
Summary
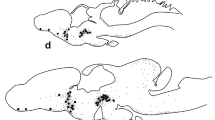
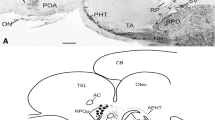
Growth hormone, prolactin and somatostatinlike immunoreactivities were demonstrated in the brains of larval, young adult (parasitic) and upstream migrant adult sea lampreys, Petromyzon marinus, by means of immunoperoxidase techniques. Growth hormone (GH) and prolactin (PRL) were observed within separate perikarya in the nucleus praeopticus, within fibers in the commissura praeinfundibularis, and in nerve endings within the neurohypophysis of larval and adult-stage lampreys. Cell bodies demonstrating immunoreactive growth hormone were more numerous than those reactive for prolactin. Unlike in the upstream migrant adult lamprey, no GH or PRL was demonstrated in the adenohypophysis of larval or parasitic lamprey.
Somatostatin (SRIF)-like immunoreactive neurons were demonstrated in the nucleus commissurae praeinfundibularis, anterior and posterior pars ventralis hypothalami, pars dorsalis thalami, and the tegmentum motorium rhombencephali of larval, parasitic and upstream migrant adult lampreys. Many of the SRIF containing neurons within the hypothalamus were cerebrospinal fluid (CSF)-contacting cells. SRIF fibers were found throughout most of the brain predominating within the nucleus praeopticus, pars ventralis hypothalami, and the nucleus interpeduncularis. No SRIF immunoreactivity was found within the neurophyophysis. The possible functions of these peptides within the brain of the lamprey are discussed.
Article PDF
Similar content being viewed by others

Avoid common mistakes on your manuscript.
References
Bern HA, Nishioka RS, Nagahama Y (1975) The relationship between nerve fibers and adenohypophysial cell types in the cichlid teleost, Tilapia mossambica. In: Arvy L (ed) Recherches biologiques contemporaines. Imprimérie Vagner, Nancy, pp 179–194
Crim JW, Urano A, Gorbman A (1979a) Immunocytochemical studies of luteinizing hormone-releasing hormone in brains of agnathan fishes. I. Comparisons of adult Pacific lamprey (Entosphenus tridentata) and the Pacific hagfish (Eptatretus stouti). Gen Comp Endocrinol 17:294–305
Crim JW, Urano A, Gorbman A (1979b) Immunocytochemical studies of luteinizing hormone-releasing hormone in brains of agnathan fishes. II. Patterns of immunoreactivity in larval and maturing western brook lamprey (Lampetra richardsoni). Gen Comp Endocrinol 38:290–299
Falkmer S, El-Salhy M, Titlbach M (1984) Evolution of the neuroendocrine system in vertebrates. A review with particular reference to the phylogeny and postnatal maturation of the islet parenchyma. In: Falkmer S, Håkanson R, Sundler F (eds) Evolution and tumour pathology of the neuroendocrine system in birds. Elsevier Science Publishers, New York
Filby AB, Gross DS (1983) Distribution of immunoreactive somatostatin in the primate hypothalamus. Cell Tissue Res 233:69–80
Fryer JN, Nishioka RS, Bern HA (1979) Somatostatin inhibition of teleost growth hormone secretion. Gen Comp Endocrinol 39:244–246
Fuxe K, Hökfelt T, Eneroth P, Gustafsson J-A, Skett P (1977) Prolactin-like immunoreactivity: localization in nerve terminals of rat hypothalamus. Science 196:899–900
Goossens N, Dierickx K, Vandesande F (1977) Immunocytochemical demonstration of the hypothalamus-hypophysial vasotocinergic system of Lampetra fluviatilis. Cell Tissue Res 177:317–323
Gorbman A, Dickhoff W, Vigna SR, Clark NB, Ralph CL (1983) Comparative endocrinology. John Wiley and Sons, New York
Grau EG, Nishioka RS, Young G, Bern HA (1985) Somatostatinlike immunoreactivity in the pituitary and brain of three teleost fish species: somatostatin as a potential regulator of prolactin cell function. Gen Comp Endocrinol 59:350–357
Gurr E (1962) Staining animal tissue. Practical and theoretical. Leonard Hill Ltd., London
Hansen BL, Hansen GN (1982) Immunocytochemical demonstration of somatotropin-like and prolactin-like activity in the brain of Calamoichthys calabaricus (Actinopterygii). Cell Tissue Res 222:615–627
Heier P (1948) Fundamental principles in the structure of the brain. A study of the brain of Petromyzon fluviatilis. Act Anat [Suppl] VI:3–213
