
Abstract
We studied the variability and plasticity of individual aggressiveness in a social insect, describing and quantifying the sting extension response (SER) of the common wasp Vespula vulgaris. As a proxy for individual aggressiveness we measured the SER of individual wasps, scoring the extent by which the sting was extruded in response to a mild electric shock (7.5 or 12 V for 2 s) on a scale from 0 to 1. We found that wasps vary greatly in their stinging propensity and aggression thresholds and that individuals change their SER during their life. Extremely aggressive or docile phenotypes, showing at first consistent mutual differences on different days, tended to converge over time and developed comparable SER responses later in their life. Older individuals tended to be more aggressive. Wasp size was not related to the stinging phenotype. Wasp foragers had a less pronounced sting extension than individuals previously involved in nest defense. For the same individual, the aggressive response was proportional to the intensity of the negative stimulus. We discuss the advantages of the SER bioassay as a tool to measure individual aggressiveness, plasticity and inter-individual variability in the Aculeata group, and its great potential in comparative and learning studies.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Social insects represent integrated and adaptive units, the result of selective forces operating at both the individual and colony level (Pinter-Wollman and Hubler 2012). Phenotypic differences between individuals of the same social insect colony can be striking, and in fact are regarded as a feature of insect societies (Jeanson and Weidenmüller 2013). Indeed, inter-individual variability is acknowledged as a necessary condition for one mainstay of eusociality, the division of labor among colony members (Jeanson and Weidenmüller 2013). The division of labor within social insect colonies has been explained by variability in (1) individual, innate, thresholds and (2) experience-based, self-reinforcement processes; two non-mutually exclusive mechanisms (Beshers and Fewell 2001). At the individual level, the role of experience is recognized as central in shaping the individual behavioral phenotype, changing over time via learning processes (Jeanson and Weidenmüller 2013).
Inter-individual differences have been long overlooked in quantitative ecological studies, but have gained new attention during the last decade, notably in the field of “animal personality/behavioral syndrome” research (Violle et al. 2012). A number of studies across the animal kingdom have demonstrated dramatic differences among individual behavioral phenotypes (Bolnick et al. 2003). Since individual phenotypes are the focus of natural selection, any variation at this level has major evolutionary and ecological consequences (Bolnick et al. 2011; Wolf and Weissing 2012).
In biological terms, aggression is a widespread solution by animals to the problems of self-preservation, protection of young and resource competition (Archer 1988). Aggressiveness is a fundamental behavioral and personality trait, varying greatly among individuals (Dingemanse and Wolf 2013; Grinsted et al. 2013). In social hymenopterans, the study of aggressive or defensive behavior has been classically undertaken in the context of nestmate recognition and enemy identification (e.g., Guerrieri and D’Ettorre 2008; Breed et al. 2004). For honey bees and other social Aculeata, one important component of aggressive/defensive behavior is the act of stinging (Paxton et al. 1994).
The stinging response (stinging reflex, sting extension response) to electric stimulation was described in Apis mellifera during the 1980s (Balderrama et al. 1987; Kolmes and Fergusson-Kolmes 1989) and has been used as a bioassay since this time. The sting extension response (SER) was measured via the stimulus intensity (Kolmes and Fergusson-Kolmes 1989; Paxton et al. 1994) or the time necessary to elicit it (Uribe-Rubio et al. 2008). The SER was initially recorded as a binary response (Balderrama et al. 1987, 2002), with some authors refining the scoring system by recording the extent to which the sting was extruded, adding intermediate levels of response corresponding to partial sting extension (Lenoir et al. 2006).
The SER has been quantified to measure honey bees’ level of aggressiveness and to discriminate among different strains (Kolmes and Fergusson-Kolmes 1989; Uribe-Rubio 2013), colonies, worker groups (Paxton et al. 1994; Uribe-Rubio 2013) and individuals from the same hive (Lenoir et al. 2006). Variation in SER was found in relation to honey bees’ age and within-colony social environment (Paxton et al. 1994), spatial segregation in the hive (Uribe-Rubio et al. 2008), task performed (Uribe-Rubio et al. 2008, 2013), and paternity line (Lenoir et al. 2006).
It is important to highlight that, depending on the organism under examination, the same type of behavioral response (e.g., SER) can have a radically different meaning in biological terms. Although honey bees can sting other insects without harming themselves, their sting is strongly barbed and remains embedded in mammalian flesh (Vetter et al. 1999). Given the characteristic sting autotomy, anchoring and the consequent abdominal rupture, honey bees’ stinging behavior can represent an extreme form of suicidal colony defense (Paxton et al. 1994; Shorter and Rueppell 2011). Relatively little is known about the stinging behavior of other eusocial bees (reviewed in Breed et al. 2004) and we are not aware of any studies quantifying the stinging response in wasps.
