
Abstract
Maple syrup urine disease (MSUD) or branched-chain α-keto aciduria (BCKA) is an inherited disorder caused by a deficiency of the branched-chain α-keto acid dehydrogenase complex (BCKAD) activity. The blockage of this pathway leads to tissue accumulation of the branched-chain amino acids (BCAA) leucine, isoleucine and valine and their respective keto-acids. The clinical features presented by MSUD patients include ketoacidosis, convulsions, coma, psychomotor delay and mental retardation. The mechanism of brain damage in this disease is still poorly understood. However, an increase in lipid peroxidation in vitro in cerebral cortex of young rats as well as a decrease in the antioxidant defenses has been previously observed. In the present work we evaluated different oxidative stress parameters, named reactive species of thiobarbituric acid (TBARS), total antioxidant reactivity (TAR) and total antioxidant status (TAS) in plasma of MSUD patients in order to evaluate whether oxidative stress is involved in this disorder. We verified a marked increase of plasma TBARS measurements, which is indicative of increased lipid peroxidation, as well as a decrease on plasma TAR reflecting a deficient capacity to efficiently modulate the damage associated with an increased production of reactive species. In contrast, TAS was not changed indicating that the total content of antioxidants in plasma of patients affected by MSUD was not altered. These results suggest that free radical generation is elicited in MSUD and is possibly involved in the pathophysiology of the tissue damage found in this disorder.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Maple syrup urine disease (MSUD) or branched-chain α-keto aciduria (BCKA) is an autosomal recessive metabolic disorder caused by a severe deficiency of the branched-chain α-keto acid dehydrogenase complex (BCKAD) activity. The blockage in this enzyme complex leads to tissue accumulation of the branched-chain amino acids (BCAA) leucine, isoleucine and valine as well as their corresponding branched chain α-keto acids (BCKA) α-ketoisocaproate, α-keto-β-methylvalerate and α-ketoisovalerate, respectively (Chuang and Shih, 2001; Treacy et al., 1992).
The major clinical features presented by MSUD patients include ketoacidosis, hypoglycemia, poor feeding, apnea, ataxia, convulsions, coma, psychomotor delay and mental retardation. Magnetic resonance imaging studies have demonstrated generalized edema and hypomyelination/demyelination in central nervous system (CNS) of MSUD patients (Chuang and Shih, 2001; Schönberger et al., 2004).
MSUD presents heterogeneous molecular and clinical phenotypes range from a severe classic form with neonatal onset to milder variant forms with later onset, and presenting different residual enzyme activity (Chuang and Shih, 2001; Schadewaldt and Wendel, 1997).
The aim of MSUD treatment is to keep the BCAA plasma concentrations in the normal range, protecting the brain from injury. The treatment consists of a low protein diet and a semi-synthetic formula restricted in BCAA and supplemented with essential amino acids. Metabolic intoxication may cause a fatal outcome in untreated patients (Chuang and Shih, 2001; Danner and Elsas, 1989).
The mechanisms of the neurological symptoms presented by MSUD patients are still poorly understood. However, considering that increased concentrations of leucine and/or α-ketoisocaproate were associated with the appearance of neurological symptoms, these compounds seem to be the main important neurotoxic metabolites in MSUD (Chuang and Shih, 2001; Snyderman et al., 1964). Furthermore, it has been demonstrated that the metabolities accumulating in MSUD cause impairment of energy metabolism by inhibiting the electron transport chain (Sgaravatti et al., 2003) and creatine kinase activity in rat brain (Pilla et al., 2003). Other investigators demonstrated that the BCAA and/or BCKA that accumulate in MSUD provoke neuronal apoptosis (Jouvet et al., 2000), as well as convulsions (Coitinho et al., 2001), impairment of neurotransmitter synthesis and function (Zielke et al., 1996; Tavares et al., 2000), myelin alteration (Treacy et al., 1992; Tribble and Shapira, 1983; Taketomi et al., 1983) and reduced uptake of essential amino acids by the brain (Araújo et al., 2001).
