
Abstract
We built empirical models to estimate the effects of land cover on stream ecosystems in the mid-Atlantic region (USA) and to evaluate the spatial scales over which such models are most effective. Predictive variables included land cover in the watershed, in the streamside corridor, and near the study site, and the number and location of dams and point sources in the watershed. Response variables were annual nitrate flux; species richness of fish, benthic macroinvertebrates, and aquatic plants; and cover of aquatic plants and riparian vegetation. All data were taken from publicly available databases, mostly over the Internet. Land cover was significantly correlated with all ecological response variables. Modeled R 2 ranged from 0.07 to 0.5, but large data sets often allowed us to estimate with acceptable precision the regression coefficients that express the change in ecological conditions associated with a unit change in land cover. Dam- and point-source variables were ineffective at predicting ecological conditions in streams and rivers, probably because of inadequacies in the data sets. The spatial perspective (whole watershed, streamside corridor, or local) most effective at predicting ecological response variables varied across response variables, apparently in concord with the mechanisms that control each of these variables. We found some evidence that predictive power fell in very small watersheds (less than 1–10 km2), suggesting that the spatial arrangement of landscape patches may become critical at these small scales. Empirical models can replace, constrain, or be combined with more mechanistic models to understand the effects of land-cover change on stream ecosystems.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Streams and rivers are among the ecosystems most affected by human activities (Dynesius and Nilsson 1994; Master and others 1997; Naiman and Turner 2000). The species composition (Richter and others 1997; Jansson and others 2000a), food web structure (Wootton and others 1996), nutrient cycling (Johnes 1996; Meyer and others 1999; Vörösmarty and others 2000a), and utility (Postel 2000; Vörösmarty and others 2000b) of thousands of kilometers of streams and rivers worldwide have been greatly altered from their natural states. One of the main causes of these alterations is land-use change. Land use is changing rapidly around the world (McCloskey and Spalding 1989; Vitousek and others 1997; Turner and others 1998); by 1994 about 75% of the habitable part of the planet was disturbed by human activity (Hannah and others 1994). Land-use change affects stream ecosystems by altering the timing, amount, and kind of inputs of water, light, organic matter, and other materials to the channel, which can have profound consequences for all aspects of the stream ecosystem.
The precise relationships between land-use conversion and ecological responses are difficult to establish because: (1) the types of land use, rates of conversion, and spatial distribution of land use vary considerably among watersheds and regions and across political boundaries, (2) changes in land use can drive channel morphology and hydrology into a state of flux that may take many decades to stabilize (Fitzpatrick and Knox 2000), (3) ecological responses may lag behind physical habitat modifications (for example, see Harding and others 1998) and we do not always know the duration of such lag effects, and (4) management actions have been introduced to mediate the effects of development on streams, yet we know little about their effectiveness. Thus, understanding and predicting the effects of land-use change on stream and river ecosystems are difficult scientific problems and major challenges for contemporary ecology.
All available approaches to studying this problem have significant weaknesses. The ideal approach—replicated long-term experiments of entire watersheds—is usually prohibitively costly, logistically impractical, and requires decades to deliver an answer. Experimentation at smaller scales of space and time can still be costly and logistically difficult. The results of such studies may be difficult to generalize to other sites. Further, unless the experimental study is long-term, its findings may be dominated by transient responses of the system that do not resemble its long-term responses (see, for example, Tilman 1989). Uncontrolled observations on streams whose watersheds simply differ in land cover suffer from problems of inferring cause from correlational patterns that are confounded by numerous uncontrolled and cross-correlated variables, large cross-site variability, and time lags (compare Pickett 1989). Finally, mechanistic models linking land cover to ecological responses of stream ecosystems can be difficult to construct (Nilsson and others 2002) and may be burdened with large and unknowable errors. Thus, no single scientific approach is adequate for understanding the effects of changing land use on stream ecosystems. Despite the difficulties with each of these approaches, each clearly can contribute significant (and sometimes unique) information about how land-cover change affects stream and river ecosystems. Ultimately, satisfactory understanding of the ecological effects of land-cover change in streams and rivers, like many complex ecological problems, probably will require a creative combination of these approaches (compare Carpenter 1998).
One approach that can help us understand how changes in land cover affect stream ecosystems is empirical modeling (see, for example, Omernik 1976; Peters 1986; Cole and others 1991; Shipley 2000). Empirical models can be constructed using either data collected expressly for the model or suitable data collected for other purposes and appropriated for use in the model (“third-party models”). The potential of this latter class of models is growing rapidly as environmental monitoring and research programs grow and as large data sets are posted over the Internet.
An important issue with empirical (and other) models is spatial scaling of variables. Stream ecosystems are affected by processes occurring at different spatial scales, from local shading caused by the canopy directly over a place on the streambed to regional loading of materials from distant parts of the water- or airshed. As tools for the analysis of Geographic Information System (GIS) become more widespread and sophisticated, it will increasingly become possible to choose the scale at which to model these effects to optimize predictive power and understanding.
Two particular scaling issues commonly confront scientists interested in the effects of land cover on stream ecosystems. The first (which we call “spatial extent”) is the effect of watershed size on the predictive power of empirical models. Large watersheds usually contain many different land-cover patches. The response of the stream is determined jointly by many landscape elements, so nonspatially explicit variables, such as the percentage of the watershed in a given land-cover class, may be adequate predictors of land-cover effects on streams and rivers. In very small watersheds, though, the number of land-cover elements usually is small, so the idiosyncrasies of the spatial arrangement or management of such individual elements may have strong effects on stream ecosystems. Thus, we expect the predictive power of models based on nonspatially explicit landscape variables to decline in watersheds below some threshold size, at which point the characteristics of individual landscape elements become important.
The second issue has to do with the distribution of land cover within the watershed. Presumably, landscape elements near the stream or river have more influence on its ecosystem than elements in a distant part of the watershed. Although land cover often is assessed for the entire watershed, it is equally possible to assess land cover in the riparian corridor or local area around a sampling point to try to account for this increased influence of near-stream landscape elements. Such alternative assessments of land cover (which we refer to as “spatial perspective”) may have different predictive power for different ecological response variables in the stream or river.
We used large, third-party data sets to construct empirical models of the effects of land-cover change on stream ecosystems in the mid-Atlantic region. Our goals were to answer the following questions:
-
1
Can simple empirical models based on readily available data be used effectively to describe ecological responses to different land covers?
-
2
How does using different spatial extents and spatial perspectives affect our ability to describe the ecological responses of streams to different land covers?
Methods
Choice of study area and variables
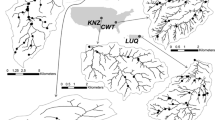
Our study required an area with diverse land cover for which large data sets of interesting ecological variables were available. We chose the Chesapeake Bay watershed in the eastern United States (Figure 1) because it is a large area with diverse and changing land cover (Table 1) and has been well studied by ecologists. We chose the following ecological response variables: fish species richness, the percentage of fish species that are non-native, annual flux of nitrate–N, species richness of benthic macroinvertebrates, cover and species richness of aquatic plants, and canopy cover by riparian vegetation. These variables are of interest to scientists and managers and are likely to exhibit different kinds of responses to land cover.
Ecological response variables
We obtained ecological data from state and federal agencies (via the Internet, CD-ROMs, and personal communication with agency personnel) and from the literature. We recorded our effort to obtain, collate, and process the data to help assess the feasibility of using these large third-party data sets in research. Different sampling points were used for each of the ecological response variables, so our analyses of the different ecological variables were run separately.