For wasps, aggression is a common practice, e.g., in foraging activities (Parrish 1984; Raveret Richter 2000), social dynamics and in high-risk situations (Ross and Matthews 1991). The majority of aculeate wasps are solitary and use their stings to paralyze hosts more or less permanently and then carry them to a shelter where an egg is laid on the host (Raveret Richter 2000). For social aculeate wasps, prey are generally killed with the characteristically strong mandibles, rather than stinging (Edwards 1980; Raveret Richter 2000). Nevertheless, several authors (reviewed in Spradbery 1973; Olson 2000) documented cases in which foraging social wasps used their sting when grappling with particularly large and active prey. Vespa orientalis was reported to make regular use of the sting when attacking A. mellifera in Israel (Ishay et al. 1967, cited in Spradbery 1973). This evidence contradicts some statements that social wasps kill their prey using exclusively their mandibles (e.g., Akre and Myhre 1992, in Olson 2000; Archer 2012).
Social wasps also utilize their stings during frequent inter- and intra-specific conflicts among gynes, workers and castes (Archer 2012). During spring, mortal fights between queens competing for nesting sites are the norm (Greene 1991; Matsuura and Yamane 1990; Hunt 2007). In some yellowjackets (e.g., Vespula, Dolichovespula), usurpation may be more common than nest initiation (Greene 1991), and disputes appear more frequent among conspecifics (Buck et al. 2008). The sting can also be used by workers to kill the males (Monceau et al. 2013), or during matricide when workers kill their queen (Loope 2014). In general, aggression (e.g., mauling, biting and grappling) among workers is common, both at the foraging sites (Parrish 1984; Raveret Richter 2000) and inside the nest (Akre et al. 1976; O’Donnell and Jeanne 1995). Fighting, including attempted stinging, usually involves workers with developed ovaries, both in colonies with and without a queen (Archer 2012).
Stinging is the immediate response in a life-threatening situation, both for the individual wasps and the colonies. Intruders inside a social nest are stung in rapid actions, involving one or more co-operating wasps (Edwards 1980). If the nest is disturbed from outside, agitated workers rush out and eventually take flight, with a minority of individuals normally stinging the target (Greene 1991). In species with large colonies, active nest defense can involve hundreds of individuals and mass stinging can potentially be lethal to humans (Vetter et al. 1999). Vespine wasps, including yellowjackets, are particularly aggressive and are the group usually responsible for hymenopteran stinging events involving humans (Vetter et al. 1999). Wasps defend their nests fiercely (Edwards 1980) and may also sting disturbers while foraging, especially in the late summer and early fall, when many hungry queen larvae must be fed with flesh (Spradbery 1973). Given the sting morphology, social wasps are usually able to sting multiple times (Vetter et al. 1999), although sting anchoring routinely occurs in some species such as V. maculifrons (Greene 1991).
Because of their aggressiveness, predatory habits and sting features, workers of social wasps have the potential to sting more than once throughout their life, in a variety of contexts. As a consequence, it is possible that the stinging phenotype of one individual may be shaped by previous experience. As highlighted in the past, there are a number of unresolved problems with the study of wasps’ stinging behavior (Greene 1991; Olson 2000). In general, the descriptions of specific aggressiveness for wasps are mostly anecdotal, qualitative and contradictory. Knowledge of the factors regulating individual wasp response thresholds for various disturbing stimuli is very limited. Alarm pheromones, worker age, colony size, brood population, previous disturbance, and meteorological conditions represent some commonly discussed factors influencing the stinging behavior (Gaul 1952; Ross and Matthews 1991; Potter 1964; London and Jeanne 1996, 2003; Reed and Landolt 2000).
In the present paper we quantify individual wasps’ stinging propensity, with a laboratory bioassay. As a proxy for aggressiveness, we describe and measure the sting extension of the common wasp, V. vulgaris, looking at its individual plasticity and inter-individual variation. Taking into account individual age and size, we measured variation in individual SER throughout the adult life of these wasps, relating it to previous experience, presumptive task performed by the tested individuals and intensity of noxious stimulation.