Krieger DT, Liotta AS (1979) Pituitary hormones in brain: Where, how and why? Science 205:366–372
Kronheim S, Berelowitz M, Pimstone BL (1977) The presence of immunoreactive growth hormone release-inhibiting hormone in normal cerebrospinal fluid. Clin Endocrinol 6:411–415
Larsen LO, Rothwell B (1972) Adenohypophysis. In: Hardisty MW, Potter IC (eds) The biology of lampreys. Vol I. Academic Press, New York, pp 1–67
Luft R, Efendri S, Hökfelt T (1978) Somatostatin-both hormone and neurotransmitter? Diabetologia 14:1–13
Nozaki M, Gorbman A (1983) Immunocytochemical localization of somatostatin and vasotocin in the brain of the Pacific hagfish Eptatretus stouti. Cell Tissue Res 229:541–550
Nozaki M, Fernholm B, Kobayashi H (1975) Ependymal absorption of peroxidase in the third ventricle of the hagfish Eptatretus burgeri (Girard). Acta Zool (Stockh) 56:265–269
Nozaki M, Tsukahara T, Kobayashi H (1984) An immunocytochemical study on the distribution of neuropeptides in the brain of certain species of fish. Biomed Res 4 [Suppl]: 135–145
Olivereau M, Ollevier F, Vandesande F, Olivereau J (1984) Somatostatin in the brain and the pituitary of some teleosts. Immunocytochemical identification and the effect of starvation. Cell Tissue Res 238:289–296
Pacold ST, Kirsteins L, Hojvat S, Lawrence AM, Hagen TC (1978) Biologically active pituitary hormones in the rat brain amygdaloid nucleus. Science 199:804–805
Palkovits M (1980) Topography of chemically identified neurons in the central nervous system: Progress in 1977–1979. Medical Biol 58:188–227
Patzner RA, Erhart G, Adam H (1982) Cell types in the adenohypophysis of the hagfish, Myxine glutinosa (Cyclostomata). Cell Tissue Res 223:583–592
Percy R, Leatherland JF, Beamish FWH (1975) Structure and ultrastructure of the pituitary gland in the sea lamprey, Petromyzon marinus at different stages in its life cycle. Cell Tissue Res 157:141–164
Rodríguez EM (1976) The cerebrospinal fluid as a pathway in neuroendocrine integration. J Endocrinol 71:407–443
Rodriguez EM, Perra P, Rodriguez S, Aguado LI (1982) Evidence for the participation of the CSF and periventricular structures in certain neuroendocrine mechanisms. Front Horm Res 9:142–158
Sétáló G, Flerkó B, Arimura A, Schally AV (1978) Brain cells as producers of releasing and inhibiting hormones. Int Rev Cytol [Suppl] 7:1–52
Sterba G (1972) Neuro- and gliasecretion. In: Hardisty MW, Potter IC (eds) The biology of lampreys. Vol. 2. Academic Press, New York, pp 69–89
Sternberger A (1979) Immunocytochemistry. John Wiley and Sons, New York
Tsuneki K, Gorbman A (1975a) Ultrastructure of the anterior neurohypophysis and the pars distalis of the lamprey, Lampetra tridentata. Gen Comp Endocrinol 25:487–508
Tsuneki K, Gorbman A (1975b) Ultrastructure of the pars nervosa and pars intermedia of the lamprey, Lampetra tridentata. Cell Tissues Res 157:165–184
Vigh-Teichmann I, Vigh B, Korf H-W, Oksche A (1983) CSF contacting and other somatostatin-immunoreactive neurons in the brains of Anguilla anguilla, Phoxinus phoxinus and Salmo gairdneri (Teleostei). Cell Tissue Res 233:319–334
Wright GM (1983) Ultrastructure of the adenohypophysis in the larval anadromous sea lamprey, Petromyzon marinus L. J Morphol 176:325–339
Wright GM (1984) Immunocytochemical study of growth hormone, prolactin, and thyroid-stimulating hormone in the adenohypophysis of the sea lamprey, Petromyzon marinus L., during its upstream migration. Gen Comp Endocrinol 55:269–274
Yui R (1983) Immunohistochemical studies on peptide neurons in the hypothalamus of the bullfrog Rana catesbiana. Gen Comp Endocrinol 49:195–209
Author information
Authors and Affiliations
Rights and permissions
About this article
Cite this article
Wright, G.M. Immunocytochemical demonstration of growth hormone, prolactin and somatostatin-like immunoreactivities in the brain of larval, young adult and upstream migrant adult sea lamprey, Petromyzon marinus . Cell Tissue Res. 246, 23–31 (1986). https://doi.org/10.1007/BF00218994
Accepted:
Issue Date:
DOI: https://doi.org/10.1007/BF00218994