
Abstract
By the year 2050, the global human population is predicted to increase by 2.5 billion reaching 9.6 billion people. To feed the world’s 9.6 billion people, the Food and Agriculture Organization estimates that global food production must increase by 70%. Moreover, the productivity of major food crops is affected by environment induced abiotic stressors that further expand the food demand-supply gap. Among the food crops cereals are most important in ensuring food security, yet they are also the most vulnerable to abiotic stresses. Due to various abiotic stressors, cereal productivity is decreasing; thus, mitigating these yield losses is critical for all nations to satisfy rising food demands. Besides abiotic stressors, ongoing climate change are also posing severe obstacles to obtaining the required agricultural production levels to meet the expanding food demands. Among the abiotic stresses drought, temperature and soil salinity are the most severe, resulting in massive crop yield losses. Therefore, tolerance to abiotic stresses has typically been a long-term goal for plant breeders. In this chapter, the consequences of abiotic stresses, mechanism of abiotic stress tolerance and the role of various breeding strategies in developing abiotic stress-tolerant cultivars have been discussed.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
8.1 Cereals: An Ideal Crop to Achieve Food Security
Cereal is derived from the Latin word ‘cerealis’, which means ‘grain’, and refers to a type of fruit called a caryopsis, which is made up of endosperm, germ and bran. Cereals, such as wheat, rice, maize, sorghum, millet, barley and rye, are annual grass members of the grass family (a monocot family Poaceae, commonly known as Gramineae), with long, thin stalks and starchy grains used as food. Cereals have evolved to thrive in settings where they are frequently subjected to various stressors, including high temperatures, drought, salt, mineral toxicity and water scarcity (Giordano et al. 2021; Kumari et al. 2021). They are widely used crops in global agriculture, with approximately 2979 million tonnes being harvested worldwide in 2019 (FAOSTAT 2021 http://www.fao.org/faostat/en/). Maize, wheat and rice are the three most significant cereal crops, accounting for at least 85% of global grain output. In 2019, 1148.49, 765.77 and 755.47 million tonnes of rice, wheat and maize, respectively, were harvested (Table 8.1). The cereal statistics for the area harvested and grain yield in the last 10 years showed significant growth and thus positively impacted the production of cereals (Fig. 8.1). Cereals provide essential nutrients including proteins, carbohydrates, minerals, amino acids, fibres and micronutrients, including vitamins, magnesium and zinc (O'Neil et al. 2010; Papanikolaou and Fulgoni 2017).

Comparative analysis on area harvested, yield and production of cereals globally (2009–2019)
Globally, 48% of the world’s cereal grains are produced in Asia, 26% in America and 18% in Europe (Fig. 8.2). Rice, sorghum, millet and wheat are commonly grown in Asia; corn and sorghum are grown in the United States, while barley, rye and oats are grown in Europe. Cereals are an essential source of nutrients in industrialized and developing countries, although their usage patterns differ. More than 70% of total grain output is used as feed for livestock in affluent nations, whereas 68 to 98% of cereal production is used for human consumption in developing countries (Olugbire et al. 2021).

Region-wise production share of cereals (2019)
Compared to dietary legumes and oilseeds, cereal grains have a low protein level, with rice having the lowest. In all cereal grains, lysine is the most limited of the necessary amino acids for humans. Cysteine, methionine and sulphur-containing amino acids are abundant in most grain proteins. Among the cereal crops, barley has a higher lysine content. The presence of antinutrients such as metal chelates, antivitamins, goitrogens, cyanogens, protease and amylase inhibitors, toxic phenolic glycosides and amino acid derivatives are known to influence the consumption of legumes (Mohan et al. 2016). As a result, adequate processing of the cereal-legume combination is necessary before ingestion to reduce these antinutrients. Cereal grain products have lower nutritional and sensory characteristics compared to animal meals. Physical, chemical, biological and physiological changes can improve the grain nutritional and visual characteristics (Piltz et al. 2021). Furthermore, natural processes such as fermentation and regulated germination with natural microflora help to improve the quality of cereal-based foods. The nutritional qualities of the most important cereal crops are furnished in Table 8.2.
8.2 Abiotic Stresses: Impact on Cereal Production
It is often difficult to breed a species for more than one robust feature at a time since individual plants react so differently to similar abiotic stress stimuli, but that is precisely what plant breeders are aiming for. Globally, abiotic and biotic stresses reduce average yield by more than 50% (Oerke et al. 1999; Raina and Khan 2020). However, abiotic stresses, especially salinity, drought, and temperature, are the major constraints for cereal production (Acquaah 2007; Martinez-Beltran and Manzur 2005; Munns 2002; Lobell and Field 2007). Abiotic stressors have a negative impact on several growth phases (Raina et al. 2020a). They are highly complicated, affecting crop dynamisms such as blooming, grain filling and maturity at the transcriptome, cellular and physiological levels (Atkinson and Urwin 2012; Maiti and Satya 2014; Paul and Roychoudhury 2019). The primary abiotic stressors impacting contemporary agricultural systems are atmosphere, soil, water and related variables (Sahu et al. 2014). Water is a major factor that induces abiotic stress in cereals, including water scarcity, salinity and waterlogging. Water scarcity, falling rainfall and rising temperature are the major limitations for agriculture, all of which substantially impact agricultural productivity.
8.2.1 Drought Stress
Undeniably, drought is one of the principal abiotic stresses in the world. Drought stress affects various morpho-physiological aspects of the plant from anthesis to maturity and significantly reduces productivity. The need of the hour is to develop stress-resistant genotypes which could thrive well under severe environment (Rabara et al. 2021). In most cases, rainwater produces a flooded condition in the field. Since water replaces almost all air in soil pore space, the oxygen content in flooded conditions decreases to zero within 24 h. Roots require oxygen to maintain vital cellular processes and cell viability. Waterlogging reduces the amount of oxygen available to the roots; if the roots consume any residual oxygen from flooded or waterlogged soils, the biological functions of the roots will get disrupted. As a result, the leaves and stems cannot acquire sufficient minerals and nutrients, and the roots begin to die due to waterlogging (Liliane and Charles 2020).