Nitrate flux
Nitrate data were obtained from four sources. Two of these were the US Geological Survey: DDS-37, which includes data from selected USGS National Stream Water Quality Monitoring Networks (WQN; Alexander and others 1996), and the National Water-Quality Assessment database (USGS 2000a). We also queried the US Environmental Protection Agency’s STORET database (USEPA 2000a), with assistance from STORET staff, and used data from (Jordan and others 1997). Some of these data were nitrate–N + nitrite–N, while others were only nitrate–N. We used both of these data interchangeably, assuming that nitrite–N was a small fraction of the total flux (Meybeck 1982). All sites were sampled at least once a month for a year and had data on daily water discharge. Nitrate flux was calculated as the product of nitrate concentration (monthly means) and water discharge. In all, we acquired data on nitrate flux for 110 sites that were sampled between 1989 and 1999.
Plants
The Maryland Biological Stream Survey (MBSS), a program of the Maryland Department of Natural Resources (MDNR), provided data on species richness of aquatic plants for 269 sites in that state (MBSS 2000); we randomly subsampled these sites from the larger MBSS database. Data were collected between 1995 and 1997 between 1 June and 30 September (Mercurio and others 1999). The USEPA’s Environmental Monitoring and Assessment Program [EMAP; including data from the Mid-Atlantic Integrated Assessment (MAIA)] collected data on cover of aquatic plants and riparian vegetation for 165 sites in Delaware, Maryland, Pennsylvania, Virginia, and West Virginia (USEPA 1999a). Data were collected between 1993 and 1995 during April to mid-June (USEPA 1999b). Cover of all macrophytes (excluding macroalgae) was visually estimated along eleven 10-m-wide transects across the stream as absent, sparse (less than 10%), moderate (10%–40%), heavy (40%–75%), or very heavy (greater than 75%). A site mean cover was calculated by assigning cover class midpoint values to each transect and taking the simple mean of the 11 transects. Further details on sampling and analysis were presented by (Lazorchak and others 1998) and (Chaloud and Peck 1994).
Benthic invertebrates
The USEPA EMAP database also included data on species richness of benthic macroinvertebrates for the same sites and times as for plant cover (see above). Sampling for the MAIA program occurred primarily near road crossings using standard USEPA rapid assessment techniques for sampling riffles (Plafkin and others 1989) in the Piedmont ecoregion and modified USEPA methods for lowland coastal plain streams (USEPA 1997). Both methods integrate multiple D-net “jabs” stratified by habitat availability in shallow wadeable habitats only (not pools). The first 100 macroinvertebrates were removed from random subsamples and identified to genus or family. We used data for 303 sites at the family level to avoid confounding land-use effects with local differences in community structure resulting from differences in species geographic ranges. Further details on sampling and analysis were presented by (USEPA 1997), (Lazorchak and others 1998), and (Maxted and others 1999).
Fish
We analyzed fish species richness and the percentage of fish species that were non-native (using the criteria of Hocutt and others 1986) at 944 sites. Data were obtained from several sources. The USEPA EMAP program (USEPA 1999a) sampled streams using a single pass with an electrofisher in multiple habitats throughout the stream and provided data for 165 sites. Data were collected between 1993 and 1995 during a two-month sampling window from April to mid-June. Further details on sampling and analysis for the EMAP program were presented by (Lazorchak and others 1998) and (Chaloud and Peck 1994). Data for 33 sites in Delaware were obtained from the Delaware Division of Fish and Wildlife (Mr. Craig Shirey, Delaware Division of Fish & Wildlife, personal communication). Fish were sampled with a backpack electrofisher in the summer months of 1986, 1988, 1989, and 1990. The Maryland Biological Stream Survey (MBSS 2000) provided data for 269 sites in that state (we randomly subsampled these sites from the larger MBSS database). Data were collected between 1995 and 1997 from 1 June to 30 September using two electrofishing passes (Mercurio and others 1999). The Pennsylvania Fish and Boat Commission (PFBC) provided data on fish communities from Pennsylvania (PFBC 2000). Data were collected between 1975 and 1995 during annual summer fish surveys. Sampling gear type was not always included in the database, but, where mentioned, a form of electrofishing was used (electrobackpack, electrotowboat, day or night electroboat). We randomly selected 290 sites from this large database. The PFBC (2000) data set is censored to remove rare and endangered species; we determined that our study sites did not contain any of these species and was therefore complete (David Argent, Pennsylvania State University, personal communication). Data for Virginia (from 1985 to 1990) were obtained from the Virginia Department of Game and Inland Fisheries (Shelly Miller personal communication). We randomly chose 187 sites from this database.
All ecological variables were sampled in free-running reaches, not in impoundments. Therefore, our analysis underestimates the impact of impoundments to the extent that impounded reaches differ from the free-running reaches that they replaced.
Predictor variables
Land-cover data
For each point where ecological data were available, we derived information on land cover, dams, and point sources of pollution. For this process, GIS layers for the various ecological data sets were created using the reported latitude and longitude for each sample site. Land-cover data were calculated from the National Land-Cover Data (NLCD) and were obtained from the Multi-Resolution Land Characteristics Consortium (MRLC) (USEPA 2000b). These data were developed from 30-m Landsat thematic mapper data ranging from 1986 to 1994. We aggregated the NLCD land-cover categories into seven land-cover classes (Table 2). We overlaid the NLCD GIS layer over the watershed, stream corridor, and local boundaries to determine land cover within each of these areas. The land-cover variables were calculated for three spatial perspectives: watershed, stream corridor, and local (Figure 2). The watershed perspective included land cover in the entire watershed of each sampling site, regardless of the size of the watershed. To obtain this perspective, we delineated watersheds using 90-m Digital Elevation Models (DEM) obtained from the USGS (USGS 2000b). Using the National Hydrography Dataset (USGS 2000c), the DEMs were modified to include known stream locations (Moglen and Beighley 2000). Then, knowing elevation and applying the rule that water flows in the direction of the steepest downhill gradient, it is possible to infer flow directions, flow lengths, slopes, drainage area, and watershed boundaries (see, for example, O’Callaghan and Mark 1984; Jensen and Domingue 1988; Tarboton and others 1991). For example, the drainage area of a watershed was calculated as the number of cells draining to the sampling site times the area of an individual cell (0.0081 km2). Although only a few of the ecological data sets included drainage area for assessing our GIS-derived areas, a similar application by (Beighley and others 2002) showed that the delineation procedure we used resulted in approximately ± 5% error in drainage area for watersheds ranging from 14 to 67,000 km2 in the Chesapeake Bay Watershed. The stream corridor perspective included land cover within the pixel containing the stream plus one pixel on either side of the stream, resulting in a 270-m-wide window centered on the stream. The local perspective included land cover within a circle (300-m radius) centered on the sample site.
In addition, the DEM elevation and ecoregion (Omernik 1987) at each sample site were determined. Ecoregions also were aggregated into major physiographic provinces (that is, coastal plain, piedmont, or uplands).
