sharp TL, Sambandam N, Olson KM, Ory DS, Schaffer JE (2005) Transgenic expression of fatty acid transport protein 1 in the heart causes lipotoxic cardiomyopathy. Circ Res 96:225–233
Braissant O, Foufelle F, Scotto C, Dauca M, Wahli W (1996) Differential expression of peroxisome proliferator-activated receptors (PPARs): tissue distribution of PPAR-alpha, -beta, and -gamma in the adult rat. Endocrinology 137:354–366
Djouadi F, Weinheimer CJ, Saffitz JE, Pitchford C, Bastin J, Gonzalez FJ, Kelly DP (1998) A gender-related defect in lipid metabolism and glucose homeostasis in peroxisome proliferator-activated receptor alpha-deficient mice. J Clin Invest 102:1083–1091
Leone TC, Weinheimer CJ, Kelly DP (1999) A critical role for the peroxisome proliferator-activated receptor alpha (PPARalpha) in the cellular fasting response: the PPARalpha-null mouse as a model of fatty acid oxidation disorders. Proc Natl Acad Sci USA 96:7473–7478
Kersten S, Seydoux J, Peters JM, Gonzalez FJ, Desvergne B, Wahli W (1999) Peroxisome proliferator-activated receptor alpha mediates the adaptive response to fasting. J Clin Invest 103:1489–1498
Brandt JM, Djouadi F, Kelly DP (1998) Fatty acids activate transcription of the muscle carnitine palmitoyltransferase I gene in cardiac myocytes via the peroxisome proliferator-activated receptor alpha. J Biol Chem 273:23786–23792
Stavinoha MA, RaySpellicy JW, Essop MF, Graveleau C, Abel ED, Hart-Sailors ML, Mersmann HJ, Bray MS, Young ME (2004) Evidence for mitochondrial thioesterase 1 as a peroxisome proliferator-activated receptor-alpha-regulated gene in cardiac and skeletal muscle. Am J Physiol Endocrinol Metab 287:E888–E895
Campbell FM, Kozak R, Wagner A, Altarejos JY, Dyck JR, Belke DD, Severson DL, Kelly DP, Lopaschuk GD (2002) A role for peroxisome proliferator-activated receptor alpha (PPARalpha) in the control of cardiac malonyl-CoA levels: reduced fatty acid oxidation rates and increased glucose oxidation rates in the hearts of mice lacking PPARalpha are associated with higher concentrations of malonyl-CoA and reduced expression of malonyl-CoA decarboxylase. J Biol Chem 277:4098–4103
Lu B, Jiang YJ, Zhou Y, Xu FY, Hatch GM, Choy PC (2005) Cloning and characterization of murine 1-acyl-sn-glycerol 3-phosphate acyltransferases and their regulation by PPARalpha in murine heart. Biochem J 385:469–477
Vance JE, Vance DE (1985) The role of phosphatidylcholine biosynthesis in the secretion of lipoproteins from hepatocytes. Can J Biochem Cell Biol 63:870–881
Murphy DJ, Vance J (1999) Mechanisms of lipid-body formation. Trends Biochem Sci 24:109–115
Yamaguchi T, Matsushita S, Motojima K, Hirose F, Osumi T (2006) MLDP, a novel PAT family protein localized to lipid droplets and enriched in the heart, is regulated by peroxisome proliferator-activated receptor alpha. J Biol Chem 281:14232–14240
Boren J, Veniant MM, Young SG (1998) Apo B100-containing lipoproteins are secreted by the heart. J Clin Invest 101:1197–1202
Veniant MM, Zlot CH, Walzem RL, Pierotti V, Driscoll R, Dichek D, Herz J, Young SG (1998) Lipoprotein clearance mechanisms in LDL receptor-deficient “Apo-B48-only” and “Apo-B100-only” mice. J Clin Invest 102:1559–1568
McCormick SP, Ng JK, Veniant M, Boren J, Pierotti V, Flynn LM, Grass DS, Connolly A, Young SG (1996) Transgenic mice that overexpress mouse apolipoprotein B. Evidence that the DNA sequences controlling intestinal expression of the apolipoprotein B gene are distant from the structural gene. J Biol Chem 271:11963–11970
Nielsen LB, Veniant M, Boren J, Raabe M, Wong JS, Tam C, Flynn L, Vanni-Reyes T, Gunn MD, Goldberg IJ, Hamilton RL, Young SG (1998) Genes for apolipoprotein B and microsomal triglyceride transfer protein are expressed in the heart: evidence that the heart has the capacity to synthesize and secrete lipoproteins. Circulation 98:13–16