8.2.2 Temperature Stress
Crop species have been divided into three categories based on their temperature sensitivity: chilling-sensitive, freezing-sensitive and freezing-resistant plants (Kai and Iba 2014). Freezing may affect growth and produce frost-hardening/cold hardening, as well as causing the formation of reactive oxygen species, which could disrupt membrane components and induce protein denaturation (Beck et al. 2004; Baek and Skinner 2012). The crops subjected to a high temperature result in expansion-induced lysis, phase changes, lesions in membranes and physical damage (Tomás et al. 2020).
8.2.3 Salinity Stress
Salinity stress has various consequences in plants, including ionic and osmotic effects, nutritional and hormonal imbalances and the formation of reactive oxygen species (Rao et al. 2019). The buildup of sodium (Na+) and chloride (Cl−) ions has a significant impact on plant development and production, resulting in ionic, osmotic and oxidative stress (Yildiz et al. 2020). Multiple metabolic activities, such as protein translation, transcription and enzyme activity, are influenced by Na+, resulting in osmotic stress. There is a genetic basis for salt responsiveness, as evidenced by salt-tolerant and salt-sensitive crop species (Roychoudhury et al. 2008). Salt tolerance across varieties has been recognized since the 1930s (Epstein 1977, 1983), and intra-specific salt tolerance selection has been documented in rice (Akbar and Yabuno 1977) and barley (Epstein et al. 1980).
8.2.4 Heavy Metal Stress
Abiotic stress arises from soil-related factors including soil properties, pollution and degradation. The injudicious use of certain hazardous pesticide chemicals facilitates their entry in the natural environment in various ways based on their solubility. Abiotic stress can also be caused by a nutrient shortage or the presence of harmful chemicals in the soil, such as heavy metals (Sahu et al. 2014). Heavy metals including manganese, zinc, copper, magnesium, molybdenum, boron and nickel substantially impact plant morphological, metabolic and physiological abnormalities (Roychoudhury et al. 2012). It includes shoot chlorosis, lipid peroxidation and protein breakdown (Emamverdian et al. 2015). Nutrient insufficiency has long been thought to be the root cause of low agricultural yields. Only 3.03 billion hectares (22%) of the world’s 13.5 billion hectares is cultivable, while over two billion hectares is not suitable for cultivation. Oil shale disposal, soil heavy metal pollution and crude oil leakage negatively impact the root systems (Shah and Wu 2019).
In addition to the above-mentioned stresses, plant tissues are injured when the weather is hot, humid and foggy with a slight breeze. Plant reproductive development, chlorosis and necrosis are all affected by chilling stress, which includes decreased leaf growth and wilting. Ultraviolet and ionizing radiations have a variety of effects on the growth and development of cereal crops. Radiation affects stomatal function, cell survival, seed development and fertility (Foroughbakhch Pournavab et al. 2019; Metwally et al. 2019). Photon irradiation causes cellular damage in root and leaf tissues of cereals. Fast-flowing winds also reduce the phytohormonal content of cereal crop roots and shoots. The wind direction and velocity have an impact on plant growth and development (Sahu et al. 2014). Rainfall is one of the major abiotic stress variables impacting soil erosion and crop productivity in rain-fed agriculture in semi-arid areas. It regulates the acidity and salinity of the soil. Acid rain occurs when sulphur dioxide (SO2) and nitrogen oxides (NOx) combine with water and oxygen in the atmosphere (Gong et al. 2019). Acid rain impairs vital processes of growth and development in cereal crops.
8.3 Origin of Abiotic Stress Tolerance
The origin of abiotic stress tolerance in agricultural plants can be found in a variety of places. Landraces, wild relatives, high-yielding varieties, initial breeding materials and advanced breeding materials may all harbour tolerance. Landraces from arid regions have been effectively employed in breeding to create open-pollinated types or hybrids for water-scarce situations. Abiotic stress providers include wild species and progenitors of our farmed crops (Table 8.3). The likelihood of identifying the desired genes and even the projected challenges and expected success in introgression of these genes into the chosen recurrent cultivar all influence the genetic resource used as a source for abiotic stress resistance. A substantial genetic diversity occurs in the breeding materials and even in some improved cultivars of different crop species for drought and salinity resistance (Basu and Roychoudhury 2021). Because this is the least troublesome of all sources of drought and salinity resistance, an initial aim of the breeder should be to find and use such sources. Drought and salt resistance traits are often found in landraces (old or desi varieties) that have evolved and are adapted to drought and salinity conditions. Efforts in utilizing wild relatives should be concentrated only when the diversity in top breeding materials and landraces has been exhausted.
8.4 Response of Cereals Towards Abiotic Stress
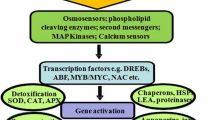
Cereals have a diverse set of strategies at the genetic, physiological, biochemical and molecular levels. Nevertheless, new progress in traditional, marker-assisted breeding and genetic engineering has made it possible to develop drought-tolerant crops (Oladosu et al. 2019; Rosero et al. 2020). Many crops are sensitive to high salinity and could not withstand saline conditions; however, certain crops are adapted to thrive in harsher salt environment prevalent in coastal locations like salt marshes. The high rate of evaporation within those areas concentrates salts in the mineral composition of the soil. These crops have evolved morpho-physiological and reproductive adaptations to salty, waterlogged and anaerobic environments. Such crops can withstand salt stress primarily through three main mechanisms: osmotic tolerance, ion exclusion and tissue tolerance. Long-distance signalling waves control osmotic tolerance by reducing cell growth in root tips, leaves and regulate stomatal conductance (Rajendran et al. 2009; Roy et al. 2014). Ion exclusion is primarily concerned with the transfer of sodium (Na+) and chlorine (Cl−) into roots, which prevents the buildup of Na+ in shoots. Tissue tolerance entails exposing tissues to accumulating Na+ and Cl− at the cellular and subcellular levels, the build up of suitable solutes and the enzyme that catalyzes the detoxification of reactive oxygen species (Reddy et al. 2017). The capacity of cereal crops to withstand dominating abiotic stress, which includes water deficiency (drought), flood (anoxia), salinity, high/low temperature and other osmotic stressors, is an essential component of yield resilience and has long been a goal for plant breeders (Halford et al. 2014). Due to the ever-increasing population, the fast-changing global environment impacts agricultural production and food supply, resulting in a need for stress-tolerant crop (Takeda and Matsuoka 2008; Newton et al. 2011). Understanding how plant responses to diverse stressors, interact at the molecular level is critical for creating stress-tolerant crops (Paul and Roychoudhury 2019). The adaptive mechanism of abiotic stress in plants is associated with various traits are shown in Fig. 8.3.