Materials and methods
A V. vulgaris nest (colony 1) was excavated in Christchurch, New Zealand, on the 19th of January 2014. The nest was boxed and carried to a greenhouse facility at the Plant and Food Research Biosecurity laboratory, Lincoln. Wasps were allowed to forage outside of the nest box, in the grounds surrounding the laboratory. On the 18th of March 2014, the colony was anesthetized with carbon dioxide, the box opened, and three pieces of nest combs with capped brood were removed and placed in an incubator (30 °C, 50 % relative humidity, and complete darkness). A total of 292 known-age workers were obtained between the 19th of March and the 7th of April 2014. Newly emerged adults were collected daily, caged and tested for their SER. The cages were 50-ml plastic jars modified by covering either end with a metal mesh. The individually caged wasps were fed daily by imbibing with 1 ml of 30 % sugar solution one cotton bud suspended from the lid mesh. The cages were numbered and organized on trays lined with absorbent paper that was changed daily, and kept in the incubator under the same conditions as the nest combs. Two additional V. vulgaris nests were located in the grounds of the research facility at Lincoln and exiting wasps from both colonies (colony 2, 3) were collected and tested (see below).
SER scoring
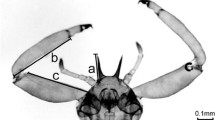
The caged, known-age individuals were lightly anesthetized with carbon dioxide and, when subdued but still moving, harnessed by holding their petiole from the side with metal forceps (Bioquip, feather weight forceps, narrow tip), kept closed with one clip (Fig. 1, Fig. 1S). Each individual was allowed 30 min to acclimate to the harnessing before testing. The forceps restraining the wasp were connected to one polarized cable, while the other one was connected to a metal arch covered in conductive gel (Spectra electrode gel, contents 8.5 OZ, 250 GMS, salt free, Parker laboratories Inc.) (Figure 2S). The electric stimulation was delivered for 2 s by touching the wasp’s cervix with the metal arch. The standard stimulus intensity was 7.5 Volts (Balderrama et al. 2002). Each wasp was tested four consecutive times (rounds), with an inter-round interval of 1 min. Responses during each stimulation were recorded by observing the tested individuals’ abdomen from the side, adopting the four-level scoring system proposed by Lenoir et al. (2006) for honey bees. The absence of any response was scored 0, the response was scored 0.33 when the sting was extended less than the half of its length, 0.67 when it was extended between half and all its length and finally, the response was scored 1 when the sting was fully extended and the sting chamber was completely open. The wasps’ final SER score was obtained by averaging the four consecutive measures and hence varied between 0 (minimum aggression response) and 1 (maximum aggression response) (Fig. 3S). Student’s pairwise t tests were used to compare the individual SER between the first trial’s rounds.

a Individual plasticity of the sting extension response (SER, circles) throughout life, on the basis of previous noxious experience. Points are mean ± SE. Continuous lines significant differences between consecutive trials. Pictures phenotypes at the two extremes (docile vs aggressive). The inset histogram in the box shows the distribution of all the first trial’s SER scores (n = 122) and the two subgroups (white and black bar). b Individual SER plasticity for the initially aggressive group (SER MAX, black points, individuals with the highest stinging score recorded on the first trial) and the initially docile group (SER MIN, white points, individuals showing no response on the first day trial). Trials were separated by approximately 48 h. Points are mean ± SE. p values refer to difference in SER score between aggressive and docile group, tested trial by trial. ***p < 0.001, ns p > 0.05. Please refer to Table 2S (supplementary material), for details of the statistical comparisons
SER in relation to individual age and size
A total of 264 known-age individuals were tested for the first time at different ages (1–27 days of adult life). To study the relationship between individual size and stinging propensity, a subset of 112 wasps was subsequently preserved in 90 % ethanol at −20 °C. To estimate individual body size, we measured the wasps’ head width, using a digital caliper to the nearest 0.01 mm. To separately test for an effect of age and size on the SER score, linear models were used. We also performed a multiple regression testing the effect of age and size simultaneously, using data from 105 individuals where we had both size and age data.
Plasticity in individual SER throughout life
One subset of 122 known-age wasps was kept in individual cages (see above) and the individuals were retested at 48 h intervals (trials) throughout their remaining life. A total of 112 individuals were tested at least twice (1st test age: 10.0 ± 5.5, 1–19 days). Their average adult lifespan was 18.1 ± 5.1, with a range of 8–28 days.
On the base of the initial SER score at day 1, two subgroups were distinguished: 31 individuals had an SER score = 0 for the first trial (no sting extension response on the four consecutive rounds). Of these, 26 individuals could be retested and were pooled in the “docile group” (1st test age: 9.4 ± 4.2, 1–13 days). The 29 individuals showing the highest SER score on the first trial (=0.83) were assigned to the “aggressive group” (1st test age: 8.1 ± 5.7, 1–16 days).
