
Abstract
Abiotic stresses collectively are responsible for crop losses worldwide. Among various abiotic stresses, drought and salinity are the most destructive. Different strategies have been adopted for the management of these stresses. Being complex traits, conventional breeding approaches have shown less success in improving salinity and drought stress tolerance. Roles of compatible solutes in salinity and drought stress tolerance have been studied extensively. At physiological level, osmotic adjustment is an adaptive mechanism involved in drought and/or salinity tolerance and permits the maintenance of turgor pressure under stress conditions. Increasing evidences from series of in vivo and in vitro studies involving physiological, biochemical, genetic, and molecular approaches strongly suggest that osmolytes such as ammonium compounds (polyamines, glycinebetaine, b-alanine betaine, dimethyl-sulfonio propionate and choline-O-sulfate), sugars and sugar alcohols (fructan, trehalose, mannitol, d-ononitol and sorbitol) and amino acids (proline and ectoine) perform important function in adjustment of plants against salinity and drought stresses. Thus, aim of this review is to expose how to osmoprotectants detoxify adverse effect of reactive oxygen species and alleviate drought and salinity stresses. An understanding of the relationship between these two sets of parameters is needed to develop measures for mitigating the damaging impacts of salinity and drought stresses.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
Among abiotic stresses, salinity and drought are of special concerns. They are the most recognizable in arid and semi-arid regions of the world and responsible for decline in crop yield. Plants are continuously facing harsh environment, which is regulated by different environmental factors. Environmental factors may be characterized in two ways. First one is biotic factors comprising living creatures such as parasites, pathogens, herbivores, etc. and second one is abiotic factors including non living matters like salinity, drought, temperature, humidity, light, water, mineral nutrients, heavy metal, radiation, etc. Abiotic factors are imposing adverse impacts on the yield of crops (Vinocur and Altman 2005). There are many evidences which show that abiotic stresses such as salinity and drought, that arrest agricultural production throughout the world (Le and McQueen-Mason 2006; Galvani 2007; Gosal et al. 2009; Wani et al. 2010; Wani and Gosal 2011; Yu et al. 2013; Wani et al. 2013; Yadav et al. 2014). In response to salinity and drought stresses, plants adopt various strategies including modification in different physiological as well as biochemical processes such as signal perceptions and transduction cascades for survival against these stresses (Faical et al. 2009; Ranganayakulu et al. 2013). Studies showed that osmotic regulations in plants during salinity and drought stresses have occurred through higher accumulation of osmotically active compounds or osmoprotectants (Pilon-Smits et al. 1999; Chen et al. 2000; Sawahel 2004; Le and McQueen-Mason 2006; Galvani 2007; Parida et al. 2008; Hossain and Fujita 2010; Li et al. 2011; Ranganayakulu et al. 2013; Shahbaz et al. 2013; Talat et al. 2013; Filippou et al. 2014). Osmoprotectants or compatible solutes are small molecules having low molecular weight, electrically neutral, highly soluble and non-toxic at molar concentrations (Ahn et al. 2011). They help plants to survive in extreme osmotic environment (Lang 2007). At the same time, these compatible solutes or osmoprotectants can stabilize proteins and membranes, and also reduce the osmotic potential of membranes to prevent dehydration inside the cell (Wani et al. 2013). They mount up inside the cell and maintain balance of osmotic difference between the cell’s surroundings and the cytosol. Osmoprotectants provide adaptation under various unfavourable environmental conditions such as excess salinity and temperature by raising the osmotic pressure in the cytoplasm (Jagesh et al. 2010; Ranganayakulu et al. 2013). They are collectively known as osmoprotectants and may be characterized in three parts. Osmoprotectants containig ammonium compounds (polyamines, glycinebetaine, b-alanine betaine, dimethyl-sulfonio propionate and choline-O-sulfate), osmoprotectants containing sugars and sugar alcohols (trehalose, fructan, mannitol and d-ononitol and sorbitol) and osmoprotectants containing amino acids (proline and ectoine). The manipulation in osmoprotectants responsive genes is one of the strategies to improve stress tolerance in plants by enhancing their production (Reguera et al. 2012; Jain 2013). In this review, we discuss mechanisms how osmoprotectants or osmolytes are involved in plant defence mechanisms under salinity and drought stress conditions. We also examine recent advances in putative mechanisms whereby osmoprotectants help plants to deal with environmental fluctuations. Finally, we have discussed apparent contradictions between some deleterious effects of both the stresses on plant development and roles of osmoprotectants in response to these abiotic stresses.
2 Sources of salinity and drought stresses
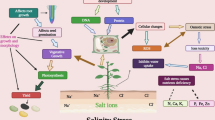
Salinity is introduced in the environment by natural as well as anthropogenic activities such as geogenic process, improper irrigation practices, industrial discharges, organic wastes, combustion of fuels and power generation. In natural phenomenon, weathering of parental rocks releases various kinds of salts particularly chlorides of sodium, calcium and magnesium and in the least amount of sulphate and carbonate are also released in the environment (Szabolcs 1989; Le and McQueen-Mason 2006; Galvani 2007; Munns and Tester 2008). The industrial discharge of drainage water used in manufacturing of dyes, paints and pigments, and leaching effect caused by improper sanitary landfills carried a huge amount of salt, which exerts pressure on hydrosphere (Zayed and Terry 2003; Rubio et al. 2010). Moreover, improper handling of municipal wastes, current agricultural practices such as excessive use of various pesticides and chemical fertilizers are considered one of the foremost causes of salinity (Villa-Castorena et al. 2003; Chen et al. 2010). At present, most of agricultural fields are severely suffering from salinity problem, and has become an important environmental concern globally due to its potential negative impacts on human and animal health. Similar to this, drought stress is due to manmade activities, can directly trigger a drought unlike other natural hazards, with exacerbating factors such as over farming, excessive improper irrigation practices, deforestation, over-exploitation of ground water and erosion, which severely inhibit the water holding capacity of land (Mishra and Singh 2010) (Fig. 1).

General scheme of salt and drought stress tolerance in plants. Some osmolytes involved in salt and drought stress tolerance through osmoprotection and reactive oxygen species (ROS) detoxification. They protect plant from osmotic and ionic stresses
3 Effects of salinity stress on plants
Area of salt affected land is near about >900 million ha at the global level or about 20 % of the world’s cultivated area, and surprisingly this area is still increasing. To date, it has been estimated that approximately half of the land dedicated to agricultural field is severely affected by salinity (Galvani 2007) or under risk of salinity due to the unmanaged irrigation handling. Salinity affects almost every aspect of physiology and biochemistry of plants and thus, significantly reduces crop yield and productivity. Plants respond to salinity in two ways, firstly higher concentration of Na+ in root causes osmotic stress, which decreases water potential (Fig. 6), and secondly impairment of nutrient balance causes ionic stress (Munns et al. 2006). Elevated concentrations of Na+ and Cl− in the soil solution may limit availability of essential nutrients by generating the most extreme ratios of Na+/Ca2+, Na+/K+, Ca2+/Mg2+, and Cl−/NO3 − (Munns et al. 2006). The enhanced level of salinity in plants has been reported to damage various physiological processes including seed germination, seedling growth, and flowering and fruiting due to induction of Na+ and Cl− toxicity, plasmolysis, and the up-regulation of levels of reactive oxygen species (ROS) (Jagesh et al. 2010). Extensive works have been carried out in response to salinity in various plant species (Jagesh et al. 2010; Shahbaz et al. 2013; Talat et al. 2013) (Table 1).
3.1 Direct effects of salinity stress on plants
Salinity is the foremost environmental abiotic stress factor causing stagnant plant growth and productivity (Galvani 2007; Chen et al. 2010; Ranganayakulu et al. 2013; Shahbaz et al. 2013). The injurious effects of excessive salinity on plants can be observed at both biochemical and cellular levels but most of observations have been focused on growth as the response to the toxicant. The excess accumulation of Na+ in the cell may lead to the drastic changes in different physiological processes and hence decline growth and plant productivity (Kaya et al. 2013). Aldesuquy et al. (2014) have reported 25 % decrease in leaf area, pigment content, Hill activity and photosynthetic efficiency of wheat plants after irrigation with sea water. Translocation of salts from roots to shoots is the product of transpirational flux, and is prerequisite to maintain water level of plants (Flowers and Yeo 1992). Higher accumulations of salt in the cell diminish the photosynthetic efficiency of plants due to inhibition of photosystem II (PSII) activity (Kalaji et al. 2011). The PSII has been considered the major site of salinity stress-mediated damage to electron transport chain activity (Mehta et al. 2010). Such inhibition in photosynthetic process is also discussed by Aldesuquy et al. (2014) in terms of reduced leaf area, photosynthetic pigments content, Hill reaction and 14CO2 fixation, and altered morphology of chloroplasts, number of reaction centres, net CO2 assimilation rate and Rubisco activity. Furthermore, study showed that salinity stress declined biomass accumulation in eggplant due to down regulation of photosynthetic gas exchange rate, net photosynthetic rate, transpiration rate, stomatal conductance, water utilization efficiency, quantum efficiency of PSII photochemistry and photochemical quenching, and up regulation of non-photochemical quenching (Wu et al. 2012). It has been reported that NaCl (5–30 mM) causes inhibition in growth, chlorosis, necrosis and finally death in Rosa chinensis due to higher accumulation of Cl− than Na+ in mature leaves than younger one (Wahomea et al. 2001). Salinity may also cause conflict in nutrient homeostasis by competing with the uptake of other essential nutrients. Salinity causes Na+ toxicity which can ultimately disrupts K+ uptake by root cells, and exerts severe toxic effects on enzymes at the genetic level and thus, causing impairment in normal plant metabolism (Conde et al. 2011). Shahbaz et al. (2013) have observed decrease in K+ and Ca2+ contents due to higher accumulation of Na+ in cells of eggplant. Moreover, there are some another important cellular enzymes subjected to salinity stress. Evidences show that enzymes participated in nitrogen metabolism and synthesis of amino acids were down-regulated while activities of hydrolyzing enzymes such as RNase, DNase, protease and few others were up-regulated under salinity (Nathawat et al. 2005; Siddiqui et al. 2008).