Abiotic stress adaptive mechanism and their associated traits
8.5 Different Breeding Strategies for Improving Abiotic Stress Tolerance
Genetic modification for effective stress tolerance in plants is difficult due to the complex characteristics of abiotic stress events (Wang et al. 2007). Breeding for abiotic stress is an important strategy to fight yield loss. Even in ancient times, the necessity for stress-tolerant crops was obvious (Jacobsen and Adams 1958). However, efforts to increase crop performance under environmental stressors have mostly failed due to a lack of understanding of the underlying processes of stress tolerance in plants. Information on the genetic basis of abiotic stress tolerance, method of inheritance, size of gene effects, heterosis, combining ability and their mechanism of action are required to plan effective breeding programmes for generating abiotic stress-tolerant cultivars. Breeding methodology for crops should be adopted based on the type of reproduction, i.e. whether the species is self- or cross-pollinating. Pedigree and bulk methods may be employed for self-pollinated crops, whereas recurrent selection could be used for cross-pollinated crops. However, if the goal is to transfer a few stress-resistant characteristics to a high-yielding genotype, backcrossing is an appropriate strategy. Pedigree, modified bulk pedigree and anther culture approaches were used to create salinity-tolerant varieties. On the other hand, biparental mating (half-sib and full-sib) preserves a wide genetic basis while also allowing for the evolution of drought-resistant genotype (Yunus and Paroda 1982).
Nowadays, breeding strategies for improving abiotic stress tolerance include both conventional and non-conventional approaches. Various factors influence selecting an appropriate breeding programme developing stress-tolerant cultivars, including screening methodologies, sources and mechanisms of tolerance, gene action and heredity and their link to agronomic characteristics.
8.5.1 Conventional Approaches
Conventional approaches in plant breeding include selection and introduction, pedigree method, modified bulk pedigree method, shuttle breeding, mutation breeding, diallel selective mating system supplemented by marker-assisted selection (MAS), backcross method and recurrent selection. The timing, duration and severity of the stress are three key aspects for successful stress-resistance breeding. Using traditional breeding procedures, abiotic stress-tolerant lines of crops such as rice, wheat, maize and barley have been developed. However, identifying traits that correlate well with drought tolerance is difficult using this technique. Traditional efforts to develop crop plants resistant to abiotic stress have had mixed results (Richards 1996). This is attributed to various variables, including complexity caused by genotype due to environment (G × E) interactions. Stress-tolerant crop cultivars can be created through various methods, including introduction, selection, hybridization and mutation breeding. However, mutation breeding is considered a coherent and widely accepted tool to enhance abiotic stress tolerance in cereals.
8.5.1.1 Mutation Breeding
Nonetheless, conventional breeding approaches are laborious and time-consuming and do not yield the desired variation (Cassells and Doyel 2003). Therefore, agronomists are putting efforts into searching alternate ways to get the desired variation in crops within a short time. In this regard, mutation breeding is a coherent tool to generate mutants with desirable traits and enhanced genetic variability. Mutations are sudden heritable changes in an organism genome that play an essential role in increasing genetic variability (Hugo de Vries 1901). Mutations occur both spontaneously and are induced using different physical, chemical and combined mutagens. However, the frequency of spontaneous mutations is low and is not enough to meet the needs of crop improvement programmes. Therefore, mutations are induced artificially to enhance the frequency of mutations. In the beginning, Muller (1927), employed X-rays to irradiate Drosophila flies to enhance genetic variability. The discovery of the mutagenic potential of X-rays in maize and barley was the pioneering event in the establishment of mutation breeding for crop improvement programmes (Stadler 1928). The induction of mutations within an organism genome has been used in plant breeding since the discovery of the mutagenic effects of X-rays on Drosophila flies (Muller 1927). These discoveries on the induction of artificial mutations encouraged plant breeders to use different physical and chemical mutagens for crop improvement programmes. As a result, thousands of mutant varieties with improved yield, quality, stress tolerance and adaptability were developed in various crop species. Mutation breeding is now an established breeding strategy to achieve crop varieties with improved agronomic traits, including abiotic stress tolerance in cereals.
8.5.1.2 Role of Mutation Breeding in Improving Abiotic Stress Tolerance in Cereals
Nowadays, mutation breeding for crop improvement is based on physical and chemical mutagens and variations in in vitro culture, called ‘somaclonal variation’. The use of induced mutation for crop improvement is reflected by the fact that 3364, including 1596 improved mutant varieties of cereals, have been developed and officially released (Raina et al. 2016). These mutant varieties are cultivated on millions of hectares of cultivated land that generate billions of dollars (Laskar et al. 2018a, b; Goyal et al. 2019a, b). This has led to a tremendous economic impact on agriculture and its allied sectors worldwide (Das et al. 2014; Khursheed et al. 2018a, b, c). Ever since the historical discoveries of Muller and Stadler, different mutagens were employed to enhance genetic variability. However, radiations were preferred to achieve the improved genetic variability in food crops (Hassan et al. 2018; Laskar et al. 2019). In the last five decades, various countries such as China, India, Pakistan, Bangladesh, Vietnam, Thailand, Italy, Sweden, the United States of America, Canada and Japan took up extensive crop improvement programmes through induced mutagenesis. This has made remarkable achievements in developing elite mutant varieties in a wide range of food crops, particularly cereals and pulses (Raina et al. 2019; Wani et al. 2021a, b).