Dam data
We obtained information on dams from the National Inventory of Dams, a program of the US Army Corps of Engineers (NID 2001). This is the most comprehensive listing of dam information for the United States and includes information on 77,000 dams in the United States and Puerto Rico. The NID includes a dam only if it is in a high or significant hazard class, if it exceeds 7.6 m in height and 18,500 m3 in storage, or if it exceeds 1.8 m in height and 62,000 m3 in storage; consequently many small dams are excluded. For each sampling point, we calculated the density of dams (number/km2) in the watershed, the closest dam upstream of the study site based on hydrologic flow path length, and the number of dams in the watershed weighted by their inverse distance squared from the study site. Additionally, two “storage” variables were calculated from the NID database: maximum storage and live storage (the difference between maximum and normal storage—a measure of the ability of the reservoir to alter hydrology downstream). Both of these storage variables were summed over all reservoirs upstream of the study site and normalized by the area of its watershed.
Point-source data
We also used point-source data as predictor variables. The point-source data used in the study were obtained from USEPA BASINS database (USEPA 2000c): Permit Compliance Systems, Industrial Facilities Discharge Sites, Toxic Release Inventory, National Priority List Sites, and Hazardous and Solid Waste Sites. These databases include a wide range of point sources, including permitted industrial and municipal discharges, wastewater treatment plants, landfills, Superfund sites, and locations of handlers of hazardous wastes. We interpreted this point-source information as a general measure of industrial activity in the watershed. We calculated three point-source variables: the density (number/km2) of point sources in the watershed, the closest point source upstream of the study site based on hydrologic flow path length, and the number of point sources weighted by their inverse distance squared from the study site. Table 1 shows the range for all predictor variables in our data sets.
Approach to statistical analysis
Our basic statistical tool was stepwise multiple linear regression, with forward selection of variables and p = 0.15 to enter or remove variables. Cases with missing data were excluded (that is, listwise exclusion). Statistical analyses were done using SYSTAT 8.0, SYSTAT 9.0, Statistica 5, and SPSS 10.0; comparisons among these packages showed that they gave comparable results.
We ran a series of models (Figure 3) to examine the relationships between land-cover and hydrological variables and our ecological response variables. In addition to the predictor variables mentioned below, we used stream size (as log10 watershed area), elevation at the study site, ecoregion, latitude, and longitude as covariates, as appropriate. Data were transformed prior to analysis as needed. First, we examined the effectiveness of adding complexity to our models by regressing the ecological response variables against (1) land cover only, (2) land-cover + dam variables, and (3) land-cover + dam variables + point-source variables, and comparing the resulting models. Second, we examined the effectiveness of assessing land cover from three different spatial perspectives. Here, we ran regression models using (a) land cover of the watershed, (b) land cover of the stream corridor, or (c) local land cover around the study site as predictor variables. Third, we examined the effect of spatial extent on predictive power by running regressions of the ecological response variables on land cover separately for watersheds of different sizes. We divided our data set into subsets of equal size (but with n > 50) by watershed area and ran our analysis separately on each subset.
We assessed the importance of our predictor variables in two ways. First, we simply examined the slopes for all variables that were included in our stepwise models. These slopes show the change in an ecological response variable that is expected from a unit change in the predictor variable. However, because predictor variables vary over different ranges (for example, the percentage of forested land in a watershed varies over a much greater range than the percentage of open water for most watersheds), these slopes do not express the change in an ecological response variable that might be expected from changes in land cover. As an index of the amount of variation to be expected in each independent variable, we took the difference between the 5th and 95th percentiles for each independent variable in our data set. We then multiplied this number by the regression slope to calculate the expected variation in an ecological response variable that might be induced by likely changes in each independent variable. We call this quantity “scope for change.”
Correlations between independent variables affect the interpretation of our models in two ways. First, the percentages of land in major land-cover classes are inevitably negatively correlated with one another. In our data sets, % forest was negatively correlated with % pasture and % cultivated land with r 2 = 0.25–0.76, depending on spatial perspective. Thus, a significant positive relationship between % forest and an ecological response variable might also be interpreted as evidence of a negative relationship between that variable and % pasture or % cultivated land. Second, the percentage of land in a given land-cover class was correlated across spatial perspectives, particularly between the watershed and stream corridor perspectives. Thus, the % forest in the entire watershed was correlated with the % forest in the stream corridor. Consequently, even if land cover at only one spatial perspective were truly related to an ecological response variable, we would observe weaker correlations at other spatial perspectives.
Results
Costs of analysis
Because the data used in this study were available free of charge, our only costs were labor. It took 60–65 person-days to find and assemble the data sets (Table 3). Three tasks were especially time-intensive. First, it took about 18 person-days of searching the Internet, telephone calls, and emails to find appropriate data. This task was eased because many of the people contacted were helpful and knowledgeable, but it was hampered because none of us was very familiar with the study area before undertaking this work. Second, because data from different agencies came in different formats, we spent considerable time bringing the data together into a form that we could use. Third, we devoted several days to determining the appropriate DEM resolution and developing the drainage network for the study region. Our final analyses were done using a DEM pixel resolution of 90 m. Initially the analyses were performed using 300-m pixels, which resulted in poor inferred drainage networks at small drainage areas (less than 30 km2). Because many of the sampling sites had small drainage areas and one of the study goals was to investigate the effects of drainage area on predictive power, the 300-m pixel resolution was too coarse. Although the study region has complete coverage at 30 m, the computational time and data storage required to use this higher resolution data would have been prohibitive.
Identity and effectiveness of predictor variables
The regressions generally had low-to-moderate coefficients of determination (Tables 4, 5). Land-cover, dam, and point-source variables together accounted for 26-100% of the overall R 2 of these models. Despite the modest contribution of such variables in predicting ecological responses, the estimates of regression slopes often were fairly precise (Figure 4).
Several land-cover variables affected ecological response variables (Figure 4). A high proportion of cultivated land was associated with high nitrate flux, low fish species richness, and a high cover of aquatic plants. The other agricultural land cover (pastures) had quite different effects and was associated with high fish species richness, not with high nitrate flux. Wetlands were consistently associated with high fish species richness, a low percentage of non-native fish species, and high aquatic plant species richness. Forested land was negatively correlated with nitrate flux and the proportion of non-native fish species and, not surprisingly, positively correlated with riparian plant cover. The percentage of open water appeared to have large effects on nitrate flux (negative), % non-native fish species (positive), and canopy cover (negative), although these correlations were imprecise. Urban land cover was positively associated with the proportion of exotic fish species and riparian canopy cover and negatively associated with macroinvertebrate species richness.
Given the differences in % coverage by each of the land-cover classes, it appears that changes in % forest, % wetland, and % cultivated land all have great potential to change the ecological response variables that we studied (Figure 4, right-hand panels). Changes in % open water, % urban land, and % pasture also have the potential to greatly affect some ecological response variables.
Adding dam- and point-source variables to our models increased predictive power by less than 5% (Table 4). Fish species richness was slightly elevated near dams and in watersheds with many dams (data not shown). Riparian plant cover was weakly associated with low amounts of storage capacity and high amounts of live storage capacity in the watersheds. Watersheds with a high density of point sources tended to have reduced species richness of fish and macroinvertebrates (data not shown), but this effect was weak (Table 4).
Effects of spatial perspective
The spatial perspective at which land cover was assessed affected the effectiveness of empirical models in different ways for the different response variables studied (Table 5). Land cover over the whole watershed best predicted nitrate flux and aquatic plant species richness. Fish species richness and aquatic plant cover were most closely related to land cover in the entire watershed or riparian corridor. In contrast, riparian plant cover was best predicted by local land cover. Land cover in the riparian corridor was the best predictor of macroinvertebrate species richness (Table 5). Thus, each spatial perspective was optimal for at least one of our response variables.