3.2 Indirect effects of salinity stress on plants
Besides direct impact of salinity stress on plants, a common effect of salinity stress is induction of excessive accumulation of ROS such as singlet oxygen (1O2), superoxide radical (O ·−2 ), H2O2 and the most dangerous ·OH radical, which cause damage to lipids, proteins and nucleic acids leading to the death of plant. Chloroplasts, mitochondria and plasma membranes are linked to the electron transport system and these are key sites for generation of ROS as byproducts of oxygen metabolism (Gill and Tuteja 2010a). In chloroplasts, photosystem I and II (PSI and PSII) are specific sites for generation of ROS. In mitochondria, complex I, ubiquinone and complex III of electron transport chain (ETC) are more susceptible for generation of O ·−2 . For an instance, when cells have experienced any kind of stress, their redox balance get disturbed. During redox reactions or ETC, electrons are coming out from oxygen evolving complex and get directly bind with O2 molecules and thus leading formation of ROS (Gill and Tuteja 2010a; Chen and Murata 2011) (Fig. 5). Therefore, regulation of ROS level is a vital process to avoid unwanted cellular cytotoxicity and oxidative damage (Halliwell and Gutteridge 1989).
Theerakulpisut and Gunnula (2012) have observed a significant rise in H2O2, lipid peroxidation in terms of MDA formation and other toxic oxygen derivatives in salt treated rice seedlings. Furthermore, Kaya et al. (2013) have reported NaCl-mediated generation of H2O2 which causes tissue damage and reduction in the growth of maize plant. Thus, understanding of associations between responses and downstream events of plant system that constitute successful adjustment to its transformed environment i.e. saline condition is one of the great challenges of plant biology.
4 Effects of drought stress on plants
Drought occurs in almost all climatic zones of the world including high as well as low rainfall areas. It is predominantly associated with the reduction in the precipitation amount received over an extended period of time. Drought is highly regulated by climatic, edaphic and agronomic factors including temperature, low relative humidity, high winds and distribution of rainy days during crop growing seasons including intensity and duration of rain. Soil aridity and soil dryness are severe limiting factors often faced by plants, which cause serious reduction in crop yield. Plants experience drought stresses in two ways first one is when water supply to roots becomes complicated and second one is, when the transpiration rate exceeds to the threshold limit (Le and McQueen-Mason 2006; Anjum et al. 2011) (Table 1).
4.1 Direct effects of drought stress on plants
Evidences showed that during the development of drought stress contained by a plant, all the key processes such as biosynthesis of photosynthetic pigments, net photosynthesis, effective quantum yield of PSII, electron transport rates, protein synthesis, energy and lipid metabolism are ceased (Liu et al. 2011; Deeba et al. 2012; Filippou et al. 2014). Similar to salinity stress, drought has also been responsible for impairment of water balance, mineral nutrients, membrane permeability and limits activities of enzymes. Study showed that the accumulation of abscisic acid (ABA) in plants during drought stress helps in signal transduction regulated by phosphorylation/dephosphorylation and controls anion and K+ transport system in guard cells (Osakabe et al. 2014). In response to drought stress, plants adapt various changes in their physico-biochemical processes such as alterations in plant morphology, growth rate, osmotic potential and improvement in defence mechanisms (Duan et al. 2007). Ajithkumar and Panneerselvam (2013) have reported a decline in growth and accumulation of osmoprotectants in Setaria italic under severe drought stress.
4.2 Indirect effects of drought stress on plants
Indirect effects of drought stress include induction of excessive accumulation of ROS, which cause damage to macromolecules. Kadioglu et al. (2011) have reported that level of O ·−2 , H2O2 and MDA significantly increased in Ctenanthe setose subjected to drought stress. Furthermore, Kubis et al. (2014) have reported that water shortage in cucumber cells causes an increase of membrane injury, lipoxygenase activity and lipid peroxidation leading to a reduction in growth.
To mitigate ROS-mediated damages, plants adapt various strategies and one of them is the production of enzymatic as well as non enzymatic (low molecular weight) antioxidants. The enzymatic antioxidants include superoxide dismutases (SOD), peroxidase (POD), ascorbate peroxidase (APX), guaiacol peroxidase (GPX), glutathione-s-transferase (GST) and catalase (CAT), which directly detoxify ROS (Vardharajula et al. 2011; Yadav et al. 2014). Non-enzymatic antioxidants such as ascorbate (ASC), glutathione (GSH), α-tocopherol, carotenoids and flavonoids also detoxify ROS and regulate many essential enzymatic reactions (Gill and Tuteja 2010a; Anjum et al. 2011). Moreover, several other enzymes like glutathione reductase (GR), monodehydroascorbate reductase (MDHAR) and dehydroascorbate reductase (DHAR) act in the ascorbate–glutathione cycle for quenching of toxic oxygen derivatives (Gill and Tuteja 2010a; Kadioglu et al. 2011).
5 Plant osmotic adjustment strategies against salinity and drought stresses
In response to salinity and drought stresses, plants develop several mechanisms, which involve changes at morphological, physiological and molecular levels. In defence mechanisms, plant enhances the production of osmoprotectants or osmolytes and regulates the nutrient homeostasis at cellular level against salinity and drought stresses. Osmoprotectants are probably universal and regulate the cellular osmotic adjustment, mitigate damaging risk caused by ROS, prevent membrane injury and stabilize proteins and enzymes (Le and McQueen-Mason 2006; Galvani 2007; Bohnert and Jensen 1996; Ashraf and Foolad 2007) (Table 3). They comprise proline, sucrose, polyols, trehalose as well as quaternary ammonium compounds (QACs) including glycine betaine, alanine betaine, proline betaine, choline-O-sulfate, hydroxyprolinebetaine, and pipecolatebetaine (Rhodes and Hanson 1993) (Table 3). Osmoprotectants preserve the cellular apparatus from the injury caused by dehydration, and at the same time they do not interfere with the normal metabolic processes at cellular level.
5.1 Groups of osmoprotectants
5.1.1 Ammonium compound group
5.1.1.1 Polyamines
Polyamines (PAs) are low molecular weight aliphatic nitrogen containing compounds, which are positively charged at physiological level and regulate the pH of cellular components (Groppa and Benavides 2008) (Table 3). PAs have great affinity to bind with DNA, RNA and proteins through electrostatic linkages (Kusano et al. 2007). Moreover, phytohormones like putrescine (Put), spermidine (Spd) and spermine (Spm) containing aliphatic amine compounds with aliphatic nitrogen structure have been reported in bacteria, plants as well as animals (Gill and Tuteja 2010b). In plants, research showed that on one hand Spm occurs only in angiosperms while on the other hand, thermospermine is likely to exist in the entire plant kingdom (Minguet et al. 2008).
PAs, belonging to biogenic amines group are widely distributed in plants and regulate diverse cellular functions essential for the cell growth. In plants, works on PAs are so heterogeneous despite they perform a major role in different vital physiological and biochemical events including senescence, development, cell proliferation, signal transduction, and also regulate expression of genes in response to various stresses in eukaryotic as well as in prokaryotic cells (Wang et al. 2003; Alcázar et al. 2006; Kusano et al. 2007) (Table 3). Studies show that plants tend to accumulate a huge amount of PAs against varied biotic and abiotic stresses (Alcázar et al. 2006; Hussain et al. 2011) (Table 2). Among a wide range of compatible solutes, PAs have great potential to reduce the risk of damage caused by extreme environmental stress (Liu et al. 2007; Hussain et al. 2011). Furthermore, many studies show that exogenous application of PAs enhances plant growth and provides protection against drought (Nayyar and Chander 2004; Yamaguchi et al. 2007; Kubis et al. 2014) and salinity (Duan et al. 2008; Kuznetsov and Shevyakova 2010) (Table 2).