Mutation breeding has been successful in developing mutant varieties with improved tolerance to abiotic stresses (Laskar et al. 2015; Khursheed et al. 2019; Goyal et al. 2021). The improved mutant varieties play a vital role in mitigating the chronic hunger and malnutrition issues and achieving global food and nutrition security (Khursheed et al. 2016; Goyal et al. 2020a, b). Mutation breeding offers an opportunity to improve abiotic stress tolerance without altering the genetic constitution (Raina et al. 2017; Tantray et al. 2017). Unlike genetically engineered crops, mutant varieties do not possess any alien genes. Hence, there are no detrimental issues of induced mutations associated with human health, religious and social ethics. In the last two decades, enormous advances have been achieved in this field, and thousands of mutants have been released as new cultivars (Wani et al. 2017; Ansari et al. 2021). In rice, the results have been remarkable. The FAO/IAEA database displays about 853 mutant varieties of rice, 311 mutant varieties of barley, 298 varieties of wheat and 96 mutant varieties of maize developed by the use of induced mutations (https://mvd.iaea.org/ accessed on 10-07-2021). Till now, 160 mutant varieties of cereals have been developed that reflect improved tolerance to a wide range of abiotic stresses such as drought, heat, salinity and cold (Baloch et al. 2002; Saleem et al. 2005; Cassells and Doyel 2003; Parry et al. 2009) (Table 8.4). For instance, rice seeds irradiated with carbon or neon ions have generated a high salt tolerance mutant variety (Hayashi et al. 2007). The gamma radiation mutant rice varieties, Zhefu 802 and Basmati 370, revealed improved cold tolerance (Ahloowalia et al. 2003). Likewise Bastion, Secret and Taran are cold-tolerant barley mutant varieties (Shevtsov et al. 2003). In Finland, a barley mutant, Balder J, had a higher yield and improved drought resistance (Kharkwal and Shu 2009). Luther and Pennrad are high-yielding and lodging-resistant barley mutant varieties (Kharkwal and Shu 2009).
Many such varieties of abiotic stress-tolerant mutant crops have been released in different countries all over the world (Raina and Danish 2018; Raina et al. 2020b). Therefore, induced mutagenesis for resistance to abiotic stresses is a possible breeding approach that creates new desirable genetic variability of agronomic importance (Amin et al. 2016, 2019; Raina et al. 2018a, b). Thus, mutation-assisted plant breeding has a crucial role in developing ‘designer crop varieties’ to address the qualms and challenges of global climate variability and plant-product insecurity (Raina et al. 2021a, b, c). Both efficiency and efficacy of mutation techniques in crop breeding can significantly be increased through molecular mutation breeding. High-throughput DNA technologies such as targeting induced limited lesions in genomes, high-resolution melt analysis and Ecotype targeting induced local lesions in genomes are the key techniques and resources in molecular mutation breeding (Das et al. 2014). Unlike biotic stresses, herbicides, pesticides and fertilizers cannot mitigate the harmful effects of abiotic stresses.
8.5.2 Non-Conventional Approaches
The traditional breeding method has been sluggish in creating high-yielding, stress-tolerant cultivars, owing to challenges in accurately defining the target environment, complicated interactions of stress tolerance with surroundings and a lack of adequate screening methods (Cooper et al. 1999; Wade et al. 1999). The necessity for numerous backcrosses to remove undesired characteristics, restriction to loci that provide a readily apparent phenotype and inadequacy if the gene pool lacks sufficient diversity in the trait of interest are all key drawbacks of traditional breeding. As a result, the current focus is on marker-aided breeding, which allows for the ‘pyramiding’ of desired characteristics for faster crop development with minimal resource input. Non-conventional approaches and discoveries such as the somaclonal approach, F1 anther culture, marker assisted selection and genetically modified crops are the most recent advances in developing abiotic stress tolerance in cereals. The most immediate and future solutions for increasing abiotic stress tolerance in cereal crops might be the recent ground breaking developments in bioinformatics and integrating omics technology. Identifying the genetic basis of stress tolerance and using the required salt stress tolerance-associated genes or QTL (quantitative trait loci) to produce varieties with increased salinity tolerance are prerequisites for improving salt tolerance. Omics techniques, such as genomics, functional genomics, genetic engineering, gene expression, protein or metabolite profile(s) and their overall phenotypic impacts, contribute to a better knowledge of stress tolerance mechanisms at the molecular level. The discovery and characterization of genes and particular genomic areas linked with quantitative and qualitative agronomic characteristics that have been critical in crop breeding approaches. A high-throughput marker-assisted approach has been widely employed in recent breeding projects to improve selection effectiveness and precision. The exploitation of natural genetic variants, either through direct selection in stressful situations or by the identification of QTLs and subsequent marker-assisted selection, and creating transgenic plants to introduce new genes or change the expression levels of existing genes to influence salt stress resistance are two primary techniques now being used to increase stress resistance. Simple genetic models were used to assess the genetic basis of stress tolerance in plants in the beginning. With the advent of molecular markers, inheritance of salt tolerance became more manageable since particular QTLs could be discovered. It is now feasible to establish the genetic basis of a trait and map particular chromosomal segments or QTL and estimate the relative contribution of each QTL to the variation of a trait. Genomic maps have been created in several crops to exploit genetic diversity, tag qualitative and quantitative characteristics (Butruille et al. 1999) and assess the stability of identified QTL across diverse environmental conditions (Hittalmani et al. 2002). Stable and consistent QTLs offer a great way to increase selection efficiency, especially for characteristics that are regulated by several genes and heavily impacted by the environment, such as salinity (Dudley 1993). The effectiveness of marker assisted selection is influenced by several parameters, including the distance between observed QTL and marker loci (Dudley 1993) and the fraction of total additive variation explained by the QTL (Lande and Thompson 1990). New genomic technologies are promising to advance breeding resistance to these stressors due to a better understanding of underlying mechanisms and identifying the implicated genes. Modern biotechnological tools such as genetic engineering have successfully developed transgenic plants resistant to various abiotic stresses (Jewell et al. 2010). However, stress-resistant transgenic plants did not receive public acceptance due to risks concerning human health, social and religious issues and environmental safety (Carpenter 2010; James 2011; Kathage and Qaim 2012; Seralini et al. 2012). Hence, conventional breeding methods seem more appropriate to develop stress-tolerant and environment-friendly crop varieties.