Effect of watershed size (spatial extent)
There was no general relationship between watershed size and the predictive power of empirical models that held for all of the variables studied (Figure 5). For some of our response variables (nitrate flux and plant cover), we had too few data to test for a clear pattern. (The apparent decline in predictive power for aquatic plant cover in large watersheds is an artifact because none of the larger streams had any aquatic plants.) Nevertheless, all three of the variables for which we had many data (plant, macroinvertebrate, and fish species richness) showed a decline in predictive power in very small watersheds (less than 1–10 km2).
Comparisons among ecological response variables
As hypothesized, the ecological response variables we studied had a wide range of associations with the predictor variables. Different variables were the best predictors for different ecological responses. Thus, % cultivated land was the best predictor for nitrate flux, while % wetlands was the best predictor for aquatic plant species richness (Figure 4). Likewise, the spatial perspective that was most effective in predicting ecological responses differed across variables (Table 5). Finally, the predictive power of different models varied widely (Tables 4, 5). On the other hand, some patterns were consistent across ecological response variables. Thus, dam- and point-source variables were ineffective predictors of ecological responses and added little to our models (Table 4). Further, to the extent that our data set allowed us to discern scale dependence of predictive power, it appears that predictive power may decline at small spatial extents for some variables (Figure 5).
Discussion
We were able to construct empirical models that shed light on the effects of land cover on stream ecosystems and on the influence of spatial scaling on our perception of those effects. The specific nature of these effects varied considerably across ecological response variables.
Performance of the empirical models
Review of model characteristics.
Land cover of the entire watershed, particularly the percentage of the watershed in cultivated land, forest, and open water, was the best predictor of nitrate flux (Figure 4 and Table 5). These correlations presumably are a result of high loadings and low retention of nitrogenous fertilizers in cultivated fields, contrasted with lower anthropogenic inputs and high retention of nitrate in forests and open water. These results agree with previous attempts to predict nitrogen flux from large watersheds (for example, see Jordan and Weller 1996; Caraco and Cole 1999; Galloway 2000; Vörösmarty and Sahagian 2000). Models based on land cover in the 270-m-wide streamside corridor were nearly as good as watershed-wide models in predicting nitrate flux (Table 5). In such streamside models, wetland cover was added as a predictive variable (negatively correlated with nitrate flux), suggesting that wetlands close to the stream channel may be biogeochemical hot spots for nitrogen cycling (Lowrance and others 1984; Peterjohn and Correll 1984). Models based on local land cover were poor at predicting nitrate flux (Table 5), suggesting that nitrate is readily moved from distant parts of the watershed.
Aquatic plant species richness was readily predicted from the wetland cover of the watershed (Tables 4, 5 and Figure 4), suggesting the importance of wetlands in supplying propagules to the stream community. In contrast, aquatic plant cover was correlated only weakly with the predictor variables. Perhaps this weak correlation was due in part to the inadequacy of a single measurement of plant cover, which presumably is temporally variable as a result of floods and seasonal development of the vegetation (Alvarez–Cobelas and others 2001). The positive association between aquatic plant cover and % cultivated land may be a result of increased light and nutrients in these streams (Hansen and others 2001). The positive association between riparian plant cover and local forest cover probably arose because the riparian trees themselves were categorized as forest in the NLCD.
For macroinvertebrate species richness, the most effective spatial perspective was the streamside corridor, with urban, forest, and wetland cover being important (Figure 4 and Table 5). Several studies (Richards and others 1997; Fitzpatrick and others 2001; Sponseller and others 2001) likewise have found that reach-scale features were better predictors than watershed-scale features of macroinvertebrate community structure in streams. Many authors (for example, Cummins 1993a; Maridet and others 1998; Yeates and Barmuta 1999; Quinn and others 2000) have found that streamside vegetation is important to macroinvertebrate communities in streams through its provision of leaf litter, shading of the channel, and woody debris.
The spatial perspective at which land cover was assessed did not have a strong effect on the effectiveness of our models of fish species richness (Table 4), in contrast to some previous work suggesting that watershed-scale processes were most important (Roth and others 1996). Richness was positively related to cover of pasture and wetlands and negatively related to cover of cultivated land. Many studies (for example, Roth and others 1996; Walser and Hart 1999; Cuffney and others 2000; Fitzpatrick and others 2001) have shown that a high cover of cultivated land results in low fish species richness, probably a result of high loading of sediments, a lack of instream and streamside cover, and alterations to thermal and hydrological regimes. Wetlands also have been shown to have a positive effect on fish species richness in streams (Roth and others 1996).
This brief review of model characteristics suggests several general points. First, the most effective predictor variables and spatial perspectives varied widely across ecological response variables. Generally, the predictors and spatial perspectives best used to predict each variable were readily interpretable in light of the ecological mechanisms thought to control that variable. To the extent that each response variable is controlled by its own distinct set of mechanisms, there will be no single optimal structure or scale to empirical models of the effects of land use on stream ecosystems. Fitzpatrick and others (2001) reached a similar conclusion in their study of the effects of environmental variables assessed at different scales on fish, macroinvertebrates, and algae in Wisconsin streams.
Second, two land-cover variables had strong effects on several ecological response variables. In our region, wetlands were associated with high species richness of fish and aquatic plants and low richness of non-native fish (Figure 4). Cultivated lands were correlated with high nitrate flux, low fish species richness, and high cover of aquatic plants. Notably, the other agricultural land cover (pasture) had none of these effects. Thus, wetlands and cultivated fields had strong influences on the overall structure of stream ecosystems in the Chesapeake Bay watershed.
Finally, dam- and point-source variables were consistently ineffective in predicting ecological response variables (Table 4). Certainly, point-source pollution is well known to have strong effects on many aspects of stream ecosystems (Mason 1996). However, our point-source variables may have been ineffective predictors because this class contained a wide range of sources with highly varied historical and current effects on stream ecosystems. Likewise, dams undoubtedly have strong, wide-reaching effects on streams and rivers (Petts 1984; Rosenberg and others 1995; Nilsson and Berggren 2000).
Three factors may be behind the weakness of dam variables in our models. First, the sampling programs that produced the data we used deliberately avoided sampling in impoundments. Ecological conditions in impoundments usually are radically different than those in free-flowing reaches (Nilsson and others 1997; Jansson and others 2000b). As a result, the omission of impoundments from the sampling programs led us to underestimate the impacts of dams. Second, the National Inventory of Dams is incomplete and does not contain information on all of the numerous small dams in the study area. Third, our characterization of dams was very coarse and, in particular, contained no information on how the dams actually altered hydrology, temperature, water chemistry, or sediment dynamics of reaches downstream. A more complete characterization of dams, along with information on ecological conditions in impoundments, probably would have led to different conclusions than our models suggested about the effects of dams.
Adequacy of model power.
The power of our models can be judged by two criteria: the R 2 of the model and the precision with which regression slopes are estimated (for example, the standard error or coefficient of variation of the slope). The former criterion is a measure of how much variation in a response variable is accounted for by the predictor variables; it is relevant if the model is to be used to predict an ecological variable at a specific site. The latter is a measure of the precision with which land-cover effects are known; it is relevant if one is trying to describe the average change in an ecological response variable that is expected from a unit change in the predictor variable.