In plants, levels of PAs are regulated by anabolic and catabolic processes, as well as by their conjugation to hydroxycinnamic acids. The PAs synthesis initiated with decarboxylation of ornithine or arginine, which is catalyzed by ornithine or arginine decarboxylases enzymes (ODC, ADC) for generation of diamine Put (Fig. 2). Put biosynthesis takes place in ADC pathway through three enzymatic steps in sequential action of ADC, agmatine iminohydrolase (AIH) and N-carbamoylputrescine amidohydrolase (CPA). Although osmoprotectants have low molecular weight, however, some higher molecular weight osmoprotectants (PAs) such as Spd and Spm are also produced via chronological addition of aminopropyl group to Put and Spd, respectively from the activities of spermidine synthase and spermine synthase. Although, methionine initiates the PAs metabolic pathway, however, it does not play any role in Spd or Spm synthesis but it provides an alternative pathway for the synthesis of SMM-induced PAs (Gill and Tuteja 2010b). Furthermore, aminopropyl group is generated from S-adenosylmethionine (SAM) via the action of SAM decarboxylase enzyme (dcSAM). The SAM is also a precursor of ethylene, which is produced from SAM via 1-aminocyclopropane-1-carboxylic-acid (ACC) by the activities of ACC synthase and ACC oxidase (Alcázar et al. 2006) (Fig. 2). It is reported that different sets of genes such as SPDS1 and SPDS2 play an important role in Spd biosynthesis, which is further required in PAs synthesis (Alcázar et al. 2006). In a similar way, degradation of PAs are also take place via oxidative deamination process catalyzed by amine oxidases, specifically diamine oxidases (DAO) and polyamine oxidases (PAO) enzymes (Alcázar et al. 2006; Gill and Tuteja 2010b) (Fig. 2). Diamine oxidases enzymes (DAOs) have a great affinity to diamines, whereas polyamine oxidases enzymes (PAOs) cause oxidation of secondary amine group of Spd and Spm (Fig. 2). The Put catabolization takes place via the action of diamine oxidases (DAOs) enzymes (Gill and Tuteja 2010b). In this process, Put converts into Δ1-pyrroline and generates ammonia and H2O2 as a by product. Further, in oxidation of Put, Δ1-pyrroline is catabolized into γ-aminobutyric acid (GABA) which is then converted into succinic acid, a component of the Krebs’ cycle (Eller et al. 2006; Gill and Tuteja 2010b).

Polyamine biosynthetic, breakdown pathways and related metabolites in higher plants. ACC 1-amino-cyclopropane-1-carboxylic-acid, ADC arginine decarboxylase, AIH agmatine iminohydrolase, CPA N-carbamoylputrescine amidohydrolase, DAO diamine oxidase, dcSAM decarboxylated S-adenosylmethionine, GABA c-aminobutyric acid, ODC ornithine decarboxylase, PAO polyamine oxidase, SAM sadenosylmethionine, SAMDC S-adenosylmethionine decarboxylase, SPDS spermidine synthase, SPMS spermine synthase, TPMS thermospermine synthase
5.1.1.2 Betaines
Betaines, belonging to quaternary ammonium compounds group including glycine betaines (GB), b-alaninebetaine, prolinebetaine, choline-O-sulphate, dimethyl sulphoniopropionate, hydroxyproline betaine, and pipecolate betaine act as efficient compatible solutes (Ashraf and Harris 2004; Flowers and Colmer 2008; Vinocur and Altman 2005; Koyro et al. 2012) (Tables 2, 3). Among them, GB is widely distributed and acts as an key osmoprotectant which have the highest potential to mitigate negative impact caused by a variety of abiotic stresses in numerous organisms including bacteria, cyanobacteria, algae, fungi, animals and some higher plant families such as Chenopodiaceae and Gramineae (Türkan and Demiral 2009) (Table 2). In response to various environmental stresses particularly drought and salinity, GB accumulates in chloroplasts of some plant species and also reported in diverse kind of microbes (Sakamoto and Murata 2002; Sawahel 2004; Ranganayakulu et al. 2013). At the same time, GB encourages water flow into cells for maintaining the intracellular osmotic equilibrium and regulates the cascade of signal transduction under varied stresses (Kumar et al. 2003; Ashraf and Foolad 2007; Ranganayakulu et al. 2013). The fundamental role of osmoprotectants is to maintain turgescent cells and regulate the water potential in order to ensure impartial water relation (Wang et al. 2003). One of the foremost constructive role of GB is that it renders adaptation in mediating the osmotic adjustment via Na+/K+ discrimination, inducing defence enzymes and membrane consistency, which considerably contribute tolerance in plants against salinity (Ashraf and Foolad 2007). Extensive research has been focused on accumulation of betaines inherently in various crop plants including rice, groundnut and some other higher plants against salinity and drought stresses (Jagendorf and Takabe 2001; Sawahel 2004; Ranganayakulu et al. 2013). In a same manner, exogenous applications of GB enhance its level internally and may promote plant growth and crop productivity under various abiotic stresses (Table 2).
In nature, the biosynthesis of GB in animals, microorganisms and plants systems takes place via two distinct pathways with assistance of two different substrates namely choline and glycine (Sakamoto and Murata 2002; Wani et al. 2013) (Fig. 3). In glycine-betaine biosynthetic pathway, GB is formed during two successive oxidation processes of choline via choline monooxygenase (CMO) and betaine aldehyde dehydrogenase, and whole reactions are performed in chloroplasts specially in stroma and catalyzed by CMO and NAD+ dependent betaine aldehyde dehydrogenase (BADH) enzymes (Wani et al. 2013). In recent year, it has been reported that the Escherichia coli and some animals are capable to synthesize GB from activities of dehydrogenases including membrane attached CDH and BADH (Ahmad et al. 2013; Wani et al. 2013) (Fig. 3a). The GB biosynthesis pathway has been familiar in extremely halophilic microbes such as Ectothiorhodospira halochloris and Actinopolyspora halophilia (Ahmad et al. 2013; Wani et al. 2013). In GB biosynthesis, three consecutive methylations of amino residue take place with the help of two enzymes namely; glycine sarcosine methyltransferase (GSMT) and sarcosine dimethylglycine methyltransferase (SDMT), with S-adenosylmethionine serve as a donor of methyl group (Nyyssola et al. 2000). Both enzymes: GSMT and SDMT have higher affinity to their substrates and also exist great potential to complete all required processes for methylation during the GB biosynthesis (Nyyssola et al. 2000; Chen and Murata 2002) (Fig. 3b).

a Biosynthetic pathway of glycine betaine in higher plants. Choline is converted to betaine aldehyde, by choline monooxygenase, which is then converted to glycine betaine by betaine aldehyde dehydrogenase. b Biosynthesis of glycine betaine from glycine in halophilic micro-organisms such as, Ectothiorhodospira halochloris and Actinopolyspora halophilia (Modified from Sakamoto and Murata 2002)
5.1.2 Sugars and sugar alcohols
5.1.2.1 Carbohydrate sugars
Sugars provide carbon and energy for normal functioning of cellular metabolism and regulate growth and development of plants. Roles of sugars and sugar alcohols have been widely accepted as osmoprotectants, which regulate the osmotic adjustment, provide membrane protection and scavenge toxic ROS against various kind of stresses (Murakeozy et al. 2003; Ahmad and Sharma 2008; Livingston et al. 2009; Van den Ende and Valluru 2009; Koyro et al. 2012). Evidences show that higher accumulation of reduced form of sugars such as glucose, sucrose, fructose and fructans function as osmoprotectants under salinity and drought stresses (Kerepesi and Galiba 2000; Murakeozy et al. 2003) (Table 2). Fructan may be treated as reserve carbohydrate source and accumulates in tubers or taproots of plants. Pilon-Smits et al. (1995) reported that fructan protects plant against severe drought stress. In a same way, trehalose is an oxidized form of sugar exists in some microbes including bacteria and fungi as well as some desiccation-tolerant plants (Vinocur and Altman 2005). Like other reduced form of sugar, trehalose also acts as an osmoprotactant or osmolyte and protects membranes, proteins and decreases aggregation of denatured proteins (Ashraf and Harris 2004; Koyro et al. 2012). Various sugars are shown in Table 2, which involve in mitigation of the negative impact of abiotic stresses.
5.1.2.2 Sugar alcohols
Sugar alcohols, sometime also called polyols may be defined in two ways on the basis of their structure. The first one is cyclic structure consisting of myoinositol and pinitol, and another one is linear structure including sorbitol, mannitol, xylitol and ribitol. Polyols originate via the reduction of aldoses or their phosphate esters and are generally water soluble in nature (Tari et al. 2010). Sorbitol, a kind of polyol having a six alcoholic carbon sugar is widely distributed in plants. It originates in parallel with sucrose during the photosynthesis and translocates the carbon skeleton and energy between sources and sinks (Jain et al. 2010). Williamson et al. (2002) have reported that sugar alcohols including mannitol, sorbitol and inositol enhance tolerance potential of plants against drought and salinity. Plants synthesize a great amount of mannitol (Mitoi et al. 2009) for quenching the most toxic hydroxyl radical (·OH) generated via the fenton reaction (Gill and Tuteja 2010a) (Fig. 5). Polyols regulate the osmotic balance, facilitate water adjustment in the cytoplasm (Table 3), enable Na+ sequestration into the vacuole or apoplast and thus, provide protection to cellular structures by interacting with membrane protein complexes and enzymes against drought (Li et al. 2011) and salinity stress (Kanayama et al. 2006). Several studies show that sugar molecules improve plant growth and may reduce risk of damage caused by varied abiotic stresses (Table 2).