8.6 Conclusions and Future Directions
The majority of crop losses are caused by abiotic stressors, which account for more than half of all harvest losses. According to several research findings, salt and drought stress have a detrimental effect on plant growth, development, physiology and production. During the last century, conventional plant breeding significantly enhanced crop quality and yield and improved abiotic stress resistance, such as drought and salinity tolerance. However, establishing abiotic stress tolerance varieties/hybrids will take longer time. Crop varieties/lines/hybrids with higher tolerance to drought, salinity, high temperature and nutrient deficiency, developed through conventional and molecular breeding methods and genetic engineering, are important for meeting global food demands. Traditional breeding knowledge combined with marker-assisted selection makes it quicker and more effective to generate drought tolerance in crop plants using genotypic data to improve and sustain productivity in drought-prone environmental settings. There is a pressing need to develop strategies to boost food output, particularly in the stressed zones of the world. A breeder must identify the genetic basis of stress tolerance in crop plants to generate improved genotypes using either traditional breeding or biotechnological methods. Scientists from all around the globe are working hard to develop varieties with enhanced heterosis in stress-prone settings. The most promising, less resource-intensive, commercially feasible and socially acceptable strategy is to develop crop varieties with built-in salt, drought and heat tolerance.
References
Acquaah G (2007) Principles of plant genetics and breeding. Blackwell, Oxford
Ahloowalia BS, Maluszynski M, Nichterlein K (2003) Global impact of mutation-derived varieties. Euphytica 135:187–204
Akbar M, Yabuno T (1977) Breeding saline-resistant varieties of rice. IV. Inheritance of delayed type panicle sterility induced by salinity. Jap J Breed 27:237–240
Amin R, Laskar RA, Khursheed S, Raina A, Khan S (2016) Genetic sensitivity towards MMS mutagenesis assessed through in vitro growth and cytological test in Nigella sativa L. Life Sci Int Res J 3:2347–8691
Amin R, Wani MR, Raina A, Khursheed S, Khan S (2019) Induced morphological and chromosomal diversity in the mutagenized population of black cumin (Nigella sativa L.) using single and combination treatments of gamma rays and ethyl methane sulfonate. Jordan J Biol Sci 12(1):23–33
Ansari SB, Raina A, Amin R, Jahan R, Malik S, Khan S (2021) Mutation breeding for quality improvement: a case study for oilseed crops. In: Bhat TA (ed) Mutagenesis, cytotoxicity and crop improvement: revolutionizing food science. Cambridge Scholars, Newcastle upon Tyne, pp 171–221
Atkinson NJ, Urwin PE (2012) The interaction of plant biotic and abiotic stresses: from genes to the field. J Exp Bot 63:3523–3543
Baek KH, Skinner DZ (2012) Production of reactive oxygen species by freezing stress and the protective roles of antioxidant enzymes in plants. J Agric Chem Environ 1:34–40
Baloch AW, Soomro AM, Javed MA, Ahmed M, Bughio HR, Bughio MS, Mastoi NN (2002) Optimum plant density for high yield in rice (Oryza sativa L.). Asian J Plant Sci 2:273–276
Basu S, Roychoudhury A (2021) Transcript profiling of stress-responsive genes and metabolic changes during salinity in indica and japonica rice exhibit distinct varietal difference. Physiol Plant 173(4):1434–1447. https://doi.org/10.1111/ppl.13440
Beck EH, Heim R, Hansen J (2004) Plant resistance to cold stress: mechanisms and environmental signals triggering frost hardening and dehardening. J Biosci 29:449–459
Butruille DV, Guries RP, Osborn TC (1999) Linkage analysis of molecular markers and quantitative trait loci in populations of inbred backcross lines of Brassica napus L. Genetics 153:949–964
Carpenter J (2010) Peer-reviewed surveys indicate positive impact of commercialized GM crops. Nat Biotechnol 28:319–321
Cassells AC, Doyel BM (2003) Genetic engineering and mutation breeding for tolerance to abiotic and biotic stresses: science, technology and safety. Bulg J Plant Physiol 2003:52–82
Cooper M, Fukai S, Wade LJ (1999) How can breeding contribute to more productive and sustainable rainfed lowland rice systems? Field Crop Res 64:199–209
Das P, Mishra M, Lakra N, Singla-Pareek SL, Pareek A (2014) Mutation breeding: a powerful approach for obtaining abiotic stress tolerant crops and upgrading food security for human nutrition. In: Tomlekova N, Kozgar MI, Wani MR (eds) Mutagenesis: exploring novel genes and pathways. Wageningen Academic Publishers, Wageningen, pp 17–35
Dudley JW (1993) Molecular markers in plant improvement: manipulation of genes affecting quantitative traits. Crop Sci 33:660–668
Emamverdian A, Ding Y, Mokhberdoran F, Xie Y (2015) Heavy metal stress and some mechanisms of plant defense response. Sci World J 2015:75612
Epstein E (1977) Genetic potentials for solving problems of soil mineral stress: adaptation of crops to salinity. In: Wright MJ (ed) Plant adaptation to mineral stress in problem soils. Cornell University Agricultural Experiment Station, Ithaca, NY, pp 73–123
Epstein E (1983) Crops tolerant to salinity and other mineral stresses. In: Nugent J, O'Connor M (eds) Better crops for food, Ciba foundation symposium. Pitman, London, pp 61–82
Epstein E, Norlyn JD, Rush DW, Kingsbury R, Kelley DB, Wrana AF (1980) Saline culture of crops: a genetic approach. Science 210:399–404