Because our models usually had R 2 of 0.05–0.5 (Tables 4, 5), they rarely will be adequate to predict ecological conditions at a given site. On the other hand, slopes often were estimated with good precision (one third of the slopes shown in Figure 4 have CV < 25%), and may be quite adequate for specifying the average amount of change in an ecological response variable that is associated with a specified difference in land cover. Many empirical models in ecology have similar characteristics, with good ability to quantify general trends but poor ability to predict individual data points (Peters 1983, 1986; Brown 1995).
Several factors may underlie the imprecision of our models. First, large measurement error is associated with some of the variables. For example, 25% of the variation in fish species richness is accounted for by differences in estimates of fish species richness made a few days apart. Thus, the maximum R 2 possible for a model based on a single measurement of fish species richness (our models were based on such single measurements) at each site is 0.75. We do not have such direct estimates of measurement error in other variables, but it probably was substantial.
Second, our land-cover classes (and other predictor variables) are not homogeneous. What we called “forested” land comprised forests with different species compositions on different soils and included such diverse habitats as quasinatural forests, plantations, and urban woodlands. Other land-cover classes are similarly heterogeneous. Likewise, our characterization of impoundments by just a few variables (Table 1) did not distinguish between peaking hydropower operations and flood control reservoirs, if they had the same live storage capacity. This incomplete characterization of dams may have led to our failure to detect the strong effects of dams on ecological response variables.
Third, although the study area was chosen to be relatively homogeneous in terms of climate and biota, all large study areas are inevitably heterogeneous. Large-scale heterogeneity within the study area may have introduced variance not accounted for by our models. Where the data sets were large enough (fish and macroinvertebrates), we subdivided the data by physiographic province (coastal plain, piedmont, or uplands) and ran separate models for each ecoregion. Although this subdivision into presumably more homogeneous study areas did not consistently increase R 2 (Table 6), heterogeneity within our study region probably was an important source of error.
Fourth, although the data sets we analyzed were large, our analyses were clearly limited by the size of the available data sets. The relatively large standard errors of the nitrate flux model (Figure 4), despite its high R 2 (0.55), reflects the effect of the small size (n = 110) of the data set on nitrate flux. Having a larger data set would increase the precision of slope estimates and may have allowed for an increase in R 2 by subdividing the data set. Furthermore, if we had had a larger data set, we could have investigated the relationships between watershed size and predictive power (Figure 5) in more detail.
Finally, our approach was basically a “snapshot,” with the assumption that measured land-cover variables had an instantaneous effect on ecological response variables. There are at least two problems with this assumption. There may be very important time lags in the expression of land-cover changes on stream ecosystems. Thus, the effects of land-cover change may affect stream ecosystems for decades. In fact, Harding and others (1998) found that past land cover was a better predictor of present-day diversity than current land cover. Also, the data used in our analyses were not actually collected contemporaneously but rather somewhere in the period 1986–1994, which could have introduced some error. The former problem probably is more serious than the latter.
Several strategies could improve the predictive power of empirical models linking land cover and stream ecosystems. First, larger data sets could improve the precision of regression slopes, although not necessarily R 2 of the models. Second, making land-cover classes more homogeneous through disaggregation of very heterogeneous classes could make empirical models more predictive, as well as provide insight into the characteristics that give land covers their effects on streams. Similarly, better use of covariates or blocking within large, heterogeneous study areas could remove unwanted variance from the models. Both of these last two strategies require large data sets to be effective. Finally, further exploration of the use of multiple temporal scales (we used multiple spatial scales in this study) could significantly improve empirical models in the many situations where temporal lags are important. Nevertheless, although all of these strategies could significantly improve the performance of empirical models, we doubt that they will improve the empirical models enough to allow the precise forecasting of conditions at a particular site.
Domains of models
Even when empirical models have adequate predictive power, they are valid over only a limited domain of conditions (domains of scale in the sense of Wiens 1989). Our models, for instance, could reasonably be expected to apply to streams and rivers in the mid-Atlantic states east of the Alleghenian Divide, and they may systematically fail outside of this region because of differences in the regional species pool, environmental conditions (climate, soils), or details of land use within a land-cover class (for example, amount of fertilizer used on cultivated fields in different regions). Furthermore, our models were based on a particular range of values of predictor variables (Table 1) and may not be reliable when extrapolated beyond this range. Finally, if these models are used to forecast future impacts of land-cover change, there is an assumption of uniformitarianism—that the processes that operated in the past to produce the patterns we observed will continue to operate unchanged in the future. This assumption will be violated if, for example, residents begin using new pesticides on their lawns or forest species composition changes because of the arrival of an exotic tree species. Because of this last assumption, empirical models such as ours must be used cautiously when forecasting the future.
Influence of spatial extent and perspective
The spatial perspective at which land cover was assessed had strong and variable effects on the performance of our empirical models (Table 5 and Figure 5). The most effective spatial perspective differed among the ecological response variables and was linked to the mechanisms thought to control each variable. Thus, broad spatial perspectives (watershed or streamside corridor) were most effective in estimating the flux of nitrate, which is thought to move readily through watersheds. The importance of wetland cover assessed at broad (watershed) perspectives to aquatic plant species richness agrees with recent studies (see for example, Nilsson and others 1993), suggesting that plant propagules can be transported over long distances, even during single flood events. In contrast, local or streamside corridor perspectives were useful in estimating macroinvertebrate species richness and riparian plant cover, which may be controlled by more local processes such as shading and loading of leaf litter and wood into the channel and provision of egg-laying sites (Cummins 1993a, 1993; Gore and others 1995; Maridet and others 1998; de Szalay and Resh 2000). The relative insensitivity of fish species richness to spatial perspective suggests that processes operating at all three spatial scales are important (compare Fitzpatrick and others 2001). Examples include hydrological influences (watershed scale), loading of organic matter, including allochthonous forage items (streamside corridor scale), and canopy and streamside cover and local channel form (local scale).
The effect of watershed size (spatial extent) was less clear. For at least three of our response variables, predictive power appeared to drop off to nearly zero in very small watersheds (less than 1–10 km2) (Figure 5), supporting the idea that nonspatially explicit characteristics of watersheds are inadequate predictors at very small spatial scales. Given that the average size of an individual landscape patch in our study area was 0.3 km2, a typical watershed of 3 km2 contained only ten patches. It is reasonable to guess that the spatial arrangement of such patches will matter when so few are present. In addition, our ability to predict aquatic plant cover rose dramatically (from R 2 of 0.07 to as high as 0.47) when we split the data set into watersheds of different sizes. Other variables did not show a clear pattern with watershed size, but our ability to discern such patterns was limited by inadequate data, even in our relatively large data sets. Thus, we still cannot specify the watershed size at which the mere percentage of land covers does not provide predictive value and the spatial arrangement of landscape elements must be considered. This size might vary from one region to another (especially as patch size varies across regions) and from one response variable to another.
Alternatively, predictive power may decline with watershed size because of the increasing importance of organismal dispersal in small watersheds. In a large river, the dispersal range of an organism may be small compared to the scale of land-cover influences; a site 1 km downstream of an agriculturally dominated site is also likely to be agriculturally dominated. In contrast, in a small watershed, the same dispersal distance may carry an organism into a very different environment, in terms of land-cover influences. Thus, in small watersheds, local dispersal may homogenize the distributions of organisms relative to land-cover patterns.