In between carbohydrates, sugars such as glucose and fructose contain wide range of aldehyde or ketone groups. In a non reducing sugar, such as sucrose, the ketone or aldehyde group is reduced to an alcohol or combined with a similar group on another sugar. Fructans biosynthesis takes place in the vacuole system via the activity of fructosyltransferases enzyme in plants, and in some cereals which have bulk amount of β-2,6-linked fructosyl units with short β-2,1-linked branches (Chalmers et al. 2005). The β-2,6-linked fructan is synthesized via successive co-action of sucrose:sucrose 1-fructosyltransferase (1-SST) and sucrose:fructan 6-fructosyltransferase (6-SFT) (Livingston et al., 2009). The enzyme responsible for synthesis of the β-2,1-linked branch in graminan is fructan:fructan 1-fructosyltransferase (1-FFT) (Kawakami and Yoshida 2005; Garcia et al. 2011). In most of the cases, sucrose biosynthesis is regulated by the interconversion of fructose-6-phosphate and fructose-1,6-bisphosphate, and this phenomenon is totally based on three distinct enzymes activities viz. fructose-1,6-bisphosphatase which catalyses a reaction in sucrose biosynthesis, phosphofructokinase (ATP-dependent), which catalyses a reaction to glycolysis, and pyrophosphate:fructose-6-phosphate 1-phosphotransferase (PFP) which catalyses a reaction in both directions using pyrophosphate (PPi) as a phosphoryl donor and these PPis come out as a result of interconversion of glucose-1-phosphate to UDP-glucose (Hare et al. 1998). Similarly, sugar alchohal such as, trehalose is synthesized in a two-reaction process, in which trehalose-6-phosphate (T6P) is first synthesized from glucose-6-P as a acceptor and UDP-glucose as a glucosyl donor, and these reactions are catalyzed by the enzyme trehalose phosphate synthase (TPS) and subsequently de-phosphorylated by trehalose-6P phosphatase (TPP) (Vandesteenea et al. 2010). Further, hydrolysis of trehalose is catalyzed via the action of trehalase (TRE) enzyme (Avonce et al. 2006; Vandesteene et al. 2010).
Mannitol, also belongs to sugar alcohol groups, originates via the action of mannose-6-phosphate reductase. In this way, mixture of glucose/fructose is got interchanged into gluconic acid and mannitol via simultaneous activities of two enzymes (Nidetzky et al. 1996). The first one is the NADH-dependent mannitol dehydrogenase (MDH) and the second one is NAD-dependent glucose dehydrogenase. Similarly, in the sorbitol biosynthetic process, it shares ordinary hexose phosphate group with sucrose production in the cytosol. Kanayama et al. (1992) and Cheng et al. (2005) have reported that glucose-6-phosphate interchanged with sorbitol-6-phosphate via the activity of sorbitol-6-phosphate dehydrogenase (S6PDH or aldose-6-phosphate reductase). Zhou et al. (2003) have also used sorbitol-6-phosphate for the production of sorbitol via the dephosphorylation of sorbitol-6-phosphatase. Moreover, numerous study have revealed that synthesis of sugar alcohols enhanced via over expression of different sets of genes. For instance, trehalose is synthesized by otsA (trehalose-6-phosphate synthase), otsB (trehalose-6-phosphate phosphatase), and AtTPS1 and AtTPS2 (trehalose-6-phosphate synthase (Garg et al. 2002; Baea et al. 2005), mannitol, sorbitol and ononitol via action of mt1D (d-mannitol-1-phosphate dehydrogenase), S6PDH (sorbitol-6-phosphate dehydrogenase), OemaT1 (mannitol-1) and imt1 (d-myo-inositolmethyltransferase) under stress conditions (Sheveleva et al. 1997; Abebe et al. 2003; Gao et al. 2001).
5.1.3 Amino acids
5.1.3.1 Proline
Proline (Pro) is the most important osmolyte and signalling molecule which accumulates generally in the cytosol and also contributes in protection of membranes, proteins and enzymes against various stresses (Tables 2, 3). At the same time, Pro regulates different metabolic processes such as stabilizing sub-cellular structures including membranes and proteins, scavenges toxic oxygen derivatives and buffering the cellular redox potential under different stress conditions (Sharma and Dietz 2006; Hoque et al. 2008) (Table 3). It may alleviate the cytoplasmic acidosis at that extent which has necessary for maintaining homeostasis between the NADP+ and NADPH required for normal metabolism (Hoque et al. 2008). Similar to Pro, ectoine is a compatible solute that is synthesized de novo by many halotolerant bacteria (Bernard et al. 1993) and acts as a potent osmoprotectant in some bacteria. Proline accumulation in the cytoplasm facilitates water uptake and also reduces levels of toxic ions under stress condition (Ashraf and Foolad 2007). Filippou et al. (2014) have reported higher accumulation of Pro in A. altissima seedlings under drought and salinity stresses, suggesting its protective role. Furthermore, Pro provides protection by improving the redox homeostasis (Hoque et al. 2008) and quenching of toxic free radicals under stress circumstances (Sharma and Dietz 2006). In process of adaptation, plants firstly enhance level of amino acids along with organic acids and ammonium compounds, collectively treated as cytoplasmic solutes, which regulate the osmotic adjustment against stress circumstances. Several researchers showed that Pro alleviates negative impact on growth and development of crop and vegetable plants caused by different abiotic stresses (Sharma and Dietz 2006; Hoque et al. 2008) (Table 2). For instance, under salinity Pro reduces the accumulation of Na+ and Cl− (Khattab and Afifi 2009), and in drought stress it facilitates water availability to the plant cell (Ashraf and Foolad 2007). Many researchers reported that the exogenous application of Pro enhances its endogenous level and therefore, promoting growth, antioxidant defence system and also decreases rate of uptake of Na+ and Cl− (Nounjana et al. 2012; Ranganayakulu et al. 2013; Shahbaz et al. 2013) (Table 2).
In the plant system, the biosynthesis of Pro occurs in the cytosol and plastid while its degradation takes place in mitochondria (Ashraf and Foolad 2007). Its biosynthesis also depends on up-regulation of Pro biosynthesis genes such as pyrroline-5-carboxylatesynthetase (P5CS) and pyrroline-5-carboxylate reductase (P5CR) (Nounjana et al. 2012). In the Pro biosynthesis, there are two different precursors i.e. glutamate and ornithin (Orn). The conversion of glutamate to Pro takes place by two consecutive steps, first is catalyzed by P5CS, which is bifunctional enzyme. It catalyzes activation of glutamate by phosphorylation and second is P5CR activity, which reduces labile intermediate c-glutamyl phosphate into glutamate semialdehyde (GSA), both are involved in Pro biosynthesis and catabolism (Hu et al. 1992; Hare and Cress 1997; Verbruggen and Hermans 2008) (Fig. 4). In a same way, ornithine enzyme occurs in mitochondria can be transmitted to P5C through the action of Orn-d-aminotransferase (OAT) (Verbruggen and Hermans 2008). In contrast to this, degradation of Pro is caused by the reverse action of proline dehydrogenase (PDH) and P5C dehydrogenase (P5CDH) (Fig. 4). Moreover, another pathway of Pro biosynthesis is glutathione via the action of glutamic-g-semialdehyde (GSA) and D1-pyrroline-5-carboxylate (P5C). The P5C synthase (P5CS) enzymes catalyses conversion of glutathione to P5C, followed by the action of P5C reductase (P5CR) enzyme, which reduces the P5C to Pro (Ashraf and Foolad 2007).

Biosynthetic pathway of proline through glulamate and ornathine in plants (adopted from Ahmad and Sharma 2008). P5CS pyrroline-5-carboxilate synthetase, P5CR pyrroline-5-carboxilate synthetase, ProDH proline dehydrogenase
6 Tolerance mechanism of osmoprotectants against salt and drought stresses
Plants produce a wide range of organic solutes or osmoprotectants belonging to distinguished groups such as ammonium compounds, sugar and sugar alcohols, and amino acids group (Rhodes and Hanson 1993; Koyro et al. 2012). The elevated concentrations of these osmoprotectants in plant system may provide protection to cellular organs by increasing water uptake potential and facilitating the enzyme activity (Wang et al. 2003; Attipali et al. 2004; Ashraf and Foolad 2007; Kusano et al. 2007; Koyro et al. 2012; Ranganayakulu et al. 2013). These osmoprotectants detoxify adverse impact of salinity and drought in plants via adopting two different mechanisms first is the improvement of antioxidant defence system (Fig. 5) and another one is sustainability in ion homeostasis (Fig. 6).

During abiotic stress conditions changes in the photosynthetic electron transport lead to the formation of superoxide radicals because the molecular oxygen competes with NADPH for reduction at the acceptor side of photosystem I. Also, damages in proteins of photosystem II (PS II) are associated with the decrease of PSII chemistry caused by ROS. At the same time, different osmolytes accumulate under salt and drought stress conditions and they detoxify the adverse effects of stress by increasing enzymatic and non-enzymatic antioxidants and also they maintain water and ion movement

The schematic arrangement of a plant cell includes three compartments that are defined by the extracellular space; cytosolic space and vacoular space. Indicated are the osmolytes and ions compartmentalized in the cytoplasm and vacuole, and transport proteins responsible for Na+ and Cl− homeostasis across the plasma membrane and tonoplast. Similarly, under drought stress ABA plays important role in ion homeostasis and water balance through stomata closing and decreases in water potential (Ψ)
6.1 Antioxidant defence systems
In response to severe oxidative stress caused by salinity and drought, plants trigger a complex enzymatic antioxidant enzymes including superoxide dismutase (SOD), peroxidase (POD), catalase (CAT) and ascorbate peroxidase (APX) and some other non-enzymatic low molecular weight antioxidants like glutathione (GSH), ascorbate (ASC) and carotenoids which provide protection via quenching of toxic ROS (Gill and Tuteja 2010a; Hossain and Fujita 2010; Kadioglu et al. 2011; Vardharajula et al. 2011; Kaya et al. 2013; Kubis et al. 2014). In a series of detoxifying mechanisms, plants enhanced the production of metalloenzyme SOD which is responsible for the conversion of O ·−2 to H2O2 and further CAT and PODs catalyze breakdown of H2O2 (Gill and Tuteja 2010a). Although CAT is apparently absent in the chloroplast, however, H2O2 can be detoxified in a reaction catalyzed by an ascorbate specific peroxidase often present in high levels in this organelle through the ascorbate–glutathione cycle. Ascorbate can also be oxidized via direct reaction with O ·−2 or by serving as a reductant of a-chromoxyl radical of oxidized α-tocopherol (Gill and Tuteja 2010a). Recently, Vardharajula et al. (2011) and Wu et al. (2012) have also reported enhanced activities of diverse range of antioxidant enzymes including SOD, POD, CAT and APX in Solanum melongena seedlings and maize plant under salinity and drought stresses.