To analyze the change of the individual SER during lifetime, repeated measures analysis of variance with Bonferroni confidence interval adjustment was performed in pairwise comparisons among trials (days). Since data were not normally distributed, Kruskal–Wallis test was used to compare the SER score of the docile group vs aggressive group, trial by trial.
SER and presumptive task (forager/guard)
Two task-related groups of wasps were sampled from colony 1. Over 3 days, we captured 52 incoming foragers with a butterfly net, while flying with their foraging loads in proximity of the nest. On the same days, after the capturing the foragers, we hit the substrate supporting the nest with a stick, provoking a colony defense reaction, and captured a total of 45 “guards” that emerged from the nest. Wasps from both groups were transferred in individual cages, and their SER examined within 2 h. Since data were not normally distributed, Kruskal–Wallis test was used to test for differences in the SER between the groups of foragers and guards.
SER and stimulus intensity
Individuals exiting the three colonies were captured and then tested at 7.5 and 12.5 Volts (colony 1: 78 individuals; colony 2: 26 individuals; colony 3: 12 individuals). For each individual, the inter-test interval was set to 30 min and the voltage order established randomly. Student’s pairwise t test was used to compare the individual SER between the two tests.
All data analyses were performed using R 3.1.0 (R Development Core Team Team 2012) and IBM SPSS Statistics 21 (IBM Corp. 2012).
Results
SER in relation to age and size
Substantial and consistent variation in individual stinging response was found between wasps (average SER score ± SD: 0.64 ± 0.40; n = 292). On the first trial, one individual out of five showed no response to the stimulation and half of the wasps showed the full sting extension on at least one round. Within each trial, the individual SER score was consistent across rounds (Table 1S). Only starting from the second trial, five individuals (one for the next two) scored 1 (full sting extension on the four consecutive rounds). Despite wide variability in SER scores, a positive relationship between age and SER was found (multiple R 2 = 0.066, \( R_{\text{adj}}^{ 2} \) = 0.062, F 1,262 = 18.45, p < 0.0001) (Fig. 2). When both age and size were considered, for a subset of 105 individuals, no significant effect of the two factors was found (effect of age: F 1,102 = 1.35, p = 0.251; effect of size: F 1,102 = 0.09, p = 0.765. Model R 2 = 0.013, \( R_{\text{adj}}^{ 2} \) = −0.006).

Regression between the sting extension response (SER) and the age of the wasp. Point symbols are mean ± SE. The point’s diameter is proportional to the sample size of each age (total n = 264, n per day = 2–47)
Individuals varied considerably in size (3.08–3.71 mm head width). However, no relationship was found between individual size and SER score (Multiple R 2 = 0.0024, \( R_{\text{adj}}^{ 2} \) = −0.007, F 1,110 = 0.264, p = 0.608) (Fig. 3).

Correlation between the sting extension response (SER) and the size of the wasp. Each point represents one individual and its SER score at the first test
Individual SER plasticity throughout life
There was a significant change in individual SER over successive trials and days. Overall, the average stinging response was stronger on the second trial and subsequently decreased (Fig. 1a). For the group of individuals retested, the distribution of SER on the first trial was bimodal, with 21 % of individuals displaying an SER score of 0 (pooled in the “docile” group) and 24 % showing an SER score of 0.83 (pooled in the “aggressive” group) (Fig. 1a inset). The docile and aggressive groups showed opposite trends over time. The aggressive group displayed a significantly higher SER score than the docile group for the first three trials, but with a decline in SER over time, while the docile group showed an increase in SER over the first seven trials. The two groups showed a similar score from the fourth to the seventh trial (Fig. 1b; Table 2S). Some individuals within these groups never changed, always displaying very low or very high SER scores.
SER and presumptive task (forager/guard)
On average, the SER score of presumptive guards was threefold higher than the SER score of the foragers (χ2 = 19.54, df = 1, p < 0.0001). Nevertheless, inter-individual variation within the two groups was wide (Fig. 4).

Sting extension response (SER) and presumptive task undertaken by tested individuals. “Foragers” were captured on their way back to the nest. “Guards” were collected after striking the nest box. Boxes represent the lower and upper quartile, the bold line is the median and whiskers represent extreme values of SER score, with the circles identifying outliers. ***p < 0.001
SER and stimulus intensity
The average individual from all the three colonies showed a significantly stronger stinging response in correspondence to higher stimulus intensity. The SER score at 12.5 V was up to almost four times higher than at 7.5 V (colony 1: t = −13.70, df = 77, p < 0.0001; colony 2: t = −7.84, df = 25, p < 0.0001; colony 3: t = −3.06, df = 11, p = 0.0110). Some individuals showed no or little response at both stimulus intensity levels (Fig. 5).