Conclusions
Third-party empirical models based on large data sets clearly can contribute to understanding the effects of land cover on stream ecosystems. Our models revealed strong relationships between land cover and a wide range of ecological variables. The models further suggested that the spatial scales over which land cover and ecological variables were associated varied widely, depending on the mechanisms thought to link land cover and each ecological variable.
The potential of large, third-party empirical models probably will grow rapidly over the coming decade. More and more ecological data sets are being compiled and posted via the Internet. GIS data and software are improving in several important ways (Nilsson and others 2003). Land-cover and digital elevation models will become available at finer spatial scales, allowing scientists to explore the use of fine spatial perspectives. Finer spatial resolution may also allow for more precise distinctions between land covers that are now classed together in heterogeneous cover classes. As land-cover maps are routinely updated, it will be easier to evaluate the importance of temporal change in land cover (Harding and others 1998). Finally, we expect GIS software to become more powerful and easier for nonspecialists to use. For instance, we had to develop the drainage network for the entire Chesapeake Bay watershed, which accounted for a large portion of our total effort. Future software may better unify the relationship between watershed data management and applications. One form of this concept is referred to as “Geo-Object” modeling (Davis and Maidment 1999), in which the various spatial data (DEM, land cover, stream network) are organized as objects in a database that can be extracted with minimal effort through object-oriented GIS applications. This system will reduce the demanding preprocessor work required to develop predictor variables for empirical models. All of these improvements are likely to result in incremental, quantitative improvements in the predictive power of empirical models, rather than a fundamental qualitative shift in the utility of such models.
Broadly, there are three ways in which empirical models may relate to more mechanistic models of the effects of land-cover change on stream ecosystems. Empirical models may replace more mechanistic models if there is inadequate information or resources to build reliable mechanistic models (compare Nilsson and others 2003). However, without an explicit consideration of mechanism, empirical models may fail to give reliable predictions if they are taken outside their domains (see above). Alternatively, empirical models may be used to constrain mechanistic models when both are applied to the same conditions. The fact that empirical models have such a strong grounding in actual field conditions may make them useful in testing various mechanistic models. Finally, empirical models may be combined with mechanistic models as part of a comprehensive program to understand the ecological effects of land-cover change on streams and rivers. Benda and others (2002) considered the problem of predicting ecological change from economic or land-use change as a logical chain of several steps. Some of these steps may be empirical models, while others may be mechanistic models, depending on the predictive requirements and scientific understanding associated with each step. Thus, empirical modeling has the potential to play several important roles in the understanding of the effects of land-cover change on running water ecosystems.

Map of the study area.

Diagram showing the three different spatial perspectives at which land cover was assessed: a watershed, b stream corridor, c local.

Flow chart showing the major steps in data collection and analysis.

Influence of land-cover variables on ecological response variables, assessed as the slope (partial regression coefficients) of the regression model and the “scope for change.” “Scope for change” is the regression slope multiplied by the amount of variation to be expected in the predictor variable (see Methods). Only variables that were significant in the stepwise multiple regression models are included. Results are for the spatial perspective with the highest R 2 (see Table 5).

Prediction of ecological response variables for different watershed sizes and spatial perspectives. Plots show the total R 2 of the models using all variables. Watershed size is given for the midpoint of each watershed-size class.
References
Alexander RB, Ludtke AS, Fitzgerald KK, Schertz TL. 1996. Data from selected U.S. Geological Survey National Stream Water-Quality Monitoring Networks (WQN) on CD-ROM, USGS Open-File Report 96–337 (report.text revised 1/15/97), Reston, Virginia
M Alvarez–Cobelas S Cirujano S Sanchez–Carrillo (2001) ArticleTitleHydrological and botanical man-made changes in the Spanish wetland of Las Tablas de Daimiel. Biol Conserv 97 89–98 Occurrence Handle10.1016/S0006-3207(00)00102-6
RE Beighley DL Johnson AC Miller (2002) ArticleTitleA subsurface response model for storm events within the Susquehanna River Basin. ASCE J Hydrol Eng 7 185–91 Occurrence Handle10.1061/(ASCE)1084-0699(2002)7:2(185)
LE Benda NL Poff C Tague MA Palmer J Pizzuto S Cooper E Stanley G Moglen (2002) ArticleTitleHow to avoid train wrecks when using science in environmental problem solving. BioScience 2 1127–1136
J Brown (1995) Macroecology. University of Chicago Press Chicago 269
NF Caraco JJ Cole (1999) ArticleTitleHuman impact on nitrate export: an analysis using major world rivers. Ambio 28 167–70
SR Carpenter (1998) The need for large-scale experiments to assess and predict the response of ecosystems to perturbation. M Pace P Groffman (Eds) Successes, limitations, and frontiers in ecosystem science Springer-Verlag New York 287–312
Chaloud DJ, Peck DV. 1994. Environmental Monitoring and Assessment Program: Integrated quality assurance project plan for the Surface Waters Resource Group, 1994 activities. EPA 600/X-91/080, Rev. 2.00. Las Vegas, NY: US EPA
JJ Cole GM Lovett S Findlay (1991) Comparative analyses of ecosystems: patterns, mechanisms, and theories. Springer-Verlag New York 375
TF Cuffney MR Meador SD Porter ME Gurtz (2000) ArticleTitleResponses of physical, chemical, and biological indicators of water quality to a gradient of agricultural land use in the Yakima River Basin, Washington. Environ Monitor Assess 64 259–70 Occurrence Handle10.1023/A:1006473106407 Occurrence Handle1:CAS:528:DC%2BD3cXntVWjurc%3D
Cummins KW (1993a) Riparian-stream linkages: in-stream issues. In: Bunn SE, Pusey BJ, Price P, editors. Ecology and management of riparian zones. Land and Water Resources Research and Development Corporation, Canberra. pp 5–20
KW Cummins (1993) Invertebrates. P Calow G Petts (Eds) Rivers handbook Blackwell Oxford 234–50
Davis KM, Maidment DR. 1999. A geoobject model for rivers and watersheds. Center for Research in Water Resources On-Line Publication, University of Texas at Austin, http:http://www.crwr.utexas.edu/giswr/resources/library/geoobject.pdf , September 16, 1999