A number of studies showed that under various stress circumstances, osmoprotectants such as PAs, GB, sugar and alchohal sugar and Pro, up-regulate antioxidant enzymes activities to reduce adverse effect of oxidative stress (Ashraf and Foolad 2007; Kubis 2008; Koyro et al. 2012). Filippou et al. (2014) have also reported that Pro efficiently removes toxic ROS and regulates the osmotic balance between cells and their surroundings under salinity and drought. In a same way, reports showed that exogenous application of mannitol, sorbitol and trehalose, PAs and glycine betaine improve plant growth in maize (Kaya et al. 2013), rice (Theerakulpisut and Gunnula 2012) brinjal (Wei et al. 2009) and mung bean (Hossain and Fujita 2010), respectively by improving antioxidant defence system against salinity stress. These osmoprotectants avoid participation in any biochemical reaction and stored in the cytosol, may enhance the potential of defence enzymes and resolve water crisis during osmotic stress caused by salinity and drought (Kaya et al. 2013; Filippou et al. 2014) (Fig. 5). For instance, Verma and Mishra (2005) reported that PAs (Put) inverted salinity-induced reduction in seedling growth and biomass accumulation in Brassica juncea. Further, enhanced activities of antioxidant enzymes and carotenoids may protect Brassica juncea seedlings against salinity stress. The results obtained in last few years strongly argue that osmolytes could be very promising compounds for the reduction of abiotic stress sensitivity of crops, because under certain conditions they have been found to mitigate damaging effects of various stress factors in plants by enhancing antioxidant system (Table 2; Fig. 5).
6.2 Ion homeostasis
Ion homeostasis is one of the most important strategies, adapted by the plant for mitigating toxic impact caused by salinity and drought. Due to similar responses of both stresses, plants accumulate diverse groups of osmoprotectants, which provide osmotic adjustment and ion homeostasis via the ion exchange activity (Ranganayakulu et al. 2013). The most common effect of soil salinity is the reduction in growth and productivity due to direct specific ion toxicity like Na+ and Cl− toxicity that decrease the uptake of essential nutrients like phosphorus (P), potassium (K+), nitrogen (N), and calcium (Ca++) (Zhu 2001; Niu et al. 1995; Munns and Tester 2008; Parihar et al. 2014). These toxic ions exert negative impacts on intracellular K+ influx, attenuating acquisition of this essential nutrient by cells. Moreover, Na+ toxicity is associated with its competitive nature with K+ for binding essential sites, which carry normal cellular function. It has been reported that decrease in K+/Na+ ratio may result in K+ deficiency in the cell therefore, K+ homeostasis is an important factor in salinity tolerance (Munns and Tester 2008) (Fig. 6). In order to tolerate salinity, plants tend to store Na+ in vacuoles, which defend the cytosolic enzymes from toxic action (Apse et al. 1999). However, excess concentration of Na+ in the vacuolar system may enhance the osmotic pressure of other cellular compartments leading to damage of cellular organelles (Munns and Tester 2008). At the same time, ion homeostasis depends on transportation potential of transmembrane proteins (Osakabe et al. 2014), which regulate ion fluxes (influx and efflux) including H+ translocating ATPases, Ca++ ATPases and pyrophosphatases (Niu et al. 1995). The extrinsically bound Na+/H+ antiporters at the plasma membrane and tonoplast acquired energy from electrochemical H+ gradients, and couple passive movement of H+ to active movement of Na+ out of the cell and into the vacuole, respectively (Blumwald 1987) (Fig. 6). Accumulation of K+ into the vacuole against electrochemical gradient is essential to generate efficient turgor for stomatal opening, and this uphill K+ transport has to be mediated by secondary active carriers (Zhu 2001). Some compatible osmolytes such as PAs, Pro, sucrose, soluble carbohydrates, glycinebetaine, and other solutes accumulate in the cytosol and organelles to balance the osmotic pressure of ions in the vacuole thus maintaining the cell turgor (Munns and Tester 2008) and improving water uptake from drying soil. The accumulation of these compounds under salinity was linked to the maintenance of a high K+ concentration and thus a lower Na+/K+ ratio, with a better performance of the cell water status (Rivero et al. 2014). Moreover, Nounjana et al. (2012) have reported that exogenous application of Pro and trehalose suppressed Na+ uptake and accumulation, reduced Na+/K+ ratio as well as enhanced activities of antioxidants enzymes in rice plants under salinity.
Similar to salinity, drought stress also causes an impairment of ionic balance in plants. During drought stress, anions and cations, such as Cl− and K+, water transport systems in the plasma membrane and tonoplast induce turgor pressure changes in guard cells, which result into stomatal closure (Kim et al. 2010). Under water deficit, ABA inhibits H+ATPase activity by reducing H+ ATPase phosphorylation level, and this is important to maintain membrane depolarization (Hayashi et al. 2011; Osakabe et al. 2014). Shinozaki and Yamaguchi-Shinozaki (2007) have reported that H+ATPase induces negative electric potential gradient inside the plasma membrane which causes a hyper polarization of the plasma membrane and subsequent opening of voltage-regulated inward K+ channels, resulting in stomatal opening. Andrés et al. (2014) have reported that the vacuolar K+/H+ exchange is a critical component involved in vacuolar remodeling and the vacuolar pH regulation during stomatal movements. The ABA mainly produced in leaf vascular tissues in response to drought stress is transported to guard cells where it induces stomatal closure. The ABA, which is produced rapidly in response to drought and salinity, plays a critical role in the regulation of stress responses and induces a series of signalling cascades (Osakabe et al. 2014). Specifically, the accumulation of ABA promotes stomatal closure to minimize water loss, accelerates leaf senescence, down regulates plant growth, and induces the biosynthesis of protective substances under drought and salinity (Nishiyama et al. 2011). Similar to salinity, critical roles of osmolytes accumulation under drought stress has been investigated to understand tolerance of plants to dehydration (Wani et al. 2013). The ABA promotes an efflux of K+ ions from the guard cells, which results in loss of turgor pressure leading to stomata closure (Fig. 6). Nayyar and Walia (2003) have reported that accumulation and mobilization of Pro was associated with enhanced tolerance to drought stress. Exogenous GB application caused increased accumulation of endogenous osmolytes such as free Pro and GB; they maintain ion homeostasis under stress condition (Ashraf and foolad 2007; Zhang et al. 2014).
7 Conclusion and future perspectives
Environmental constrains such as salinity and drought are major factors limiting plant growth and productivity. Furthermore, ever increasing global population is compelling the scientists to develop more efficient strategies for augmenting crop production in order to ensure food security under such adverse conditions. To date, there are some excellent studies which bear significance of osmoprotectants in plants under salinity and drought. For instance, endogenous osmoprotectants level as well as introduction of gene(s) of their biosynthetic pathways appear to commence a wide range of biochemical pathways, which may enhance plant tolerance against salinity and drought. In a way of defence, plants have evolved adaptive mechanisms which can be harnessed as important resources for development of crops tolerant to extremities. Research on appreciating salinity and drought stress tolerance mechanisms of plants has been on the upfront using wide array of “omics” technologies, such as genomics, proteomics, and metabolomics, which are assisting scientists to identify the genetics behind plant tolerance mechanism. Investigation of the gene regulation and balance of individual stress tolerance mechanisms will aid in translating information to other salinity-sensitive crops. Such studies will be obliging for ensuring sustainability of future research efforts to get better crop performance on marginal and irrigated land through genetic manipulation. Considering significance of osmoprotectants in many plant functions at physiological and biochemical level under salinity and drought stresses, further research dealing with endogenous regulation of osmoprotectants metabolism may contribute much to deal with such adverse conditions.