Free radicals seem to be involved in a large number of human diseases. Increasing evidence has shown that damage caused by free radicals is an important contributing factor in chronic-inflammatory, vasculary, neoplastic and neurodegenerative diseases (Halliwell, 1994; Reznick and Packer, 1993; Przedborski et al., 1996; Bem-Menachem et al., 2000).
Oxidative stress has been observed in some inborn errors of intermediary metabolism owing to the accumulation of toxic metabolites which leads to excessive free radical production (Colome et al., 2000). Restricted diets utilized to treat patients affected by metabolic disorders may result in a low antioxidant status (Colome et al., 2000).
Recently, it was demonstrated that the BCAA and their respective BCKA that accumulate in MSUD stimulate in vitro lipid peroxidation in brain homogenates of rats (Fontella et al., 2002). It was later demonstrated that these compounds, particularly leucine and α-ketoisocaproate, not only stimulate in vitro lipid peroxidation but also reduce the cerebral capacity to modulate the damage associated with the increased free radical production (Bridi et al., 2003, 2005a). Furthermore, it was shown that the increased lipid peroxidation induced by leucine could be attenuated by the free radicals scavengers ascorbic acid, α-tocopherol, gluthatione and superoxide dismutase (Bridi et al., 2005b).
The aim of the present work was to evaluate some parameters of oxidative stress, namely thiobarbituric acid-reactive substances (TBARS), total antioxidant reactivity (TAR) and total antioxidant status (TAS), in plasma of MSUD patients at the time of diagnosis in order to verify whether free radicals could be involved in the pathophysiology of this disease.
Material and methods
Patients and controls
Plasma from five MSUD patients (classic form) aged between 15 days and 4 months at diagnosis were used to evaluate the parameters of oxidative stress. The most common clinical features presented by these patients were convulsions, hypoglycemia, poor feeding, ketoacidosis and psychomotor delay. Samples were obtained at the time of the diagnosis, which was made by increased plasma levels of leucine (2,346.1±810.7 μmol/L), isoleucine (304.8±185.2 μmol/L) and valine (456.5±275.1 μmol/L) by HPLC method (Joseph and Marsden, 1986). Control group was composed of healthy age matched individuals (leucine 158.33±37.63 μmol/L; isoleucine 76.54±18.02 μmol/L; valine 260.73±39.79 μmol/L).
Reagents
All chemicals were of PA purity and were purchased from Sigma (St. Louis, MO, USA) except for thiobarbituric acid, which was purchased from Merck (Darmstadt, Germany) and a kit for TAS measurement that was purchased from Randox Laboratories (Antrim, United Kingdom). TAR was assayed using a beta liquid scintillation spectrometer (Wallac model 1409) and TBARS was measured with a spectrofluorimeter (Hitachi F2000).
Plasma preparation
Plasma was prepared from whole blood samples obtained from fasting individuals (controls and MSUD patients) by venous puncture with heparinized vials. Whole blood was centrifuged at 1,000g. Plasma was removed by aspiration and frozen at −80°C until determination.
Thiobarbituric Acid-Reactive Species (TBARS)
Thiobarbituric acid-reactive substances (TBARS) were determined according to the method described by Buege and Aust (1978). Briefly, 250 μL of 10% trichloroacetic acid were added to 125 μL of plasma, then 375 μl 0.67% thiobarbituric acid (in 7.1% sodium sulphate) were added and incubated at 100°C for 30 min. After the incubation, the mixture was extracted with 750 μL butanol. The resulting pink stained TBARS were determined in a spectrofluorimeter at 515 nm. Calibration curve was performed using 1,1,3,3-tetramethoxypropane subjected to the same treatment as that of the samples. TBARS were calculated as nmol TBARS/mg protein.