FA de Szalay VH Resh (2000) ArticleTitleFactors influencing macroinvertebrate colonization of seasonal wetlands: responses to emergent plant cover. Freshw Biol 45 295–308 Occurrence Handle10.1046/j.1365-2427.2000.00623.x
M Dynesius C Nilsson (1994) ArticleTitleFragmentation and flow regulation of river systems in the northern third of the world. Science 266 753–62 Occurrence Handle1:CAS:528:DyaK2MXitVGgsL8%3D
FA Fitzpatrick JC Knox (2000) ArticleTitleSpatial and temporal sensitivity of hydrogeomorphic response and recovery to deforestation, agriculture, and floods. Phys Geogr 21 89–108
FA Fitzpatrick BC Scudder BN Lenz DJ Sullivan (2001) ArticleTitleEffects of multi-scale environmental characteristics on agricultural stream biota in eastern Wisconsin. J Am Water Resources Assoc 37 1489–507
JN Galloway (2000) ArticleTitleNitrogen mobilization in Asia. Nutrient Cycl Agroecosyst 57 1–12 Occurrence Handle10.1023/A:1009832221034
JA Gore FL Bryant DJ Crawford (1995) River and stream restoration. J Cairns (Eds) Rehabilitating damaged ecosystems. 2nd ed. Lewis Publishing Ann Arbor 245–75
L Hannah D Lohse C Hutchinson JL Carr A Lankerani (1994) ArticleTitleA preliminary inventory of human disturbance of world ecosystems. Ambio 23 246–50
B Hansen HF Alroe ES Kristensen (2001) ArticleTitleApproaches to assess the environmental impact of organic farming with particular regard to Denmark. Agric Ecosystems Environ 83 11–26 Occurrence Handle10.1016/S0167-8809(00)00257-7
JS Harding EF Benfield PV Bolstad GS Helfman EBD Jones (1998) ArticleTitleStream biodiversity: The ghost of land use past. Proc Nat Acad Sci USA 95 14843–7 Occurrence Handle10.1073/pnas.95.25.14843 Occurrence Handle1:CAS:528:DyaK1cXotVGmur0%3D Occurrence Handle9843977
CH Hocutt RE Jenkins JR Stauffer (1986) Zoogeography of the fishes of the central Appalachians and central Atlantic Coastal Plain. C Hocutt E Wiley (Eds) The zoogeography of North American freshwater fishes Wiley New York 161–211
R Jansson C Nilsson M Dynesius E Andersson (2000a) ArticleTitleEffects of river regulation on river-margin vegetation: a comparison of eight boreal rivers. Ecol Appl 10 203–24
R Jansson C Nilsson B Renöfält (2000b) ArticleTitleFragmentation of riparian floras in rivers with multiple dams. Ecology 81 899–903
SK Jensen JO Domingue (1988) ArticleTitleExtracting topographic structure from digital elevation data for geographic system analysis. Photogrammetr Eng Remote Sensing 54 1593–600
JP Johnes (1996) ArticleTitleEvaluation and management of the impact of land use change on the nitrogen and phosphorus load delivered to surface waters: the export coefficient modeling approach. J Hydrol 183 323–49 Occurrence Handle10.1016/0022-1694(95)02951-6 Occurrence Handle1:CAS:528:DyaK28XmtFOnsb4%3D
TE Jordan DE Weller (1996) ArticleTitleHuman contributions to terrestrial nitrogen flux. BioScience 46 655–64
TE Jordan DL Correll DE Weller (1997) ArticleTitleEffects of agriculture on discharges of nutrients from Coastal Plain watersheds of Chesapeake Bay. J Environ Quality 26 836–48 Occurrence Handle1:CAS:528:DyaK2sXjsVCqtLs%3D
Lazorchak JM, Klemm DJ, Peck DV, editors. 1998. Environmental Monitoring and Assessment Program—Surface Waters: Field operations and methods for measuring the ecological condition of wadeable streams. EPA/620/R-94/004F. Washington, DC: US EPA
RR Lowrance RL Todd J Fail O Hendrickson R Leonard L Asmussen (1984) ArticleTitleRiparian forests as nutrient filters in agricultural watersheds. BioScience 34 374–7
L Maridet JG Wasson M Philippe C Amoros RJ Naiman (1998) ArticleTitleTrophic structure of three streams with contrasting riparian vegetation and geomorphology. Arch Hydrobiol 144 61–85
C Mason (1996) Biology of freshwater pollution. 3rd ed Longman Harlow, UK 356
LL Master SR Flack BA Stein (1997) Rivers of life; critical watersheds for protecting freshwater biodiversity. The Nature Conservancy Arlington, VA 71
Maxted JR, Barbour MT, Gerritsen J, Poretti V, Primrose N, Silvia A, Penrose D, Renfrow R. 1999. Data report: assessment framework for mid-Atlantic coastal plain streams using benthic macroinvertebrates. Report prepared for Mid-Atlantic Integrated Assessment Programs, USEPA Region 3, Fort Meade, MD
[MBSS] Maryland Biological Stream Survey. 2000. http://www.dnr.state.md.us/streams/mbss/mbss_pubs.html, Monitoring and Nontidal Assessment Division, Maryland Department of Natural Resources, 580 Taylor Ave., Annapolis, MD 21401
JM McCloskey H Spalding (1989) ArticleTitleA reconnaissance-level inventory of the amount of wilderness remaining in the world. Ambio 18 221–7
Mercurio G, Chaillou J, Roth N. 1999. Guide to using 1995–1997 Maryland Biological Stream Survey data. Report prepared by Versar, Inc., 9200 Rumsey Rd., Columbia, MD 21045, for Maryland Department of Natural Resources, Tawes State Office Building, 580 Taylor Ave., Annapolis, MD 21401
M Meybeck (1982) ArticleTitleCarbon, nitrogen, and phosphorus transport by world rivers. Am J Sci 282 401–50 Occurrence Handle1:CAS:528:DyaL38Xks1Sntbk%3D
JL Meyer MJ Sale PJ Mulholland NL Poff (1999) ArticleTitleImpacts of climate change on aquatic ecosystem functioning and health. J Am Water Resources Assoc 35 1373–86
GE Moglen RE Beighley (2000) ArticleTitleUsing GIS to determine extent of gauged streams in a region. ASCE J Hydrol Eng 5 190–6 Occurrence Handle10.1061/(ASCE)1084-0699(2000)5:2(190)
RJ Naiman MG Turner (2000) ArticleTitleA future perspective on North America’s freshwater ecosystems. Ecol Appl 10 958–70
[NID] National inventory of dams. 2000. http://crunch.tec.army.mil/nid/webpages/nid.cfm
C Nilsson K Berggren (2000) ArticleTitleAlterations of riparian ecosystems caused by river regulation. BioScience 50 783–92
C Nilsson R Jansson U Zinko (1997) ArticleTitleLong-term responses of river-margin vegetation to water-level regulation. Science 276 798–800 Occurrence Handle10.1126/science.276.5313.798 Occurrence Handle1:CAS:528:DyaK2sXivFOrtL0%3D Occurrence Handle9115205
C Nilsson E Nilsson ME Johansson M Dynesius G Grelsson S Xiong R Jansson M Danvind (1993) Processes structuring riparian vegetation. J Menon (Eds) Current topics in botanical research. Council of Scientific Research Integration. Trivandrum, India 419–31
Nilsson C, Pizzuto JE, Moglen GE, Palmer MA, Stanley EH, Bockstael NE, Thompson LC (2003) Ecological forecasting and urbanizing streams: challenges for economists, hydrologists, geomorphologists, and ecologists. Ecosystems (in press)
JF O’Callaghan DM Mark (1984) ArticleTitleThe extraction of drainage networks from digital elevation data. Comput Vision Graphics Image Process 28 323–44
JM Omernik (1976) The influence of land use on stream nutrient levels. EPA 600/3-76-014. US EPA Washington, DC 105
JM Omernik (1987) ArticleTitleAquatic ecoregions of the conterminous United States. Ann Assoc Am Geograph 77 118–25
[PFBC] Pennsylvania Fish and Boat Commission. 2000. Pennsylvania Spatial Data Access Database (PASDA), http://www.pasda.psu.edu/, Pennsylvania State University, University Park, PA