Foroughbakhch Pournavab R, Bacópulos Mejía E, Benavides Mendoza A, Salas Cruz LR, Ngangyo Heya M (2019) Ultraviolet radiation effect on seed germination and seedling growth of common species from northeastern Mexico. Agronomy 9:269
Giordano M, Petropoulos S, Rouphael Y (2021) Response and defence mechanisms of vegetable crops against drought. Heat Salinity Stress Agric 11:463
Gong Y, Hao Y, Li J, Li H, Shen Z, Wang W, Wang S (2019) The effects of rainfall runoff pollutants on plant physiology in a bioretention system based on pilot experiments. Sustainability 11:6402
Goyal S, Wani MR, Laskar RA, Raina A, Khan S (2019a) Assessment on cytotoxic and mutagenic potency of gamma rays and EMS in Vigna mungo L Hepper. Biotecnol Vegetal 19:193–204
Goyal S, Wani MR, Laskar RA, Raina A, Amin R, Khan S (2019b) Induction of morphological mutations and mutant phenotyping in black gram (Vigna mungo (L.) Hepper) using gamma rays and EMS. Vegetos 32(4):464–472
Goyal S, Wani MR, Laskar RA, Raina A, Khan S (2020a) Mutagenic effectiveness and efficiency of individual and combination treatments of gamma rays and ethylmethanesulfonate in black gram (Vigna mungo (L.) Hepper). Adv Zool Bot 8(3):163–168
Goyal S, Wani MR, Laskar RA, Raina A, Khan S (2020b) Performance evaluation of induced mutant lines of black gram (Vigna mungo (L.) Hepper). Acta Fytotechn Zootechn 23(2):70–77
Goyal S, Wani MR, Raina A, Laskar RA, Khan S (2021) Phenotypic diversity in mutagenized population of urdbean (Vigna mungo (L.) Hepper). Heliyon 7(5):e06356
Halford NG, Curtis TY, Chen Z, Huang J (2014) Effects of abiotic stress and crop management on cereal grain composition: implications for food quality and safety. J Exp Bot 66:1145–1156
Hassan N, Laskar RA, Raina A, Khan S (2018) Maleic hydrazide induced variability in fenugreek (Trigonella foenum-graecum L.) cultivars CO1 and Rmt-1. Res Rev J Bot Sci 7(1):19–28
Hayashi Y, Takehisa H, Kazama Y, Ichida H, Ryuto H, Fukunishi N, Abe T (2007) Effects of ion beam irradiation on mutation induction in rice. In: Proceedings of cyclotrons and their applications, Eighteenth International Conference, pp 237–239
Hittalmani S, Shahidhar HE, Bagali PG, Huang N, Sidhu JS, Singh VP, Khush GS (2002) Molecular mapping of quantitative trait loci for plant growth, yield and yield related traits across diverse locations in a doubled haploid rice population. Euphytica 125:207–214
Hugo de Vries (1901) Die mutation theorie. Viet and Co., Leipzig. 1 Von, 1-648
Jacobsen T, Adams RM (1958) Salt and silt in ancient Mesopotamian agriculture. Science 128:1251–1258
James C (2011) Executive summary of global status of commercialized biotech/GM crops. ISAAA briefs no. 34. ISAAA, Ithaca, NY
Jewell MC, Campbell BC, Godwin ID (2010) Transgenic plant for abiotic stress resistance. In: Kole C, Michler CH, Abbott AG, Hall TC (eds) Transgenic crop plants. Springer Verlag, Heidelberg, pp 67–132
Kai H, Iba K (2014) Temperature stress in plants. In: eLS. Wiley, Chichester
Kathage J, Qaim M (2012) Economic impacts and impact dynamics of Bt (bacillus thuringiensis) cotton in India. Proc Natl Acad Sci U S A 109:11652–11656
Kharkwal MC, Shu QY (2009) The role of induced mutations in world food security. In: Shu QY (ed) Induced plant mutations in the genomics era. Food and Agriculture Organization of the United Nations, Rome, pp 33–38
Khursheed S, Raina A, Khan S (2016) Improvement of yield and mineral content in two cultivars of Vicia faba L. through physical and chemical mutagenesis and their character association analysis. Arch Curr Res Int 4(1):1–7
Khursheed S, Raina A, Amin R, Wani MR, Khan S (2018a) Quantitative analysis of genetic parameters in the mutagenized population of faba bean (Vicia faba L.). Res Crops 19(2):276–284
Khursheed S, Raina A, Laskar RA, Khan S (2018b) Effect of gamma radiation and EMS on mutation rate: their effectiveness and efficiency in faba bean (Vicia faba L.). Caryologia 71(4):397–404
Khursheed S, Raina A, Khan S (2018c) Physiological response of two cultivars of faba bean using physical and chemical mutagenesis. Int J Adv Res Sci Eng 7(4):897–905
Khursheed S, Raina A, Parveen K, Khan S (2019) Induced phenotypic diversity in the mutagenized populations of faba bean using physical and chemical mutagenesis. J Saudi Soc Agric Sci 18(2):113–119. https://doi.org/10.1016/j.jssas.2017.03.001
Kumari VV, Roy A, Vijayan R, Banerjee P, Verma VC, Nalia A, Pramanik M, Mukherjee B, Ghosh A, Reja H et al (2021) Drought and heat stress in cool-season food legumes in sub-tropical regions: consequences, adaptation, and mitigation strategies. Plan Theory 10:1038
Lande R, Thompson R (1990) Efficiency of marker-assisted selection in the improvement of quantitative traits. Genetics 124:743–756
Laskar RA, Khan S, Khursheed S, Raina A, Amin R (2015) Quantitative analysis of induced phenotypic diversity in chickpea using physical and chemical mutagenesis. J Agron 14:3–102
Laskar RA, Laskar AA, Raina A, Amin R (2018a) Induced mutation analysis with biochemical and molecular characterization of high yielding lentil mutant lines. Int J Biol Macromol 109:167–179
Laskar RA, Wani MR, Raina A, Amin R, Khan S (2018b) Morphological characterization of gamma rays induced multipodding mutant (mp) in lentil cultivar pant L 406. Int J Radiat Biol 94(11):1049–1053
Laskar RA, Khan S, Deb CR, Tomlekova N, Wani MR, Raina A, Amin R (2019) Lentil (Lens culinaris Medik.) diversity, cytogenetics and breeding. In: Al-Khayri JM et al (eds) Advances in plant breeding: legumes. Springer, Cham. https://doi.org/10.1007/978-3-030-23400-3_9