Total Antioxidant Reactivity (TAR)
TAR, which represents the quality of the tissue antioxidants, was determined by measuring the luminol chemiluminescence intensity induced by 2,2′-azo-bis-(2-amidinopropane) (ABAP) according to the method of Lissi et al. (1992). The background chemiluminescence was measured by adding 4 mL of 2 mM ABAP (in 0.1 M glycine buffer, pH 8.6) into a glass scintillation vial. Ten microliters of luminol (4 mM) were added to each vial and the chemiluminescence was measured. This was considered to be the basal value. Ten microliters of 25–200 μM Trolox (curve calibration) or plasma was then added and the chemiluminescence was measured during 60 s. The Trolox and plasma addition reduces the chemiluminescence. The rapid reduction in luminol intensity is considered as a measure of the TAR capacity. TAR measurement was calculated as nmol Trolox/mg protein.
Total Antioxidant Status (TAS)
TAS, which represents the quantity of the tissue antioxidants, was determined by using a kit from RANDOX Laboratories. The plasma sample was incubated with ABTS (2,2′-azino-di-[3-ethylbenzthiazoline sulphonate]) plus a peroxidase (metmyoglobin) and H2O2 to produce the cation ABTS+. A relatively stable blue–green color occurred and was measured at 37°C at 600 nm. Antioxidants in the added sample cause suppression of this color production to a degree which was proportional to their concentration (Miller et al., 1993; Yu and Ong, 1999). The results were expressed in mmol/L plasma.
Protein determination
Protein concentrations were determined by the biuret method from Labtest® (Gornall et al., 1949), using albumin as standard.
Statistical analysis
The Student t test was used to compare results from controls and MSUD patients. A p value less than 0.05 was considered significant. All analyses were performed using the Statistical Package for the Social Sciences (SPSS) software in a PC-compatible computer.
Results
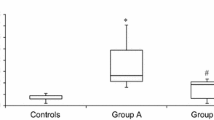
Figure 1 shows that TBARS was significantly increased in plasma of MSUD patients when compared to control group [t(8)=−4.552, p < 0.01], indicating that lipid peroxidation is stimulated in MSUD patients.

Plasma thiobarbituric acid reactive species (TBARS) from MSUD patients and controls. Data represent the mean ± SD (n=5). ** p < 0.01 (Student’ t test for unpaired samples) compared to control
TAR measurement, which is a measure of the tissue capacity to react with free radicals, was markedly reduced [t(8)=3.021, p < 0.05] in plasma of MSUD patients (Fig. 2). These results suggest a deficient capacity of plasma to modulate the damage associated with the enhanced production of reactive species in these MSUD patients. Finally, it was also observed that TAS measurement, which represents the quantity of the tissue antioxidants, was not altered in plasma of MSUD patients [t(6)=−0.713, p > 0.05] (Fig. 3), suggesting that the total nonenzymatic antioxidant defenses were not altered.

Plasma total antioxidant reactivity (TAR) from MSUD patients and controls. Data represent the mean ± SD (n=5). * p < 0.05 (Student’ t test for unpaired samples) compared to control

Plasma total antioxidant status (TAS) from MSUD patients and controls. Data represent the mean ± SD (n=4). No significant differences between means were found (Student’ t test for unpaired samples)
Discussion
Neurological symptoms are frequent in MSUD patients and untreated patients normally have a fatal outcome (Chuang and Shih, 2001; Danner and Elsas, 1989). Leucine and/or α-ketoisocaproate are considered the main neurotoxic metabolites in these disease (Chuang and Shih, 2001; Snyderman et al., 1964). However, the mechanisms underlying the sequelae presented by these patients are not well understood.