Nielsen LB, Bartels ED, Bollano E (2002) Overexpression of apolipoprotein B in the heart impedes cardiac triglyceride accumulation and development of cardiac dysfunction in diabetic mice. J Biol Chem 277:27014–27020
Bjorkegren J, Veniant M, Kim SK, Withycombe SK, Wood PA, Hellerstein MK, Neese RA, Young SG (2001) Lipoprotein secretion and triglyceride stores in the heart. J Biol Chem 276:38511–38517
Yokoyama M, Yagyu H, Hu Y, Seo T, Hirata K, Homma S, Goldberg IJ (2004) Apolipoprotein B production reduces lipotoxic cardiomyopathy: studies in heart-specific lipoprotein lipase transgenic mouse. J Biol Chem 279:4204–4211
Ameen C, Edvardsson U, Ljungberg A, Asp L, Akerblad P, Tuneld A, Olofsson SO, Linden D, Oscarsson J (2005) Activation of peroxisome proliferator-activated receptor alpha increases the expression and activity of microsomal triglyceride transfer protein in the liver. J Biol Chem 280:1224–1229
Taegtmeyer H (2002) Switching metabolic genes to build a better heart. Circulation 106:2043–2045
Razeghi P, Young ME, Alcorn JL, Moravec CS, Frazier OH, Taegtmeyer H (2001) Metabolic gene expression in fetal and failing human heart. Circulation 104:2923–2931
Sack MN, Rader TA, Park S, Bastin J, McCune SA, Kelly DP (1996) Fatty acid oxidation enzyme gene expression is downregulated in the failing heart. Circulation 94:2837–2842
Sack MN, Disch DL, Rockman HA, Kelly DP (1997) A role for Sp and nuclear receptor transcription factors in a cardiac hypertrophic growth program. Proc Natl Acad Sci USA 94:6438–6443
Depre C, Shipley GL, Chen W, Han Q, Doenst T, Moore ML, Stepkowski S, Davies PJ, Taegtmeyer H (1998) Unloaded heart in vivo replicates fetal gene expression of cardiac hypertrophy. Nat Med 4:1269–1275
Young ME, Laws FA, Goodwin GW, Taegtmeyer H (2001) Reactivation of peroxisome proliferator-activated receptor alpha is associated with contractile dysfunction in hypertrophied rat heart. J Biol Chem 276:44390–44395
Barger PM, Kelly DP (2000) PPAR signaling in the control of cardiac energy metabolism. Trends Cardiovasc Med 10:238–245
Jamshidi Y, Montgomery HE, Hense HW, Myerson SG, Torra IP, Staels B, World MJ, Doering A, Erdmann J, Hengstenberg C, Humphries SE, Schunkert H, Flavell DM (2002) Peroxisome proliferator-activated receptor alpha gene regulates left ventricular growth in response to exercise and hypertension. Circulation 105:950–955
Wayman NS, Hattori Y, McDonald MC, Mota-Filipe H, Cuzzocrea S, Pisano B, Chatterjee PK, Thiemermann C (2002) Ligands of the peroxisome proliferator-activated receptors (PPAR-gamma and PPAR-alpha) reduce myocardial infarct size. Faseb J 16:1027–1040
Ogata T, Miyauchi T, Sakai S, Irukayama-Tomobe Y, Goto K, Yamaguchi I (2002) Stimulation of peroxisome-proliferator-activated receptor alpha (PPAR alpha) attenuates cardiac fibrosis and endothelin-1 production in pressure-overloaded rat hearts. Clin Sci (Lond) 103 (Suppl 48):284S–288S
Sakai S, Miyauchi T, Irukayama-Tomobe Y, Ogata T, Goto K, Yamaguchi I (2002) Peroxisome proliferator-activated receptor-gamma activators inhibit endothelin-1-related cardiac hypertrophy in rats. Clin Sci (Lond) 103 (Suppl 48):16S–20S
Osorio JC, Stanley WC, Linke A, Castellari M, Diep QN, Panchal AR, Hintze TH, Lopaschuk GD, Recchia FA (2002) Impaired myocardial fatty acid oxidation and reduced protein expression of retinoid X receptor-alpha in pacing-induced heart failure. Circulation 106:606–612
Planavila A, Rodriguez-Calvo R, Jove M, Michalik L, Wahli W, Laguna JC, Vazquez-Carrera M (2005) Peroxisome proliferator-activated receptor beta/delta activation inhibits hypertrophy in neonatal rat cardiomyocytes. Cardiovasc Res 65:832–841
Ding G, Cheng L, Qin Q, Frontin S, Yang Q (2006) PPARdelta modulates lipopolysaccharide-induced TNFalpha inflammation signaling in cultured cardiomyocytes. J Mol Cell Cardiol 40:821–828
Larsen TM, Toubro S, Astrup A (2003) PPARgamma agonists in the treatment of type II diabetes: is increased fatness commensurate with long-term efficacy? Int J Obes Relat Metab Disord 27:147–161