Sting extension response (SER) of individuals from three V. vulgaris colonies, tested twice at different intensities of noxious stimulus (electric current), in random order. Boxes represent the lower and upper quartile, the bold line is the median and whiskers represent extreme values of SER score, with the circles identifying outliers. ***p < 0.001, *p < 0.05
Discussion
Given the ubiquitous occurrence of direct competitive interactions, aggression plays a major role in fitness outcomes (Archer 1988). Despite being reported numerous times in many contexts, differences in defensiveness/aggressiveness among social hymenopteran species (e.g., Breed et al. 2004; Greene 1991), between and within specific colonies (e.g., Jeanne 1988; Pinter-Wollman 2012) have been described mostly qualitatively. Scoring the sting extension response, as a proxy for aggressiveness, can quantify an important trait of the individual behavioral phenotype (Dingemanse and Wolf 2013; Grinsted et al. 2013). The SER bioassay hence represents a potential tool for the study of inter-individual variation and animal personalities in an evolutionary perspective. A growing body of literature is showing the importance to ecological studies of quantifying individual variability (Bolnick et al. 2003, 2011; Violle et al. 2012; Wolf and Weissing 2012). We believe that the sting extension response is an opportunity to study behavioral variability among the social Aculeata, both at an individual level and in comparative studies.
We tested for, and found, substantial and consistent variation in the stinging thresholds of common wasp nestmates. Although if data for colony 2 and 3 are very limited, this behavioral pattern was consistent for all wasp colonies examined. The individual SER measure proved to be repeatable across rounds and trials (Fig. 1b, Table 1S, 2S). The plasticity of SER recorded throughout a wasp’s life highlights the variability in time of individual behavior. Initially docile wasps developed a stronger SER, while initially aggressive wasps showed an opposite trend, almost having a lower SER score compared to the first group later in their life (Trial 6: Fig. 1b; Table 2Sb). These inverse patterns seem to exclude the possibility of a simple sensitization to the electric stimulation (Shettleworth 2010). Also, artificial, asocial and stressful captivity conditions would likely affect individuals similarly. Our data do not allow to speculate on the significance of these plastic changes in a natural context, but still show that extremely different phenotypes can converge and express similarly, on the base of the same previous experience. Taken together, our empirical findings support theoretical frameworks where individuals show innate differences but change their behavioral phenotype throughout their life (Theraulaz et al. 1998; Beshers and Fewell 2001).
Age was correlated to the stinging propensity of V. vulgaris individuals. This result finds a parallel in field-based studies performed on other social wasp species, where older individuals were found to be more likely to defend the colony (Jeanne et al. 1992; Togni and Giannotti 2010; Monceau et al. 2013). However, when including head width as a covariate, no relationship between age and SER was found, probably due to the high intra-trial variability and reduction in sample size. It must be noticed that age poorly explained the variation observed. Moreover, the isolation of known-age adults might have influenced their behavioral phenotype. The stinging reaction was observed in individuals of all ages, including in one-day-old individuals. Ontogenetic variation in stinging behavior has been reported for honey bees, with older individuals (20-day-old) showing the lowest stinging thresholds (Paxton et al. 1994). The sting reflex was found to be fully developed in bees older than 5–7 days (Breed et al. 2004). The median age at which A. mellifera workers become guards was reported to be 12 days (Lenoir et al. 2006). Older individuals have lower life expectancy and theoretically are predicted to be more likely to perform risky tasks, e.g., nest defense, their anticipated future value to their society being lower (Kolmes and Fergusson-Kolmes 1989).
We found no significant relationship among individual size and SER. Still, it should be noted that small individuals with head width <3.3 mm never displayed an SER score greater than 0.5 (Fig. 3), perhaps suggesting a non-linear relationship. To our knowledge, no study has so far investigated a possible relationship between sting extension and individual size. Vespula spp. colonies are good systems, being characterized by seasonal morphometric changes of workers, with increasing average worker size and variation as the colony develops (Spradbery 1972). Size relationships represent a crucial factor in most predator–prey and intra-specific competitive interactions (e.g., MacNulty et al. 2009; Santoro et al. 2011). Together with age, size is also extremely important in establishing dominance hierarchies in social insects (O’Donnell and Jeanne 1995; Hogendoorn and Velthuis 1999). Dominance relationship among colony members can, in turn, influence the division of labor among and within castes (O’Donnell and Jeanne 1995). Size was also shown to directly determine individual behavioral thresholds: bigger bumble bees (Bombus terrestris), for example, are more likely to forage and have more sensitive antennae, showing lower thresholds to odors (Spaethe et al. 2007).