It was previously demonstrated that the BCAA and BCKA accumulating in MSUD stimulate in vitro lipid peroxidation and reduce the antioxidant defenses in cerebral homogenates of young rats (Fontella et al., 2002; Bridi et al., 2003, 2005a). However, to our knowledge there is no report in the literature assessing whether oxidative stress occurs in tissues from MSUD patients. Therefore, in the present study we investigated some parameters of oxidative stress in plasma from these patients which were not under any therapy. So our results cannot be attributed to any medication.
We demonstrated a significant increase of TBARS in plasma from MSUD patients. Considering that TBARS reflects the content of malondialdehyde, an end product of lipid breakdown due to lipid peroxidation (Halliwell and Gutteridge, 2001; Esterbauer and Cheeseman, 1990), our data indicate that lipid peroxidation is induced in MSUD patients, probably secondary to free radical generation. Despite the fact that we did not find any decrease of total antioxidant defenses in plasma from these patients as indicated by TAS values TAR, which corresponds to a useful index of the capacity of a given tissue to modulate the damage associated with an increased production of free radicals and mainly reflects the quality of antioxidants (Lissi et al., 1995), was significantly decreased in the patients studied. These results indicate a deficient capacity of plasma from MSUD patients to rapidly handle an increase of reactive species. Considering that an imbalance between the total antioxidant defenses and the reactive species formed in the tissues are indicative of oxidative stress (Halliwell and Gutteridge, 2001), it is proposed that free radical generation is involved in the pathophysiology of the tissue damage found in MSUD.
Oxidative stress has been considered an important contributor to brain damage in neurodegenerative diseases, seizures and demyelination (Halliwell, 2001; Méndez-Álvarez et al., 2001; Karelson et al., 2001), probably because brain has relatively low levels of antioxidant defenses (Halliwell and Gutteridge, 2001), as well as high lipid content, specially unsaturated fatty acids, and iron that stimulates the Fenton reaction being therefore highly susceptible to reactive species attack. Taken together our present in vivo results and those demonstrating in studies in vitro with rats that the metabolities accumulated in MSUD cause a stimulation of lipid peroxidation and a reduction of brain antioxidant defenses, (Fontella et al., 2002; Bridi et al., 2003, 2005a), suggest that oxidative stress is probably involved in the pathophysiology of MSUD.
To our knowledge this is the first report demonstrating increased oxidative stress in patients affected by MSUD. Our results should, however, be taken with caution and confirmed with a higher number of patients and with other techniques to measure oxidative stress. In this context, CSF specimens may be useful in order to evaluate if the brain is also a target for reactive species. In case the present results are confirmed, we may conclude that oxidative stress contributes at least in part to the severe neurological dysfunction found in MSUD.
References
Araújo P, Wassermann GF, Tallini K, Furlanetto V, Vargas CR, Wannmacher CMD, Dutra-Filho CS, Wyse ATS, Wajner M (2001) Reduction of large neutral amino acid level in plasma and brain of hyperleucinemic rats. Neurochem Int 38:529–537
Bem-Menachem E, Kyllerman R, Markleind S (2000) Superoxide dismutase and glutathione peroxidase function in progressive myoclonus epilepsies. Epilepsy Res 40:33–39