Matsusue K, Haluzik M, Lambert G, Yim SH, Gavrilova O, Ward JM, Brewer B Jr, Reitman ML, Gonzalez FJ (2003) Liver-specific disruption of PPARgamma in leptin-deficient mice improves fatty liver but aggravates diabetic phenotypes. J Clin Invest 111:737–747
Hevener AL, He W, Barak Y, Le J, Bandyopadhyay G, Olson P, Wilkes J, Evans RM, Olefsky J (2003) Muscle-specific Pparg deletion causes insulin resistance. Nat Med 9:1491–1497
Norris AW, Chen L, Fisher SJ, Szanto I, Ristow M, Jozsi AC, Hirshman MF, Rosen ED, Goodyear LJ, Gonzalez FJ, Spiegelman BM, Kahn CR (2003) Muscle-specific PPARgamma-deficient mice develop increased adiposity and insulin resistance but respond to thiazolidinediones. J Clin Invest 112:608–618
Kliewer SA, Forman BM, Blumberg B, Ong ES, Borgmeyer U, Mangelsdorf DJ, Umesono K, Evans RM (1994) Differential expression and activation of a family of murine peroxisome proliferator-activated receptors. Proc Natl Acad Sci USA 91:7355–7359
Asakawa M, Takano H, Nagai T, Uozumi H, Hasegawa H, Kubota N, Saito T, Masuda Y, Kadowaki T, Komuro I (2002) Peroxisome proliferator-activated receptor gamma plays a critical role in inhibition of cardiac hypertrophy in vitro and in vivo. Circulation 105:1240–1246
Yamamoto K, Ohki R, Lee RT, Ikeda U, Shimada K (2001) Peroxisome proliferator-activated receptor gamma activators inhibit cardiac hypertrophy in cardiac myocytes. Circulation 104:1670–1675
Nemoto S, Razeghi P, Ishiyama M, De Freitas G, Taegtmeyer H, Carabello BA (2005) PPAR-gamma agonist, rosiglitazone, ameliorates ventricular dysfunction in experimental chronic mitral regurgitation. Am J Physiol Heart Circ Physiol 288(1):H77–H82
Shiomi T, Tsutsui H, Hayashidani S, Suematsu N, Ikeuchi M, Wen J, Ishibashi M, Kubota T, Egashira K, Takeshita A (2002) Pioglitazone, a peroxisome proliferator-activated receptor-gamma agonist, attenuates left ventricular remodeling and failure after experimental myocardial infarction. Circulation 106:3126–3132
Khandoudi N, Delerive P, Berrebi-Bertrand I, Buckingham RE, Staels B, Bril A (2002) Rosiglitazone, a peroxisome proliferator-activated receptor-gamma, inhibits the Jun NH(2)-terminal kinase/activating protein 1 pathway and protects the heart from ischemia/reperfusion injury. Diabetes 51:1507–1514
Yue Tl TL, Chen J, Bao W, Narayanan PK, Bril A, Jiang W, Lysko PG, Gu JL, Boyce R, Zimmerman DM, Hart TK, Buckingham RE, Ohlstein EH (2001) In vivo myocardial protection from ischemia/reperfusion injury by the peroxisome proliferator-activated receptor-gamma agonist rosiglitazone. Circulation 104:2588–2594
Song J, Knepper MA, Hu X, Verbalis JG, Ecelbarger CA (2004) Rosiglitazone activates renal sodium- and water-reabsorptive pathways and lowers blood pressure in normal rats. J Pharmacol Exp Ther 308:426–433
Guan Y, Hao C, Cha DR, Rao R, Lu W, Kohan DE, Magnuson MA, Redha R, Zhang Y, Breyer MD (2005) Thiazolidinediones expand body fluid volume through PPARgamma stimulation of ENaC-mediated renal salt absorption. Nat Med 11:861–866
Mann DL, Young JB (1994) Basic mechanisms in congestive heart failure. Recognizing the role of proinflammatory cytokines. Chest 105:897–904
Stanley WC, Recchia FA, Lopaschuk GD (2005) Myocardial substrate metabolism in the normal and failing heart. Physiol Rev 85:1093–1129
Acknowledgments
This work was partially supported by grants from the NIH (S06GM08248 and 1R01HL085499-01), a scientist development award from the American Heart Association national center, and an institutional grant from NIH (NIH/NIHGMS S). We thank Kevin Yang for editing and proofreading the manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Yang, Q., Li, Y. Roles of PPARs on regulating myocardial energy and lipid homeostasis. J Mol Med 85, 697–706 (2007). https://doi.org/10.1007/s00109-007-0170-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00109-007-0170-9