Despite their intrinsic limitations, measures obtained in solitary, standardized laboratory settings can be reliable predictors of behavior in a social context in the field (Grinsted et al. 2013). The SER bioassay detected wide variability in stinging propensity and discriminated individuals previously involved in colony defense from foragers. These results match field observations that stinging and nest active defense by social hymenopterans involves only a subset of workers (Greene 1991; Breed et al. 2004). For example, Africanized honey bee guards were faster to sting than nest bees (Uribe-Rubio et al. 2008). Given that our wasp “guards” were tested within 2 h from the colony provocation, they might have still been under the effect of an alarm pheromone, for which the composition and latency are still to be investigated (Reed and Landolt 2000).
The stinging response of individuals from the three colonies was more pronounced in association with a more intense electric stimulation. The same individual showed a more aggressive reaction to the stronger of the two noxious stimuli, regardless of which one was experienced first. Similar results were obtained with different groups of honey bee workers, whose responsiveness to electric stimulation was found to increase with higher voltages, up to 8 V (Balderrama et al. 2002).
Variation in SER can have a genetic basis. A number of studies on A. mellifera have demonstrated that guarding is a specialized task performed by a few, genetically predisposed individuals (Robinson 1992; Breed et al. 2004; Hunt 2007). Inter-individual variability in the honey bee SER has been observed as associated with different paternity lines (Lenoir et al. 2006). Polyandry is rare among social hymenopterans (Strassmann 2001) and, despite having costs in terms of colony cohesion (Crozier and Fjerdingstad 2001), was linked to colony fitness in, e.g., V. maculifrons (Goodisman et al. 2007). Polyandry is thought to have an adaptive significance by tuning group responses of insect colonies, determining worker subgroups with different task-related thresholds (Oldroyd and Fewell 2007). Both A. mellifera and Vespula are peculiar amongst hymenopterans, showing marked polyandry (Strassmann 2001, Hanna et al. 2013). Paternal effects in wasp’s aggressiveness await to be tested. However, we predict that colonies and species with single-mating queens will show lower levels of aggressive variability among nestmates, measurable with the SER bioassay.
A very promising avenue is the SER classical conditioning for research focusing on the differential learning performances among Aculeata. Successful aversive conditioning was done on A. mellifera (e.g., Vergoz et al. 2007). Wasps are traditionally described as explorative, opportunistic foragers, capable of quick associative learning (Raveret Richter 2000; Moreyra et al. 2014). Wasps’ associative learning capabilities await to be scored in the laboratory.
Our findings add to the existing literature by extending earlier SER studies on A. mellifera to a new system, vespine wasps. We quantified the degree individuals from the same insect colony can differ in the SER, providing an example of experience-based convergence of initially different behavioral phenotypes. The quantification of the stinging behavior through the SER score proved to be a repeatable, easily performed procedure to test for innate threshold differences between individual wasps. We encourage the development of standardized SER testing methods and more studies to better understand the biological significance of SER, focusing on the relationship between the measures obtained with this laboratory bioassay and aggressive/defensive interactions in different natural contexts. At these conditions, the SER bioassay has the potential to become a standard measure of aggressiveness, a cardinal personality trait. Even more interestingly, the SER comes with the promise of exciting advancement in comparative and learning studies.
References
Akre RD, Garnett WB, Mac Donald JF, Greene A, Landolt P (1976) Behavior and colony development of Vespula pensylvanica and V. atropilosa (Hymenoptera: Vespidae). J Kansas Entomol Soc 49:63–84
Archer J (1988) The behavioral biology of aggression. Cambridge University Press, Cambridge
Archer ME (2012) Vespine wasps of the world—behavior, ecology & taxonomy of the Vespinae. Siri Scientific Press, Castleton
Balderrama N, Diaz H, Sequeda A, Núñez J, Maldonado H (1987) Behavioral and pharmacological analysis of the stinging response in Africanized and Italian bees. In: Menzel R (ed) Neurobiology and behavior of honeybees. Springer, Berlin, pp 121–122