References
Abebe T, Guenzi AC, Martin B, Cushman JC (2003) Tolerance of mannitol-accumulating transgenic wheat to water stress and salinity. Plant Physiol 131:1748–1755
Ahmad P, Sharma S (2008) Salt stress and phytobiochemical responses of plants. Plant Soil Environ 54:89–99
Ahmad R, Lim CJ, Kwon SY (2013) Glycine betaine: a versatile compound with great potential for gene pyramiding to improve crop plant performance against environmental stresses. Plant Biotechnol Rep 7:49–57
Ahn C, Park U, Park PB (2011) Increased salt and drought tolerance by d-ononitol production in transgenic Arabidopsis thaliana. Biochem Biophys Res Commun 415:669–674
Ajithkumar IP, Panneerselvam R (2013) Osmolyte accumulation, photosynthetic pigment and growth of Setaria italica (L.) P. Beauv. under drought stress. Asian Pac J Reprod 3:220–224
Alcázar R, Marco F, Cuevas JC, Patron M, Ferrando A, Carrasco P, Tiburcio AF, Altabella T (2006) Involvement of polyamines in plant response to abiotic stress. Biotechnol Lett 28:1867–1876
Aldesuquy H, Baka Z, Mickky B (2014) Kinetin and spermine mediated induction of salt tolerance in wheat plants: leaf area, photosynthesis and chloroplast ultrastructure of flag leaf at ear emergence. Egypt J Bas Appl Sci 1:77–87
Andrés Z, Pérez-Hormaeche J, Leidi EO, Schlücking K, Steinhorst L, McLachlan DH, Schumacher K, Hetherington AM, Kudla J, Cubero B, Pardo JM (2014) Control of vacuolar dynamics and regulation of stomatal aperture by tonoplast potassium uptake. Proc Natl Acad Sci USA 111:1806–1814
Anjum SA, Xie X, Wang LC, Saleem MF, Man C, Lei W (2011) Morphological, physiological and biochemical responses of plants to drought stress. Afr J Agric Res 6:2026–2032
Apse MP, Aharon GS, Snedden WA, Blumwald E (1999) Salt tolerance conferred by overexpression of a vacuolar Na+/H+ antiport in Arabidopsis. Science 285:1256–1258
Ashraf M, Foolad MR (2007) Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ Exp Bot 59:206–216
Ashraf M, Harris PJC (2004) Potential biochemical indicators of salinity tolerance in plants. Plant Sci 166:3–16
Attipali RR, Kolluru VC, Munusamy V (2004) Drought induced responses of photosynthesis and antioxidant metabolism in higher plants. J Plant Physiol 161:1189–1202
Avonce N, Mendoza-Vargas A, Morett E, Iturriaga G (2006) Insights on the evolution of trehalose biosynthesis. BMC Evol Biol 6:109
Baea H, Herman E, Bailey B, Bae HJ, Sicher R (2005) Exogenous trehalose alters Arabidopsis transcripts involved in cell wall modification, abiotic stress, nitrogen metabolism, and plant defense. Physiol Plant 125:114–126
Bernard T, Jebbar M, Rassouli Y, Himdi-Kabbab S, Hamelin J, Blanco C (1993) Ectoine accumulation and osmotic regulation in Brevibacterium linens. J Gen Microbiol 139:129–138
Blumwald E (1987) Tonoplast vesicles as a tool in the study of ion-transport at the plant vacuole. Physiol Plant 69:731–734
Bohnert HJ, Jensen RG (1996) Strategies for engineering water-stress tolerance in plants. Trends Biotechnol 14:89–97
Chalmers J, Lidgett A, Cummings N, Cao Y, Forster J, Spangenberg G (2005) Molecular genetics of fructan metabolism in perennial ryegrass. Plant Biotechnol J 3:459–474
Chen WP, Li PH, Chen THH (2000) Glycine betaine increases chilling tolerance and reduces chilling-induced lipid peroxidation in Zea mays. L.. Plant Cell Environ 23:609–618
Chen THH, Murata N (2002) Enhancement of tolerance of abiotic stress by metabolic engineering of betaines and other compatible solutes. Curr Opin Plant Biol 5:250–257
Chen THH, Murata N (2011) Glycinebetaine protects plants against abiotic stress, mechanisms and biotechnological applications. Plant Cell Environ 34:1–20
Chen W, Hou Z, Wu L, Liang Y, Wei C (2010) Effects of salinity and nitrogen on cotton growth in arid environment. Plant Soil 326:61–73
Cheng LL, Zhou R, Reidel EJ, Sharkey TD, Dandekar AM (2005) Antisense inhibition of sorbitol synthesis leads to up-regulation of starch synthesis without altering CO2 assimilation in apple leaves. Planta 220:767–776
Conde A, Silva P, Agasee A, Conde C, Gerós H (2011) Mannitol transport and mannitol dehydrogenase activities are coordinated in Olea japonica under salt and osmotic stress. Plant Cell Physiol 52:1766–1775
Deeba F, Pandey AK, Ranjan S, Mishra A, Singh R, Sharma YK, Shirke PA, Pandey V (2012) Physiological and proteomic responses of cotton (Gossypium herbaceum L.) to drought stress. Plant Physiol Biochem 53:6–18
Duan B, Yang Y, Lu Y, Korpelainen H, Berninger F, Li C (2007) Interactions between drought stress, ABA and genotypes in Picea asperata. J Exp Bot 58:3025–3036
Duan JJ, Li J, Guo SR, Kang YY (2008) Exogenous spermidine affects polyamine metabolism in salinity stressed Cucumis sativus roots and enhances short term salinity tolerance. J Plant Physiol 165:1620–1635
Eller MH, Warner AL, Knap HT (2006) Genomic organization and expression analyses of putrescine pathway genes in soybean. Plant Physiol Biochem 44:49–57
Faical B, Imen A, Kaouther F, Moez H, Habib K, Khaled M (2009) Physiological and molecular analyses of seedlings of two Tunisian durum wheat (Triticum turgidum L.) varieties showing contrasting tolerance to salt stress. Acta Physiol Plant 31:145–154
Filippou P, Bouchagier P, Skotti E, Fotopoulos V (2014) Proline and reactive oxygen/nitrogen species metabolism is involved in the tolerant response of the invasive plant species Ailanthus altissima to drought and salinity. Environ Exp Bot 97:1–10
Flowers TJ, Colmer TD (2008) Salinity tolerance in halophytes. New Phytol 179:945–963
Flowers TJ, Yeo AR (1992) Solute transport in plants. Blackie, Glasgow, p 176
Galvani A (2007) The challenge of the food sufficiency through salt tolerant crops. Rev Environ Sci Biotechnol 6:3–16
Gao M, Tao R, Miura K, Dandekar AM, Sugiura A (2001) Transformation of Japanese persimmon (Diospyros kaki Thunb.) with apple cDNA encoding NADP-dependent sorbitol-6-phosphate dehydrogenase. Plant Sci 160:837–845
Garcia PMA, Asega AF, Silva EA, Carvalho MAM (2011) Effect of drought and re-watering on fructan metabolism in Vernonia herbacea (Vell.) Rusby. Plant Physiol Biochem 49:664–670
Garg AK, Kim JK, Owens TG, Ranwala AP, Choi YD, Kochian LV, Wu RJ (2002) Trehalose accumulation in rice plants confers high tolerance levels to different abiotic stresses. Proc Natl Acad Sci USA 99:15898–15903
Gill SS, Tuteja N (2010a) Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol Biochem 48:909–930
Gill SS, Tuteja N (2010b) Polyamines and abiotic stress tolerance in plants. Plant Signal Behav 5:26–33
Gosal SS, Wani SH, Kang MS (2009) Biotechnology and drought tolerance. J Crop Improv 23:19–54
Groppa MD, Benavides MP (2008) Polyamines and abiotic stress: recent advances. Amino Acids 34:35–45
Halliwell B, Gutteridge JMC (1989) Free radicals in biology and medicine, 2nd edn. Clarendon, Oxford
Hare PD, Cress WA (1997) Metabolic implications of stress-induced proline accumulation in plants. Plant Growth Regul 21:79–102
Hare PD, Cress WA, Van Staden J (1998) Dissecting the roles of osmolyte accumulation during stress. Plant Cell Environ 21:535–553
Hayashi M, Inoue S, Takahashi K, Kinoshita T (2011) Immunohistochemical detection of blue light-induced phosphorylation of the plasma membrane H+ATPase in stomatal guard cells. Plant Cell Physiol 52:1238–1248
Hayat S, Hayat Q, Alyemeni MN, Ahmad A (2013) Proline enhances antioxidative enzyme activity, photosynthesis and yield of Cicer arietinum L. exposed to cadmium stress. Acta Bot Croat 2:323–335
Hoque MA, Banu MNA, Nakamura Y, Shimoishi Y, Murata Y (2008) Proline and glycine betaine enhance antioxidant defense and methyl glyoxal detoxification systems and reduce NaCl-induced damage in cultured tobacco cells. J Plant Physiol 165:813–824
Hossain MA, Fujita M (2010) Evidence for a role of exogenous glycine betaine and proline in antioxidant defense and methyl-glyoxal detoxification systems in mung bean seedlings under salt stress. Physiol Mol Biol Plants 16:19–29
Hu CA, Delauney AJ, Verma DPS (1992) A bifunctional D1-enzyme pyrroline-5-carboxylate synthetase catalyzes the first two steps in proline biosynthesis in plants. Proc Natl Acad Sci USA 89:9354–9358
Hussain SS, Ali M, Ahmad M, Siddique KHM (2011) Polyamines: natural and engineered abiotic and biotic stress tolerance in plants. Biotechnol Adv 29:300–311
Islam MM, Hoque MA, Okuma E, Banu MNA, Shimoishi Y, Nakamura Y, Murata Y (2009) Exogenous proline and glycine betaine increase antioxidant enzyme activities and confer tolerance to cadmium stress in cultured tobacco cells. J Plant Physiol 166:1587–1597