Bridi R, Araldi J, Sgarbi MB, Testa CG, Durigon K, Wajner M, Dutra-Filho CS (2003) Induction of oxidative stress in rat brain by the metabolites accumulating in maple syrup urine disease. Int J Devl Neuroscience 21:327–332
Bridi R, Braun CA, Zorzi GK, Wannmacher CM, Wajner M, Lissi EG, Dutra-Filho CS (2005a) Alpha-keto acids accumulating in maple syrup urine disease stimulate lipid peroxidation and reduce antioxidant defences in cerebral cortex from young rats. Metab Brain Dis Jun 20(2):155–167
Bridi R, Latini A, Braum CA, Zorzi GK, Wajner M, Lissi E, Dutra-Filho CS (2005b) Evaluation of the mechanism involved in leucine-induced oxidative damage in cerebral córtex of young rats. Free Radic Res Jan 39(1):71–79
Buege JA, Aust SD (1978) Microssomal lipid peroxidation. Methods Enzymol 52:302–309
Chuang DT, Shih VE (2001) Maple syrup urine disease (branche-chain ketoaciduria). In: Scriver CR, Beaudt AL, Sly WL, Valle D (eds) The metabolic and molecular bases of inherited disease. McGraw-Hill, New York, pp 1971–2005
Coitinho AS, de Mello CF, Lima TT, de Bastiani J, Fighera MR, Wajner M (2001) Pharmacological evidence that alpha-keto isovaleric acid induces convulsions through GABAergic and glutamatergic mechanisms in rats. Brain Res 894:68–73
Colome C, SIerra C, Vilaseca MA (2000) Congenital errors of metabolism: Cause of oxidative stress? Med Clin 115(3):111–117
Danner DJ, Elsas JL II (1989) Disorders of branched chain amino acid and keto acid metabolism. In: Scriver CR, Beaudt AL, Sly WL, Valle D (eds) The metabolic and molecular bases of inherited disease. McGraw-Hill, New York, pp 671–692
Esterbauer H, Cheeseman KH (1990) Determination of aldehydic lipid peroxidation products: Malonaldehyde and 4-hydroxynonenal. Methods Enzymol 186:407–421
Fontella FU, Gassen E, Pulrolnik V, Wannmacher CMD, Klein AB, Wajner M, Dutra CS (2002) Stimulation of lipid peroxidation in vitro in rat brain by metabolites accumulating in maple syrup urine disease. Metab Brain Dis 17:47–54
Gornall AG, Bardawill CJ, David MM (1949) Determination of serum proteins by means of the biuret reaction. J Biol Chem 177:751–766
Halliwell B (1994) Free radicals, antioxidants and human disease: Curiosity, cause or consequence? Lancet 344:721–724
Halliwell B (2001) Role of free radicals in the neurodegenerative diseases. Drugs Aging 18:685–716
Halliwell B, Gutteridge JMC (2001) Detection of free radicals and others reactive species: Trapping and fingerprinting. In: Halliwell B, Gutteridge JMC (eds) Free radicals in biology and medicine. Oxford University Press, Oxford, UK, pp 351–425
Joseph MH, Marsden CA (1986) Amino acids and small peptides. In: Lim CF (ed) HPLC of small peptides. IRL Press, Oxford, pp 13–27
Jouvet P, Rustin P, Taylor DL, Pocock JM, Felderhoff-Mueser U, Mazarakis ND, Sarraf C, Joashi U, Koszma M, Greewood K, Edwards AD, Mehmet H (2000) Branched chain amino acids induce apoptosis in neural cells without mitochondrial membrane despolarization or cytochrome c release: Implications for neurological impairment associated with maple syrup urine disease. Mol Biol Cell 11:1919–1932
Karelson E, Bogdanovic N, Garlind A, Winblad B, Zilmer K, Kullisaar T, Vihalemm T, Kairane C, Zilmer M (2001) The cerebrocortical areas in normal brain aging and in the Alzheimer’s disease: Noticeable difference in the lipid peroxidation level and in antioxidant defense. Neurochem Res 26:353–361
Lissi E, Pascual C, Del Castillo MD (1992) Luminol luminescence induced by 2,2′-azo-bis-(2-amidinopropane) thermolysis. Free Rad Res Commun 17:299–311
Lissi E, Salim-Hanna M, Pascual C, Del Castillo MD (1995) Evaluation of total antioxidant potential (TRAP) and total antioxidant reactivity from luminol-enhanced chemiluminescence measurements. Free Radic Biol Med 18:153–158