Liliane TN, Charles MS (2020) Factors affecting yield of crops. In: Agronomy-climate change & food security. London, IntechOpen, p 9
Lobell DB, Field CB (2007) Global scale climate-crop yield relationships and the impacts of recent warming. Environ Res Lett 2:14002–14008
Maiti R, Satya P (2014) Research advances in major cereal crops for adaptation to abiotic stresses. GM Crop Food 5:259–279
Martinez-Beltran J, Manzur CL (2005) Overview of salinity problems in the world and FAO strategies to address the problem. In: Proceedings of International Salinity Forum, Riverside, CA, USA, pp 311–313
Metwally SA, Shoaib RM, Hashish KI, El-Tayeb TA (2019) In vitro ultraviolet radiation effects on growth, chemical constituents and molecular aspects of Spathiphyllum plant. Bull Natl Res Cent 43:94
Mohan V, Tresina P, Daffodil E (2016) Antinutritional factors in legume seeds: characteristics and determination. In: Encyclopedia of food and health, pp 211–220
Muller HJ (1927) Artificial transmutation of the gene. Science 66:84–87
Munns R (2002) Comparative physiology of salt and water stress. Plant Cell Environ 25:239–250
Newton AC, Johnson SN, Gregory PJ (2011) Implications of climate change for diseases, crop yields and food security. Euphytica 179:3–18
Oerke ECC, Weber A, Dehne WH, Schonbeck F (1999) Crop production and crop protection. Elsevier Science, Amsterdam
Oladosu Y, Rafii MY, Samuel C, Fatai A, Magaji U, Kareem I, Kamarudin ZS, Muhammad II, Kolapo K (2019) Drought resistance in rice from conventional to molecular breeding: a review. Int J Mol Sci 20:3519
Olugbire OO, Olorunfemi S, Oke DO (2021) Global utilization of cereals: sustainability and environmental issues. AgroScience 20:9–14
O'Neil CE, Nicklas TA, Zanovec M, Cho S (2010) Whole-grain consumption is associated with diet quality and nutrient intake in adults: the National Health and nutrition examination survey, 1999–2004. J Am Diet Assoc 110:1461–1468
Papanikolaou Y, Fulgoni VL (2017) Certain grain foods can be meaningful contributors to nutrient density in the diets of US children and adolescents: data from the National Health and nutrition examination survey, 2009–2012. Nutrients 9:160
Parry MAJ, Madgwick PJ, Bayon C, Tearall K, Lopez AH, Baudo M, Rakszegi M, Hamada W, Al-Yassin A, Ouabbou H, Labhilili M, Phillips AL (2009) Mutation discovery for crop improvement. J Exp Bot 60:2817–2825
Paul S, Roychoudhury A (2019) Comparative analysis of the expression of candidate genes governing salt tolerance and yield attributes in two contrasting Rice genotypes, encountering salt stress during grain development. J Plant Growth Regul 38:539–556
Piltz JW, Rodham CA, Wilkins JF, Hackney BF (2021) A comparison of cereal and cereal/vetch crops for fodder conservation. Agriculture 11:459
Rabara R, Msanne J, Basu S, Ferrer M, Roychoudhury A (2021) Coping with inclement weather conditions due to high temperature and water deficit in rice: an insight from genetic and biochemical perspectives. Physiol Plant 172(2):487–504
Raina A, Danish M (2018) Mutagenesis in plant breeding for disease and pathogen resistance. Agric Res Technol 13(1):1–2
Raina A, Khan S (2020) Increasing rice grain yield under biotic stresses: mutagenesis, transgenics and genomics approaches. In: Aryadeep C (ed) Rice research for quality improvement: genomics and genetic engineering. Springer, Berlin, pp 149–178. https://doi.org/10.1007/978-981-15-5337-0_8
Raina A, Laskar RA, Khursheed S, Amin R, Parveen K, Khan S (2016) Role of mutation breeding in crop improvement-past, present and future. Asian Res J Agr 2:1–13
Raina A, Laskar RA, Khursheed S, Khan S, Parveen K, Amin R (2017) Induced physical and chemical mutagenesis for improvement of yield attributing traits and their correlation analysis in chickpea. Int Lett Nat Sci 61:14–22
Raina A, Laskar RA, Jahan R, Khursheed S, Amin R, Wani MR, Nisa TN, Khan S (2018a) Mutation breeding for crop improvement. In: Ansari MW, Kumar S, Babeeta CK, Wattal RK (eds) Introduction to challenges and strategies to improve crop productivity in changing environment. Enriched Public Pvt Ltd, New Delhi, pp 303–317
Raina A, Khursheed S, Khan S (2018b) Optimisation of mutagen doses for gamma rays and sodium azide in cowpea genotypes. Trends Biosci 11(13):2386–2389
Raina A, Khan S, Laskar RA, Wani MR, Mushtaq W (2019) Chickpea (Cicer arietinum L.) cytogenetics, genetic diversity and breeding. In: Al-Khayri JM et al (eds) Advances in plant breeding: legumes. Springer, Cham. https://doi.org/10.1007/978-3-030-23400-3_3
Raina A, Khan S, Sahu P, Roa R (2020a) Increasing rice grain yield under abiotic stresses: mutagenesis, transgenics and genomics approaches. In: Aryadeep C (ed) Rice research for quality improvement: genomics and genetic engineering. Springer, Berlin, pp 753–777. https://doi.org/10.1007/978-981-15-4120-9_31
Raina A, Laskar RA, Tantray YR, Khursheed S, Wani MR, Khan S (2020b) Characterization of induced high yielding cowpea mutant lines using physiological, biochemical and molecular markers. Sci Rep 10(1):1–22
Raina A, Sahu D, Parmeshwar K, Laskar RA, Rajora N, Soa R, Khan S, Ganai RA (2021a) Mechanisms of genome maintenance in plants: playing it safe with breaks and bumps. Front Genet 12:675686. https://doi.org/10.3389/fgene.2021.675686
Raina A, Ansari SB, Khursheed S, Wani MR, Khan S, Bhat TA (2021b) Mutagens their types and mechanism of action with an emphasis on sodium azide and gamma radiations. In: Bhat TA (ed) Mutagenesis, cytotoxicity and crop improvement: revolutionizing food science. Cambridge scholars, Newcastle upon Tyne, pp 1–37