WT Peterjohn DL Correll (1984) ArticleTitleNutrient dynamics in an agricultural watershed: observations on the role of a riparian forest. Ecology 65 1466–75 Occurrence Handle1:CAS:528:DyaL2MXhtF2nsA%3D%3D
RH Peters (1983) The ecological implications of body size. Cambridge University Press. Cambridge 329
RH Peters (1986) ArticleTitleThe role of prediction in limnology. Limnol Oceanogr 31 1143–59 Occurrence Handle1:CAS:528:DyaL2sXptVOg
G Petts (1984) Impounded rivers. John Wiley New York
STA Pickett (1989) Space-for-time substitution as an alternative for long-term studies. G Likens (Eds) Long-term studies in ecology: approaches and alternatives. Springer-Verlag. New York 110–35
Plafkin JL, Barbour MT, Porter KD, Gross SK, Hughes RM. 1989. Rapid bioassessment protocols for use in streams and rivers: macroinvertebrates and fish. US EPA, Office of Water; EPA/444/4-89-001. Washington, DC: US EPA
SL Postel (2000) ArticleTitleEntering an era of water scarcity: The challenges ahead. Ecol Appl 10 941–8
JM Quinn BJ Smith GP Burrell SM Parkyn (2000) ArticleTitleLeaf litter characteristics affect colonisation by stream invertebrates and growth of Olinga feredayi (Trichoptera: Conoesucidae). NZ J Marine Freshw Res 34 273–87
C Richards RJ Haro LB Johnson GE Host (1997) ArticleTitleCatchment and reach-scale properties as indicators of macroinvertebrate species traits. Freshw Biol 37 219–30
BD Richter DP Braun MA Mendelson LL Master (1997) ArticleTitleThreats to imperiled freshwater fauna. Conserv Biol 11 1081–93
DM Rosenberg RA Bodaly PJ Usher (1995) ArticleTitleEnvironmental and social impacts of large-scale hydroelectric development—who is listening? Global Environ Change Hum Policy Dimens 5 127–48 Occurrence Handle10.1016/0959-3780(95)00018-J
NE Roth JD Allan DL Erickson (1996) ArticleTitleLandscape influences on stream biotic integrity assessed at multiple spatial scales. Landscape Ecol 11 141–56
B Shipley (2000) Cause and correlation in biology: a user’s guide to path analysis, structural equations and causal inference. Cambridge University Press Cambridge 317
RA Sponseller EF Benfield HM Valett (2001) ArticleTitleRelationships between land use, spatial scale and stream macroinvertebrate communities. Freshw Biol 46 1409–24 Occurrence Handle10.1046/j.1365-2427.2001.00758.x
DG Tarboton RL Bras I Rodriguez–Iturbe (1991) ArticleTitleOn the extraction of channel networks from digital elevation data. Hydrol Process 5 81–100
D Tilman (1989) Ecological experimentation: strengths and conceptual problems. G Likens (Eds) Long-term studies in ecology: approaches and alternatives. Springer-Verlag New York 136–57
MG Turner SR Carpenter EJ Gustafson RJ Naiman SM Pearson (1998) Land use. M Mac P Opler P Doran C Haecker (Eds) Status and trends of our nation’s biological resources. Volume 1. National Biological Service Washington, DC 37–61
USEPA. 1997. Field and laboratory methods for macroinvertebrate and habitat assessment oflow gradient, nontidal streams. Mid-Atlantic Coastal Streams Workgroup, Environmental Services Division, Region 3, Wheeling, WV. p. 23
USEPA. 1999a. NHEERL Western Ecology Division Corvallis, OR. http://www.epa.gov/emap/html/dataI/surfwatr/data/mastreams/9396/location/habbest.txt
USEPA. 1999b. NHEERL Western Ecology Division Corvallis, OR. Catalog documentation: EMAP surface waters program level database, 1993–1996 mid-Atlantic streams data, streams habitat data
USEPA. 2000a. Storage and Retrieval (STORET) database, http://www.epa.gov/storet/
USEPA. 2000b. Multi-Resolution Land Characteristics Consortium (MRLC) database, http://www.epa.gov/mrlcpage
USEPA. 2000c. BASINS software and database, http://www.epa.gov/OST/BASINS/
USGS. 2000a. Data on nutrients in the streams, rivers, and ground water of the United States. National Water Quality Assessment Program, http://water.usgs.gov/nawqa/nutrients/datasets/cycle91/
USGS. 2000b. U.S. GeoData Database, http://edc.usgs.gov/geodata/
USGS. 2000c. National Hydrography Dataset (NHD), http://nhd.usgs.gov/index.html
PM Vitousek HA Mooney J Lubchenco JM Melillo (1997) ArticleTitleHuman domination of Earth’s ecosystems. Science 277 494–9 Occurrence Handle1:CAS:528:DyaK2sXkvVektLs%3D
CJ Vörösmarty D Sahagian (2000) ArticleTitleAnthropogenic disturbance of the terrestrial water cycle. BioScience 50 753–65
CJ Vörösmarty BM Fekete M Meybeck RB Lammers (2000a) ArticleTitleGlobal system of rivers: its role in organizing continental land mass and defining land-to-ocean linkages. Global Biogeochem Cycles 14 599–621
CJ Vörösmarty P Green J Salisbury RB Lammers (2000b) ArticleTitleGlobal water resources: Vulnerability from climate change and population growth. Science 289 284–8
CA Walser HL Hart (1999) ArticleTitleInfluence of agriculture on in-stream habitat and fish community structure in Piedmont watersheds of the Chattahoochee River System. Ecol Freshwater Fish 8 237–46
JA Wiens (1989) ArticleTitleSpatial scaling in ecology. Funct Ecol 3 385–97
JT Wootton MS Parker ME Power (1996) ArticleTitleEffects of disturbance on river food webs. Science 273 1558–61 Occurrence Handle1:CAS:528:DyaK28Xls1WqsLc%3D
LV Yeates LA Barmuta (1999) ArticleTitleThe effects of willow and eucalypt leaves on feeding preference and growth of some Australian aquatic macroinvertebrates. Aust J Ecol 24 593–8
Acknowledgements
This work was conducted as part of the “The Ecological Consequences of Altered Hydrologic Regimes” Working Group supported by the National Center for Ecological Analysis and Synthesis, a center funded by the National Science Foundation (NSF) (Grant DEB94-21535), the University of California at Santa Barbara, and the State of California. Additional funding for this project was provided by the Scientific Committee on Water Research (SCOWAR) of the International Council for Science (ICSU); REB and SB were supported by the USEPA STAR (WWS) program. We thank the other participants in the working group for helpful discussions and ideas. Margaret Palmer, Nina Caraco, and two anonymous reviewers provided helpful reviews of the manuscript. We are extremely grateful to everyone who took the time to suggest potential data sources and supply us with data, especially Joyce Boyd, Dan Parker, Craig Shirey, Shelly Miller, Paul Cocca, and David Argent. Finally, we acknowledge the contributions of the people who have worked over many years to collect and analyze samples, enter data, and maintain the databases that made this study possible. This is a contribution to the program of the Institute of Ecosystem Studies and SCOWAR. Although the data described in this article have been funded wholly or in part by the USEPA through its EMAP Surface Waters Program, it has not been subjected to Agency review and therefore does not necessarily reflect the views of the Agency and no official endorsement of the conclusions should be inferred.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Strayer, D., Beighley, R., Thompson, L. et al. Effects of Land Cover on Stream Ecosystems: Roles of Empirical Models and Scaling Issues . Ecosystems 6, 407–423 (2003). https://doi.org/10.1007/PL00021506
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/PL00021506