Méndez-Álvarez E, Soto-Otero R, Hermida-Aeijeiras A, López-Real AM, Labandeira-García JL (2001) Effects of aluminium and zinc on the oxidative stress caused by 6-hydroxydopamine autoxidation: Relevance for the pathogenesis of Parkinson’s disease. Biochim Biophys Acta 1586:155–168
Miller NJ, Rice-Evans C, Davies MJ, Gopinathan V, Milner A (1993) A novel method for measuring antioxidant capacity and its application to monitoring the antioxidant status in premature neonates. Clin Sci 84:407–412
Pilla C, Cardozo RFD, Dutra CS, Wyze ATS, Wajner M, Wannmacher CMD (2003) Effect of leucine administration on creatine kinase activity in rat brain. Metab Brain Dis 18:17–25
Przedborski S, Donaldson DBS, Jakowec M, Kish JS, Guttman M, Rosoklija G, Hays AP (1996) Brain superoxide dismutase, catalase and glutathione peroxidase activities in amyotrophic lateral sclerosis. Ann Neurol 39:158–165
Reznick AZ, Packer L (1993) Free radicals and antioxidants in muscular neurological diseases and disorders. In: Poli G, Albano E, Dianzani MU (eds) Free radicals: From basic science to medicine. Birkhäuser Verlag, Basel, pp 425–437
Schadewaldt P, Wendel U (1997) Metabolism of branched-chain amino acids in maple syrup urine disease. Eur J Pediatr 156(Suppl 1):S62–S66
Schönberger S, Schweiger B, Schwahn B, Schwarz M, Wendel U (2004) Dysmyelination in the brain of adolescents and young adults with maple syrup urine disease. Mol Genet Metab 82:69–75
Sgaravatti AM, Rosa RB, Schuck PF, Ribeiro CAJ, Wannacher CMD,Wyse ATS, Dutra-Filho CS, Wajner M (2003) Inhibition of brain energy metabolism by the α-keto acids accumulating in maple syrup urine disease. Biochim Biophys Acta 1639:232–238
Snyderman SE, Norton PM, Roitman E (1964) Maple syrup urine disease with particular reference to diet therapy. Pediatrics 34:454–472
Taketomi T, Kunishita T, Hara A, Mizushima S (1983) Abnormal protein and lipid compositions of the cerebral myelin in patient with maple syrup urine disease. Jpn J Exp Med 53:109–116
Tavares RG, Santos CES, Tasca C, Wajner M, Souza DO, Dutra-Filho CS (2000) Inhibition of glutamate uptake into synaptic vesicles of rat brain by the metabolites accumulating in maple syrup urine disease. J Neurol Sci 181:44–49
Treacy E, Clow CL, Reade TR, Chitayat D, Mamer OA, Scriver CR (1992) Maple syrup urine disease: Interrelationship between branched chain amino-, oxo- and hydroxyacids; implications for treatment; association with CNS dysmelination. J Inherit Metab Dis 15:121–135
Tribble D, Shapira R (1983) Myelin proteins: Degradation in rat brain initiated by metabolites causative of maple syrup urine disease. Biochem Biophys Res Commun 114:440–446
Yu T-W, Ong ChN (1999) Lag-time measurement of antioxidant capacity using myoglobin and 2,29-azino-bis(3-ethyl-benzthiazoline-6-sulfonic acid): Rationale, application and limitation. Anal Biochem 275:217–223
Zielke HR, Huang Y, Tildon JT, Zielke CL, Baab PJ (1996) Elevation of amino acids in the interstitial space of the rat brain following infusion of large neutral amino and keto acids by microdialysis: Leucine infusion. Dev Neurosci 18:420–425
Acknowledgements
This work was supported by grants from Brazilian National Research Council (CNPq), FAPERGS, and FIPE/HCPA—Brazil.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Barschak, A.G., Sitta, A., Deon, M. et al. Evidence that oxidative stress is increased in plasma from patients with maple syrup urine disease. Metab Brain Dis 21, 279–286 (2006). https://doi.org/10.1007/s11011-006-9030-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11011-006-9030-5