Raina A, Laskar RA, Malik S, Wani MR, Khan S, Bhat TA (2021c) Plant mutagenesis: principle and application in crop improvement. In: Bhat TA (ed) Mutagenesis, cytotoxicity and crop improvement: revolutionizing food science. Cambridge Scholars, Newcastle upon Tyne, pp 38–65
Rajendran K, Tester M, Roy SJ (2009) Quantifying the three main components of salinity tolerance in cereals. Plant Cell Environ 32:237–249
Rao MPN, Dong ZY, Xiao M, Li WJ (2019) Effect of salt stress on plants and role of microbes in promoting plant growth under salt stress. In: Microorganisms in saline environments: strategies and functions. Springer, Cham, pp 423–435
Reddy INBL, Kim BK, Yoon IS, Kim KH, Kwon TR (2017) Salt tolerance in rice: focus on mechanisms and approaches. Ric Sci 24:123–144
Richards RA (1996) Defining selection criteria to improve yield under drought. Plant Growth Regul 20:57–166
Rosero A, Berdugo-Cely JA, Šamajová O, Šamaj J, Cerkal R (2020) A dual strategy of breeding for drought tolerance and introducing drought-tolerant, underutilized crops into production systems to enhance their resilience to water deficiency. Plan Theory 9:1263
Roy SJ, Negrão S, Tester M (2014) Salt resistant crop plants. Curr Opin Biotechnol 26:115–124
Roychoudhury A, Basu S, Sarkar SN, Sengupta DN (2008) Comparative physiological and molecular responses of a common aromatic indica rice cultivar to high salinity with non-aromatic indica rice cultivars. Plant Cell Rep 27(8):1395–1410
Roychoudhury A, Basu S, Sengupta DN (2012) Antioxidants and stress-related metabolites in the seedlings of two indica rice varieties exposed to cadmium chloride toxicity. Acta Physiol Plant 34(3):835–847
Sahu M, Dehury B, Modi MK, Barooah M (2014) Functional genomics and bioinformatics approach to understand regulation of abiotic stress in cereal crops. In: Crop improvement in the era of climate change. IK. International Publishing House Pvt Ltd, New Delhi, p 205
Saleem MY, Mukhtar Z, Cheema AA, Atta BM (2005) Induced mutation and in vitro techniques as a method to induce salt tolerance in basmati rice (Oryza sativa L.). Int J Environ Sci Technol 2:141–145
Seralini GE, Clair E, Mesnage R, Gress S, Defarge N, Malatesta M, Hennequin D, Spiroux de Vendomois J (2012) Long term toxicity of roundup herbicide and a roundup-tolerant genetically modified maize. Food Chem Toxicol 50:4221–4231
Shah F, Wu W (2019) Soil and crop management strategies to ensure higher crop productivity within sustainable environments. Sustainability 11:1485
Shevtsov VM, Serkin NV, Chanda D, Chumak VM (2003) Mutations and barley breeding for thermoperiod reaction and cold tolerance. Barley Genet Newsl 33
Stadler LJ (1928) Mutation in barley induced by X-rays and radium. Science 69:186–187
Takeda S, Matsuoka M (2008) Genetic approaches to crop improvement: responding to environmental and population changes. Nat Rev Genet 9:444–457
Tantray AY, Raina A, Khursheed S, Amin R, Khan S (2017) Chemical mutagen affects pollination and locule formation in capsules of black cumin (Nigella sativa L.). Int J Agric Sci 8(1):108–117
Tomás D, Rodrigues JC, Viegas W, Silva M (2020) Assessment of high temperature effects on grain yield and composition in bread wheat commercial varieties. Agronomy 10:499
Wade LJ, McLaren CG, Quintana L, Harnpichitvitaya D, Rajatasereekul S, Sarawgi AK, Kumar A, Ahmed HU, Sarwoto, Singh AK, Rodriguez R, Siopongco J, Sarkarung S (1999) Genotype by environment interactions across diverse rainfed lowland rice environment. Field Crop Res 64:35–50
Wang JP, Raman H, Zhou MX, Ryan PR, Delhaize E, Hebb DM, Coombes N, Mendham N (2007) High-resolution mapping of the alp locus and identification of a candidate gene HvMATE controlling aluminium tolerance in barley (Hordeum vulgare L.). Theor Appl Genet 115:265–276
Wani MR, Dar AR, Tak A, Amin I, Shah NH, Rehman R, Baba MY, Raina A, Laskar R, Kozgar MI, Khan S (2017) Chemo-induced pod and seed mutants in mungbean (Vigna radiata L. Wilczek). SAARC J Agric For 15(2):57–67
Wani MR, Laskar RA, Raina A, Khan S, Khan TU (2021a) Application of chemical mutagenesis for improvement of productivity traits in lentil (Lens culinaris Medik). Ann Biol 37(1):69–75
Wani MR, Tomlekova N, Raina A, Laskar RA, Khursheed S, Khan S, Tak MA, Bhat TA (2021b) Mutation breeding technique for the improvement of pulse crops with special reference to faba bean (Vicia faba L.). In: Bhat TA (ed) Mutagenesis, cytotoxicity and crop improvement: revolutionizing food science. Cambridge Scholars, Newcastle upon Tyne, pp 222–243
Yildiz M, Poyraz I, Çavdar A, Özgen Y, Beyaz R (2020) Plant responses to salt stress. In: Plant breeding-current and future views. IntechOpen, London
Yunus M, Paroda RS (1982) Impact of biparental mating on correlation coefficients in bread wheat. Theor Appl Genet 62:337–343
Author information
Authors and Affiliations
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2022 The Author(s), under exclusive license to Springer Nature Singapore Pte Ltd.
About this chapter

Cite this chapter
Raina, A., Laskar, R.A., Wani, M.R., Khan, S. (2022). Plant Breeding Strategies for Abiotic Stress Tolerance in Cereals. In: Roychoudhury, A., Aftab, T., Acharya, K. (eds) Omics Approach to Manage Abiotic Stress in Cereals. Springer, Singapore. https://doi.org/10.1007/978-981-19-0140-9_8
Download citation
DOI: https://doi.org/10.1007/978-981-19-0140-9_8
Published:
Publisher Name: Springer, Singapore
Print ISBN: 978-981-19-0139-3
Online ISBN: 978-981-19-0140-9
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)