Balderrama N, Núñez J, Guerrieri F, Giurfa M (2002) Different functions of two alarm substances in the honeybee. J Comp Physiol A 188:485–491
Beshers SN, Fewell JH (2001) Models of division of labor in social insects. Annu Rev Entomol 46:413–440
Bolnick DI, Svanbäck R, Fordyce JA et al (2003) The ecology of individuals: incidence and implications of individual specialization. Am Nat 161:1–28
Bolnick DI, Amarasekare P, Araújo M et al (2011) Why intraspecific trait variation matters in community ecology. Trends Ecol Evol 26:183–192
Breed MD, Guzman-Novoa E, Hunt GJ (2004) Defensive behavior of honey bees: organization, genetics, and comparisons with other bees. Annu Rev Entomol 49:271–298
Buck M, Marshall SA, Cheung DKB (2008) Identification atlas of the Vespidae (Hymenoptera, Aculeata) of the northeastern Nearctic region. Can J Arthropod Identif 5(5). doi:10.3752/cjai.2008.05
Crozier RH, Fjerdingstad EJ (2001) Polyandry in social Hymenoptera—disunity in diversity? Ann Zool Fennici 38:267–285
Dingemanse NJ, Wolf M (2013) Between-individual differences in behavioral plasticity within populations: causes and consequences. Anim Behav 85:1031–1039
Edwards R (1980) Social wasps—their biology and control. Rentokil Limited, East Grinstead
Gaul AT (1952) The awakening and diurnal flight activities of vespine wasps. Proc R Entomol Soc Lond Ser A 27:33–38
Goodisman MAD, Kovacs JL, Hoffman EA (2007) The significance of multiple mating in the social wasp Vespula maculifrons. Evolution 61:2260–2267
Greene (1991) Dolichovespula and Vespula. In: Ross KG, Matthews RW (eds) The social biology of wasps. Cornell University Press, Ithaca
Grinsted L, Pruitt JN, Settepani V, Bilde T (2013) Individual personalities shape task differentiation in a social spider. Proc R Soc B 280:20131407. doi:10.1098/rspb.2013.1407
Guerrieri FJ, D’Ettorre P (2008) The mandible opening response: quantifying aggression elicited by chemical cues in ants. J Exp Biol 211:1109–1113
Hanna C, Cook ED, Thompson AR, Dare LE, Palaski AL, Foote D, Goodisman MAD (2013) Colony social structure in native and invasive populations of the social wasp Vespula pensylvanica. Biol Invasions 16:283–294
Hogendoorn K, Velthuis HHW (1999) Task allocation and reproductive skew in social mass provisioning carpenter bees in relation to age and size. Insect Soc 46:198–207
Hunt J (2007) The evolution of social wasps. Oxford University Press, Oxford
IBM Corp. (2012) IBM SPSS Statistics for Windows, Version 21.0. Armonk, NY: IBM Corp
Jeanne RL (1988) Interindividual behavioral variability in social insects. Westview Press, Boulder
Jeanne RL, Williams N, Yandell B (1992) Age polyethism and defense in a tropical social wasp (Hymenoptera: Vespidae). J Insect Behav 5:211–227
Jeanson R, Weidenmüller A (2013) Interindividual variability in social insects—proximate causes and ultimate consequences. Biol Rev Camb Philos Soc 89:671–687
Kolmes SA, Fergusson-Kolmes LA (1989) Stinging behavior and residual value of worker honey bees (Apis mellifera). J New York Entomol Soc 97:218–231
Lenoir J-C, Laloi D, Dechaume-Moncharmont F-X, Solignac M, Pham MH (2006) Intra-colonial variation of the sting extension response in the honey bee Apis mellifera. Insect Soc 53:80–85
London KB, Jeanne RL (1996) Alarm in a wasp-wasp nesting association: do members signal cross-specifically? Insect Soc 43:211–215
London KB, Jeanne RL (2003) Effects of colony size and stage of development on defense response by the swarm-founding wasp Polybia occidentalis. Behav Ecol Sociobiol 54:539–546
Loope KJ (2014) Why do vespine wasp workers commit matricide? In: 17th Congress of the International Union for the Study of Social Insects (IUSSI), Cairns, Australia, 13–18 July 2014
MacNulty DR, Smith DW, Mech LD, Eberly LE (2009) Body size and predatory performance in wolves: is bigger better? J Anim Ecol 78:532–539
Matsuura M, Yamane S (1990) Biology of the vespine wasps. Springer, Berlin, Heidelberg, New York
Monceau K, Bonnard O, Thiéry D (2013) Relationship between the age of Vespa velutina workers and their defensive behavior established from colonies maintained in the laboratory. Insect Soc 60:437–444
Moreyra S, D’Adamo P, Lozada M (2014) The influence of past experience on wasp choice related to foraging behavior. Insect Sci 21:759–764
O’Donnell S, Jeanne RL (1995) The roles of body size and dominance in division of labor among workers of the eusocial wasp Polybia occidentalis (Olivier) (Hymenoptera: Vespidae). J Kansas Entomol Soc 68:43–50