Jagendorf AT, Takabe T (2001) Inducers of glycine betaine synthesis in barley. Plant Physiol 127:1827–1835
Jagesh K, Tiwari AD, Munshi RK, Raghu N, Pandey A, Bhat AKS (2010) Effect of salt stress on cucumber: Na+ −K+ ratio, osmolyte concentration, phenols and chlorophyll content. Acta Physiol Plant 32:103–114
Jain M (2013) Emerging role of metabolic pathways in abiotic stress tolerance. J Plant Biochem Physiol 1:108
Jain M, Tiwary S, Gadre R (2010) Sorbitol-induced changes in various growth and biochemical parameters in maize. Plant Soil Environ 6:263–267
Kadioglu A, Saruhan N, Saglam A, Terzi R, Acet T (2011) Exogenous salicylic acid alleviates effects of long term drought stress and delays leaf rolling by inducing antioxidant system. Plant Growth Regul 64:27–37
Kalaji HM, Govindjee, Bosa K, Koscielniak J, Zuk-Gołaszewska K (2011) Effects of salt stress on photosystem II efficiency and CO2 assimilation of two Syrian barley landraces. Environ Exp Bot 73:64–72
Kanayama Y, Mori H, Imaseki H, Yamaki S (1992) Nucleotide sequence of a cDNA encoding sorbitol-6-phosphate dehydrogenase from apple. Plant Physiol 100:1607–1608
Kanayama Y, Watanabe M, Moriguchi R, Deguchi M, Kanahama K, Yamaki S (2006) Effects of low temperature and abscisic acid on the expression of the sorbitol-6-phosphate dehydrogenase gene in apple leaves. J Jpn Soc Hortic Sci 75:20–25
Kawakami A, Yoshida M (2005) Fructan: fructan 1-fructosyltransferase, a key enzyme for biosynthesis of graminan oligomers in hardened wheat. Planta 223:90–104
Kaya C, Sonmez O, Aydemir S, Ashraf M, Dikilitas M (2013) Exogenous application of mannitol and thiourea regulates plant growth and oxidative stress responses in salt-stressed maize (Zea mays L.). J Plant Interact 3:234–241
Kerepesi I, Galiba G (2000) Osmotic and salt stress-induced alteration in soluble carbohydrate content in wheat seedlings. Crop Sci 40:482–487
Khattab EA, Afifi MH (2009) Effect of proline and glycinebetain on canola plants grown under salinity stress condition. Mod J Appl Biol Sci Crop Sci 3:42–51
Kim TH, Bohmer M, Hu H, Nishimura N, Schroeder JI (2010) Guard cell signal transduction network: advances in understanding abscisic acid, CO2, and Ca2+ signaling. Annu Rev Plant Biol 61:561–591
Koyro HW, Ahmad P, Geissler N (2012) Abiotic stress responses in plants: an overview. In: Ahmad P, Prasad MNV (eds) Environmental adaptations and stress tolerance of plants in the era of climate change. Springer, New York, pp 1–28
Kubiś J (2008) Exogenous spermidine alters in different ways activities of some scavenging system enzymes, H2O2 and superoxide radical levels in water stressed cucumber leaves. J Plant Physiol 165:397–406
Kubiś J, Floryszak-Wieczorek J, Arasimowicz-Jelonek M (2014) Polyamines induce adaptive responses in water deficit stressed cucumber roots. J Plant Res 127:151–158
Kumar SG, Reddy AM, Sudhakar C (2003) NaCl effects on proline metabolism in two high yielding genotypes of mulberry (Morus alba L.) with contrasting salt tolerance. Plant Sci 165:1245–1251
Kusano T, Yamaguchi K, Berberich T, Takahashi Y (2007) Advances in polyamine research in 2007. J Plant Res 120:345–350
Kuznetsov V, Shevyakova NI (2010) Polyamines and plant adaptation to saline environments. In: Remawat KG (ed) Desert Plants. Springer, Berlin, pp 261–298
Lang F (2007) Mechanisms and significance of cell volume regulation. J Am Coll Nutr 26:613S–623S
Le TN, McQueen-Mason SJ (2006) Desiccation-tolerant plants in dry environments. Rev Environ Sci Biotechnol 5:269–279
Li F, Lei HJ, Zhao XJ, Tian RR, Li TH (2011) Characterization of three sorbitol transporter genes in micropropagated apple plants grown under drought stress. Plant Mol Biol Rep 30:123–130
Liu JH, Kitashiba H, Wang J, Ban Y, Moriguchi T (2007) Polyamines and their ability to provide environmental stress tolerance to plants. Plant Biotechnol 24:117–126
Liu C, Zhao L, Yu G (2011) The dominant glutamic acid metabolic flux to produce gamma-amino butyric acid over proline in Nicotiana tabacum leaves under water stress relates to its significant role in antioxidant activity. J Integr Plant Biol 53:608–618
Livingston DP, Hincha DK, Heyer AG (2009) Fructan and its relationship to abiotic stress tolerance in plants. Cell Mol Life Sci 66:2007–2023
Luo Y, Li F, Wang GP, Yang XH, Wang W (2010) Exogenous supplied trehalose protects thylakoid membranes of winter wheat from heat-induced damage. Biol Plant 54:495–501
Mehta P, Jajoo A, Mathur S, Bharti S (2010) Chlorophyll a fluorescence study revealing effects of high salt stress on photosystem II in wheat leaves. Plant Physiol Biochem 48:16–20
Minguet EG, Vera-Sirera F, Marina A, Carbonell J, Blázquez MA (2008) Evolutionary diversification in polyamine biosynthesis. Mol Biol Evol 25:2119–2128
Mishra AK, Singh VP (2010) A review of drought concepts. J Hydrol 391:202–216
Mitoi EN, Holobiuc I, Blindu R (2009) The effect of mannitol on antioxidative enzymes In vitro long term cultures of Dianthus tenuifolius and Dianthus spiculifolius. Rom J Biol Plant Biol 54:25–30
Munns R, Tester M (2008) Mechanisms of salinity tolerance. Annu Rev Plant Biol 59:651–681
Munns R, James AJ, Läuchli A (2006) Approaches to increasing the salt tolerance of wheat and other cereals. J Exp Bot 57:1025–1043
Murakeozy EP, Nagy Z, Duhaze C, Bouchereau A, Tuba Z (2003) Seasonal changes in the levels of compatible osmolytes in three halophytic species of inland saline vegetation in Hungary. J Plant Physiol 160:395–401
Nathawat NS, Kuhad MS, Goswami CL, Patel AL, Kumar R (2005) Nitrogen-metabolizing enzymes: effect of nitrogen sources and saline irrigation. J Plant Nut 28:1089–1101
Nayyar H, Chander S (2004) Protective effects of polyamines against oxidative stress induced by water and cold stress in chickpea. J Agron Crop Sci 190:355–365
Nayyar H, Walia DP (2003) Water stress induced proline accumulation in contrasting wheat genotypes as affected by calcium and abscisic acid. Biol Plant 46:275–279
Nidetzky B, Haltrich D, Schmidt K, Schmidt H, Weber A, Kulbe KD (1996) Simultaneous enzymatic synthesis of mannitol and gluconic acid: II. Development of a continuous process for a coupled NAD(H)-dependent enzyme system. Biocatal Biotrans 14:47–65
Nishiyama R, Watanabe Y, Fujita Y, Le DT, Kojima M, Werner T, Vankova R, Yamaguchi-Shinozaki K, Shinozaki K, Kakimoto T, Sakakibara H, Schmülling T, Phan Trana LS (2011) Analysis of cytokinin mutants and regulation of cytokinin metabolic genes reveals important regulatory roles of cytokinins in drought, salt and abscisic acid responses, and abscisic acid biosynthesis. Plant Cell 23:2169–2183
Niu X, Bressan RA, Hasegawa PM, Pardo JM (1995) Ion homeostasis in NaCl stress environments. Plant Physiol 109:735–742
Nounjana N, Nghiab PT, Theerakulpisuta P (2012) Exogenous proline and trehalose promote recovery of rice seedlings from salt-stress and differentially modulate antioxidant enzymes and expression of related genes. J Plant Physiol 169:596–604
Nyyssola A, Kerovuo J, Kaukinen P, Von Weymarn N, Reinikainen T (2000) Extreme halophiles synthesize betaine from glycine by methylation. J Biol Chem 275:22196–22201
Osakabe Y, Yamaguchi-Shinozaki K, Shinozaki K, Phan Tran LS (2014) ABA control of plant macroelement membrane transport systems in response to water deficit and high salinity. New Phytol 202:35–49
Oukarroum A, El Madidi S, Strasser RJ (2012) Exogenous glycine betaine and proline play a protective role in heat-stressed barley leaves (Hordeum vulgare L.): a chlorophyll a fluorescence study. Plant Biosyst 4:1037–1043
Parida AK, Dagaonkar VS, Phalak MS, Aurangabadkar LP (2008) Differential responses of the enzymes involved in proline biosynthesis and degradation in drought tolerant and sensitive cotton genotypes during drought stress and recovery. Acta Physiol Plant 5:619–627
Parihar P, Singh S, Singh R, Singh VP, Prasad SM (2014) Effect of salinity stress on plants and its tolerance strategies: a review. Environ Sci Pollut Res. doi:10.1007/s11356-014-3739-1
Patonnier MP, Peltier JP, Marigo G (1999) Drought-induced increase in xylem malate and mannitol concentrations and closure of Fraxinus excelsior L. stomata. J Exp Bot 50:1223–1229
Paul M, Pellny T, Goddijn O (2001) Enhancing photosynthesis with sugar signals. Trends Plant Sci 6:197–200
Paul MJ, Primavesi LF, Jhurreea D, Zhang YH (2008) Trehalose metabolism and signaling. Annu Rev Plant Biol 59:417–441
Peshev D, Vergauwen R, Moglia A, Hideg É, Ende WVD (2013) Towards understanding vacuolar antioxidant mechanisms: a role for fructans? J Exp Bot 64:1025–1038