Oldroyd BP, Fewell JH (2007) Genetic diversity promotes homeostasis in insect colonies. Trends Ecol Evol 22:408–413
Olson EJ (2000) Parachartergus fraternus (Gribodo) (Hymenoptera: Vespidae: Polistinae) uses venom when taking caterpillar prey. Psyche 103:85–93
Parrish M (1984) Factors influencing aggression between foraging yellowjacket wasps, Vespula spp. (Hymenoptera: Vespidae). Ann Entomol Soc Am 77:306–311
Paxton RJ, Sakamoto CH, Rugiga FCN (1994) Modification of honey bee (Apis mellifera L.) stinging behavior by within-colony environment and age. J Apicult Res 33:75–82
Pinter-Wollman N (2012) Personality in social insects: how does worker personality determine colony personality? Curr Zool 58:580–588
Pinter-Wollman N, Hubler J (2012) How is activity distributed among and within tasks in Temnothorax ants? Behav Ecol Sociobiol 66:1407–1420
Potter NB (1964) A study on the biology of the common wasp, Vespula vulgaris L., with special reference to the foraging behaviour. Ph.D. dissertation, University of Bristol
Raveret Richter M (2000) Social wasp (Hymenoptera: Vespidae) foraging behavior. Annu Rev Entomol 45:121–150
Reed HC, Landolt PJ (2000) Application of alarm pheromone to targets by southern yellowjackets (Hymenoptera: Vespidae). Fla Entomol 83:193–196
Robinson GE (1992) Regulation of division of labor in insect societies. Annu Rev Entomol 37:637–665
Ross KG, Matthews RW (1991) The social biology of wasps. Cornell University Press, Ithaca
R Development Core Team (2012) R: a language and environment for statistical computing. doi: ISBN 3-900051-07-0
Santoro D, Polidori C, Asís JD, Tormos J (2011) Complex interactions between components of individual prey specialization affect mechanisms of niche variation in a grasshopper-hunting wasp. J Anim Ecol 80:1123–1133
Shettleworth SJ (2010) Cognition, evolution and behavior, 2nd edn. Oxford University Press, New York
Shorter JR, Rueppell O (2011) A review on self-destructive defense behaviors in social insects. Insect Soc 59:1–10
Spaethe J, Brockmann A, Halbig C, Tautz J (2007) Size determines antennal sensitivity and behavioral threshold to odors in bumblebee workers. Naturwissenschaften 94:733–739
Spradbery PJ (1972) A biometric study of seasonal variation in worker wasps (Hymenoptera: Vespidae). J Ent (A) 47:61–69
Spradbery PJ (1973) Wasps—an account of the biology and natural history of solitary and social wasps. University of Washington Press, Seattle
Strassmann J (2001) The rarity of multiple mating by females in the social Hymenoptera. Insect Soc 48:1–13
Theraulaz G, Bonabeau E, Denuebourg J-N (1998) Response threshold reinforcements and division of labor in insect societies. Proc R Soc B 265:327–332
Togni O, Giannotti E (2010) Colony defense behavior of the primitively eusocial wasp, Mischocyttarus cerberus is related to age. J Insect Sci 10:1–14
Uribe-Rubio JL (2013) Genotype and task influence stinging response thresholds of honeybee (Apis mellifera L.) workers of African and European descent. Open J Ecol 3:279–283
Uribe-Rubio JL, Guzmán-Novoa E, Vázquez-Peláez CG, Hunt GJ (2008) Genotype, task specialization, and nest environment influence the stinging response thresholds of individual Africanized and European honeybees to electrical stimulation. Behav Genet 38:93–100
Vergoz V, Roussel E, Sandoz J-C, Giurfa M (2007) Aversive learning in honeybees revealed by the olfactory conditioning of the sting extension reflex. PLoS One 2:e288. doi:10.1371/journal.pone.0000288
Vetter RS, Visscher PK, Camazine S (1999) Mass envenomations by honey bees and wasps. West J Med 170:223–227
Violle C, Enquist B, McGill B (2012) The return of the variance: intraspecific variability in community ecology. Trends Ecol Evol 27:244–252
Wolf M, Weissing F (2012) Animal personalities: consequences for ecology and evolution. Trends Ecol Evol 27:452–461
Acknowledgments
We thank Lloyd Stringer for his help during nest excavation, Flore Mas and Rafael Barbieri for insightful advice during data collection and analysis, Michael Breed, Kevin Loope and one anonymous reviewer for their suggestions. This work was funded by Victoria University of Wellington.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Santoro, D., Hartley, S., Suckling, D.M. et al. The stinging response of the common wasp (Vespula vulgaris): plasticity and variation in individual aggressiveness. Insect. Soc. 62, 455–463 (2015). https://doi.org/10.1007/s00040-015-0424-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00040-015-0424-4