Pilon-Smits E, Ebskamp M, Paul MJ, Jeuken M, Weisbeek PJ, Smeekens S (1995) Improved performance of transgenic fructan-accumulating tobacco under drought stress. Plant Physiol 107:125–130
Pilon-Smits EAH, Terry N, Sears T, Van Dun K (1999) Enhanced drought resistance in fructan-producing sugar beet. Plant Physiol Biochem 37:313–317
Ranganayakulu GS, Veeranagamallaiah G, Sudhakar C (2013) Effect of salt stress on osmolyte accumulation in two groundnut cultivars (Arachis hypogaea L.) with contrasting salt tolerance. Afr J Plant Sci 12:586–592
Reguera M, Peleg Z, Blumwald E (2012) Targeting metabolic pathways for genetic engineering abiotic stress-tolerance in crops. Biochim Biophys Acta 1819:186–194
Rhodes D, Hanson AD (1993) Quaternary ammonium and tertiary sulfonium compounds in higher plants. Annu Rev Plant Physiol Mol Biol 44:357–384
Rivero RM, Mestre TC, Mittler R, Rubio F, Garcia-Sanchez F, Martinez V (2014) The combined effect of salinity and heat reveals a specific physiological, biochemical and molecular response in tomato plants. Plant Cell Environ 37:1059–1073
Rubio JS, Rubio F, Martínez V, García-Sánchez F (2010) Amelioration of salt stress by irrigation management in pepper plants grown in coconut coir dust. Agric Water Manag 97:1695–1702
Sakamoto A, Murata N (2002) The role of glycine betaine in the protection of plants from stress: clues from transgenic plants. Plant Cell Environ 25:163–171
Sawahel W (2004) Improved performance of transgenic glycine betaine-accumulating rice plants under drought stress. Biol Plant 47:39–44
Shahbaz M, Mushtaq Z, Andaz F, Masood A (2013) Does proline application ameliorate adverse effects of salt stress on growth, ions and photosynthetic ability of eggplant (Solanum melongena L.)? Sci Hortic 164:507–511
Sharma SS, Dietz KJ (2006) The significance of amino acids and amino acid-derived molecules in plant responses and adaptation to heavy metal stress. J Exp Bot 57:711–726
Sheveleva E, Chmara W, Bohnert HJ, Jensen RG (1997) Increased salt and drought tolerance by d-ononitol production in transgenic Nicotiana tabacum L. Plant Physiol 115:1211–1219
Shinozaki K, Yamaguchi-Shinozaki K (2007) Gene networks involved in drought stress response and tolerance. J Exp Bot 58:221–227
Siddiqui MH, Khan MN, Mohammad F, Khan MMA (2008) Osmoprotectant accumulation in Brassica juncea L. under salt stress. J Agron Crop Sci 194:214–224
Szabados L, Savouré A (2009) Proline: a multifunctional amino acid. Trends Plant Sci 15:89–97
Szabolcs I (1989) Salt affected soils. CRC Press, Boca Raton
Taiz L, Zeiger E (2003) Plant Physiology, 3rd Edn. Panima Publishing Co., New delhi, pp 591–620
Talat A, Nawaz K, Hussian K, Bhatti KH, Siddiqi EH, Khalid A, Anwer S, Sharif MU (2013) Foliar application of proline for salt tolerance of two wheat (Triticum aestivum L.) cultivars. World Appl Sci J 4:547–554
Tari I, Kiss G, Deér AK, Csiszár J, Erdei L, Gallé Á, Gémes K, Horváth F, Poór P, Szepesi Á, Simon LM (2010) Salicylic acid increased aldose reductase activity and sorbitol accumulation in tomato plants under salt stress. Biol Plant 54:677–683
Theerakulpisut P, Gunnula W (2012) Exogenous sorbitol and trehalose mitigated salt stress damage in salt-sensitive but not salt tolerance rice seedlings. Asian J Crop Sci 4:165–170
Türkan I, Demiral T (2009) Recent developments in understanding salinity tolerance. Environ Exp Bot 67:2–9
Van den Ende W, Valluru R (2009) Sucrose, sucrosyl oligosaccharides, and oxidative stress: scavenging and salvaging? J Exp Bot 60:9–18
Vandesteenea L, Ramonb M, Royc KL, Dijckd PV, Rollanda F (2010) A single active trehalose-6-P synthase (TPS) and a family of putative regulatory TPS-like proteins in Arabidopsis. Mol Plant 2:406–419
Vardharajula S, Ali SZ, Grover M, Reddy G, Bandi V (2011) Drought-tolerant plant growth promoting Bacillus spp.: effect on growth, osmolytes, and antioxidant status of maize under drought stress. J Plant Interact 1:1–14
Verbruggen N, Hermans C (2008) Proline accumulation in plants: a review. Amino Acids 35:753–759
Verma S, Mishra SN (2005) Putrescine alleviation of growth in salt stressed Brassica juncea by inducing antioxidative defense System. J Plant Physiol 162:669–677
Villa-Castorena M, Ulery AL, Catalan-Valencia EA, Remmenga MD (2003) Salinity and nitrogen rate effects on the growth and yield of chile pepper plants. Soil Sci Soc Am J 67:1781–1789
Vinocur B, Altman A (2005) Cellular basis of salinity tolerance in plants. Environ Exp Bot 52:113–122
Wahomea PK, Jeschb HH, Grittnerb I (2001) Mechanisms of salt stress tolerance in two rose root stocks: Rosa chinensis ‘Major’ and R. rubiginosa. Sci Hortic 87:207–216
Wang HY, Huang YC, Chen SF, Yeh KW (2003) Molecular cloning, characterization and gene expression of a water deficiency and chilling induced proteinase inhibitor I gene family from sweet potato (Ipomoea batatas Lam.) leaves. Plant Sci 165:191–203
Wani SH, Gosal SS (2011) Introduction of OsglyII gene into Indicam rice through particle bombardment for increased salinity tolerance. Biol Plant 55:536–540
Wani SH, Lone AA, Da Silva T, Gosal SS (2010) Effects of NaCl stress on callus induction and plant regeneration from mature seeds of rice (Oryza sativa L.). Asian Aust J Plant Sci Biotechnol 4:56–71
Wani SH, Singh NB, Haribhushan A, Mir JI (2013) Compatible solute engineering in plants for abiotic stress tolerance-role of glycine betaine. Curr Genomics 14:157–165
Wei GP, Yang LF, Zhu YL, Chen G (2009) Changes in oxidative damage, antioxidant enzyme activities and polyamine contents in leaves of grafted and non-grafted eggplant seedlings under stress by excess of calcium nitrate. Sci Hortic 120:443–451
Williamson JD, Jennings DB, Guo WW, Pharr DM, Ehrenshaft M (2002) Sugar alcohols, salt stress and fungal resistance: polyols: multifunctional plant protection? J Am Soc Hortic Sci 127:467–473
Wu BH, Li SH, Nosarzewski M, Archbold DD (2010) Sorbitol dehydrogenase gene expression and enzyme activity in apple: tissue specificity during bud development and response to rootstock vigor and growth manipulation. J Am Soc Hortic Sci 135:379–387
Wu X, Zhu Z, Li X, Zha D (2012) Effects of cytokinin on photosynthetic gas exchange, chlorophyll fluorescence parameters and antioxidative system in seedlings of eggplant (Solanum melongena L.) under salinity stress. Acta Physiol Plant 34:2105–2114
Yadav G, Srivastava PK, Singh VP, Prasad SM (2014) Light intensity alters the extent of arsenic toxicity in Helianthus annuus L. seedlings. Biol Trace Elem Res 158:410–421
Yamaguchi K, Takahashi Y, Berberich T, Imai A, Takahashi T, Michael AJ, Kusano TA (2007) Protective role for the polyamine spermine against drought stress in Arabidopsis. Biochem Biophys Res Commun 352:486–490
Yu GH, Li W, Yuan ZY, Cui HY, Lv CG, Gao ZP, Han B, Gong YZ, Chen GX (2013) The effects of enhanced UV-B radiation on photosynthetic and biochemical activities in super-high-yield hybrid rice Liangyoupeijiu at the reproductive stage. Photosynthetica 51:33–44
Zayed AM, Terry N (2003) Chromium in the environment: factors affecting biological remediation. Plant Soil 249:139–156
Zhang LX, Lai JH, Liang ZS, Ashraf M (2014) Interactive effects of sudden and gradual drought stress and foliar-applied glycine betaine on growth, water relations, osmolyte accumulation and antioxidant defence system in two maize cultivars differing in drought tolerance. J Agron Crop Sci 200:425–433
Zhou R, Cheng LL, Wayne R (2003) Purification and characterization of sorbitol-6- phosphate phosphatase from apple leaves. Plant Sci 165:227–232
Zhu JK (2001) Plant salt tolerance. Trends Plant Sci 6:66–71
Acknowledgments
Authors are thankful to the University Grants Commission, New Delhi, for financial assistance to carry out this work. Dr. Vijay Pratap Singh is thankful to Central Regional Office, Bhopal of University Grants Commission, New Delhi for providing financial assistance (PI-UGC Minor Research Project, File No. MS-27/201022/XII/13-14/CRO). Jitendra Kumar, is thankful to UGC, New Delhi for providing the financial support as SRF under Rajiv Gandhi National Fellowship (Ref. No. RGNF-2012-13-SC-UTT-33185).
Author information
Authors and Affiliations
Corresponding authors
Rights and permissions
About this article

Cite this article
Singh, M., Kumar, J., Singh, S. et al. Roles of osmoprotectants in improving salinity and drought tolerance in plants: a review. Rev Environ Sci Biotechnol 14, 407–426 (2015). https://doi.org/10.1007/s11157-015-9372-8
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11157-015-9372-8